










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Aedes (Stegomyia) aegypti (Linnaeus, 1762) y Aedes (Stegomyia) albopictus (Skuse, 1895) constituyen dos de las especies más importantes de mosquitos, debido a que son vectores primarios de enfermedades emergentes y reemergentes.1 Estos culícidos se consideran especies invasoras, que colonizan exitosamente varios sitios fuera de sus ámbitos nativos,2 lo que contribuye a la expansión global, emergencia y reemergencia de las arbovirosis, entre las que se incluyen el dengue, fiebre amarilla, chikungunya y Zika.3,4 Este es el caso de Ae. aegypti y Ae. albopictus, los que colonizaron con éxito en ambientes urbanos y silvestres.5
Ambas especies pueden relacionarse y competir por espacio y recursos como puede ser alimento, luz, agua, territorio, apareamiento, etc.6
En el hábitat acuático donde se crían los mosquitos es común este tipo de competencia,7 la cual puede estar ligada en diversos casos a factores densodependientes que pueden alterar algunas características del ciclo de vida de estos.8 Entre esas características se encuentran la duración del ciclo larvario, la supervivencia larvaria, y la supervivencia y talla de los adultos.9
La importancia de analizar la competencia intraespecífica en estas especies vectores radica en que la dinámica del ciclo de vida de estas puede alterarse como consecuencia de la reducción competitiva.10
Adicionalmente la competencia larvaria puede ser capaz de generar variaciones en la respuesta inmunitaria de los mosquitos,11 que podría alterar el comportamiento en la trasmisión viral.
En este trabajo nos proponemos evaluar el efecto de la competencia larvaria intraespecífica causada por baja y alta densidad de Ae. aegypti y Ae. albopictus sobre aspectos del ciclo de vida de estas especies en condiciones de laboratorio.
Métodos
Material biológico
Se utilizaron dos poblaciones de mosquitos (Ae. aegypti y Ae. Albopictus), ambas colectadas en estado de larva en el reparto Juan de Dios Fraga, La Lisa, Cuba, 2019 (23°02'43.0"N 82°29'02.0"W).
Procedimiento experimental
El estudio se llevó a cabo bajo condiciones de laboratorio a 26 ± 2 °C, con 70 ± 5 % de humedad relativa y un fotoperiodo de 12/12 (luz/oscuridad). Las larvas de Ae. aegypti y Ae. albopictus colectadas se criaron hasta la etapa adulta y se mantuvieron en jaulas de 60 x 60 x 60 cm, proporcionándoles una solución de sacarosa al 10 %.12 Para la hematofagia de las hembras se utilizaron ratones de laboratorio BALB/c dos veces por semana, suministrados por el Centro Nacional para la Producción de Animales de Laboratorio (CENPALAB), Cuba. Los ratones se mantuvieron en condiciones convencionales y se trabajó con ellos bajo las normas éticas para el uso de animales de experimentación.13
Manejo de los huevos de mosquitos
En cada jaula se colocó un recipiente con 250 mL con agua declorinada, donde se introdujo una tira de papel de filtro en su borde interior para la recolección de los huevos. Las tiras de papel con los huevos se colocaron a madurar en bandejas plásticas sobre una superficie húmeda durante un periodo de 72 h para garantizar el correcto desarrollo del embrión. Las tiras de papel de guardaron en bolsas de nylon a temperatura ambiente por un periodo nunca superior a un mes. Para la eclosión, se sumergieron las tiras con los huevos en agua a 36 °C.
Conformación de los grupos
Se contaron las larvas de primer estadio en vasos plásticos con 100 mL de agua y se transfirieron a bandejas de 29 x 20 x 4 cm con 600 mL de agua declorinada. El volumen de agua de cada réplica fue mantenido de forma constante diariamente.
Mantenimiento de estadios acuáticos y adultos
Las larvas se alimentaron con harina de pescado (0,1 g diariamente). Las pupas fueron transferidas individualmente a viales plásticos con 20 mL de agua, hasta la emergencia de los adultos, los cuales fueron examinados de forma individual para comprobar especie y sexo. Se liberaron hembras adultas en jaulas metálicas de cría de 60 x 60 x 60 cm, con el fondo cubierto con un papel blanco. Se suministró solución azucarada al 10 % de manera constante. Se colectaron y contaron los individuos muertos cada 24 h hasta la muerte del último ejemplar.
Tratamientos larvarios
Se establecieron condiciones de alta densidad (140 larvas a razón de 1 por cada 8 mL de agua) y baja densidad (70 larvas a razón de 1 por cada 8 mL) con 5 réplicas por cada especie. Las densidades utilizadas fueron seleccionadas basadas en reportes de estudios similares.11
Variables estimadas
Duración del ciclo larvario (días): Se realizó un registro del número de larvas presentes por bandeja hasta que todas las larvas transitaron al estadio de pupa.
Supervivencia de las larvas: Se determinó mediante el promedio de larvas que alcanzaron el estadio de pupa.
Supervivencia de hembras adultas: Se determinó mediante el registro de las hembras adultas recuperadas de cada densidad poblacional y su muerte en cada una de las jaulas.
Talla de hembras adultas: Se estableció mediante la medida del ala como un estimador de tamaño. Se tomaron fotografías de las alas con una cámara Leica ICC50 acoplada a un microscopio de disección, con un ocular micrómetro LEICA con un lente 4X a través del programa tpsDig2. Se seleccionaron aleatoriamente 30 hembras muertas (por cada condición de densidad poblacional y especie). Se les removió el ala izquierda para su montaje, desde la unión en la porción basal articulada del tórax, y se colocaron en un portaobjeto limpio. Para cada fotografía se seleccionaron y digitalizaron un conjunto de cuatro puntos de referencia, ubicados en puntos de inicio y finalización de manchas o venas (Fig. 1). La talla del ala se comparó midiendo el largo, el ancho del ala y el largo de la vena A.
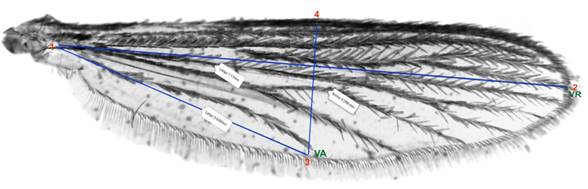
Fig. 1 -Puntos de referencia seleccionados en las alas de hembras adultas de Aedes aegypti y Aedes albopictus empleados en los ensayos de competencia larvaria. Las líneas trazadas entre puntos de referencia representan las distancias medidas. 1-2 Largo del ala: inicio de la vena A (VA) hasta finalización de la vena R (VR). 1-3 Ancho del ala: distancia desde el lóbulo anal hasta la finalización de la vena A. 3-4 Largo de la vena A: distancia desde la finalización de la vena hasta el extremo distal.14
Análisis estadístico
Los datos obtenidos de los parámetros reproductivos fueron analizados con el sistema estadístico SPSS versión 21.0. Se exploró la distribución de los datos mediante el cálculo de la simetría y la curtosis. La duración del ciclo larvario se analizó mediante la prueba t de student para comparar las medias entre las condiciones de alta y baja densidad larvaria. Para la supervivencia larvaria, se realizaron tablas de contingencia. Para determinar variaciones en la supervivencia de las hembras de Ae. aegypti y Ae. albopictus, se realizaron curvas de supervivencia mediante el estimador no paramétrico de Kaplan-Meier y, después se utilizó la prueba de Mantel-Cox para evaluar el efecto de la densidad en la supervivencia dentro de cada colonia y comparar si hubo diferencias significativas entre las colonias dentro de una misma condición. Finalmente, para el análisis de los promedios de la longitud de las alas de cada una de las colonias de campo, en comparación con la colonia control dentro de una misma condición (alta densidad o baja densidad larvaria), se determinó la normalidad, entonces se utilizó la prueba t de student para comparar las medias entre las condiciones de alta y baja densidad larvaria.
Procedimientos éticos
Todos los ensayos y procedimientos del presente estudio se realizaron tomando en consideración las normas éticas y científicas para realizar estudios biomédicos a partir de la Declaración de Helsinki y las normas éticas establecidas por la Comunidad Europea relativa a la protección de los animales utilizados para fines científicos (Directiva 22 de septiembre de 2010).13
Resultados
Duración del ciclo larvario de Aedes aegypti y Aedes albopictus
En la tabla se puede observar que el promedio de duración del ciclo larvario de Ae. aegypti en condiciones de baja y alta densidad fue de 7 y 11 días, respectivamente. Por su parte, el promedio de duración del ciclo larvario de Ae. albopictus en condiciones de baja y alta densidad larvaria fue de 6 y 10 días, respectivamente. De manera general podemos decir que no existieron diferencias significativas entre las especies en ninguna de las densidades evaluadas; sin embargo se observó diferencias significativas entre la duración promedio del ciclo larvario en condiciones de baja y alta densidad para ambas especies (U de Mann-Whitney) (p < 0,05), debido a que las larvas de ambas especies tardaron significativamente más tiempo en transitar a pupa cuando experimentaron condiciones de alta densidad larvaria que las que se criaron en condiciones de baja densidad larvaria.
Tabla - Duración del ciclo larvario (días) de Aedes aegypti y Aedes albopictus en competencia intraespecífica a baja y alta densidad larvaria en condiciones de laboratorio
| Baja | 6,89* | 1,356 | 4 | 11 | 6,21* | 2 | 5 | 12 |
| Alta | 11,15 | 3,046 | 6 | 17 | 9,75 | 2,511 | 5 | 18 |
*Diferencias significativas en baja y alta densidad en la misma especie.
DE: Desviación estándar.
Prueba de U Mann-Whitney
Supervivencia larvaria de Aedes aegypti y Aedes albopictus
En la figura 2 se aprecia como las dos especies mostraron porcentajes similares de supervivencia larvaria en cada una de las densidades larvarias. No obstante, se evidenció una disminución en el porciento de supervivencia en las larvas que se criaron en condiciones de alta densidad, en comparación con las que se criaron en baja densidad larvaria en ambas especies, mostrando diferencias significativas (p < 0,05).

Supervivencia de hembras adultas de Aedes aegypti y Aedes albopictus
Los resultados de la supervivencia de hembras adultas en competencia intraespecífica muestran que en condiciones de baja densidad larvaria no existen diferencias significativas entre Ae. aegypti y Ae. albopictus (U de Mann-Whitney) (p = 0,16). Sin embargo, en condiciones de alta densidad larvaria Ae. aegypti presentó una menor supervivencia que Ae. albopictus (log Rank, p < 0,05) (Fig. 3).
En cuanto a la supervivencia de las hembras de Ae. aegypti en condiciones de baja y alta densidad fue de 35 y 23 días, respectivamente. Mientras que en Ae. albopictus fue de 31 y 27 días, respectivamente. De manera general, al comparar cada una de las especies en las diferentes densidades larvarias, se encontró que la supervivencia de las hembras adultas disminuyó de manera significativa (log Rank, p < 0,05), en condiciones de alta densidad larvaria (Fig. 3).

Fig. 3 -Supervivencia de hembras de Aedes aegypti (A) y Aedes albopictus (B) en los tratamientos de competencia intraespecífica de baja y alta densidad en condiciones de laboratorio.
Talla corporal de hembras adultas de Aedes aegypti y Aedes Albopictus
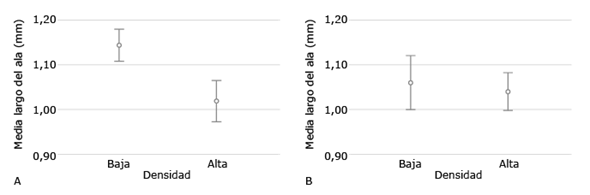
Al comparar el largo del ala de Ae. aegypti entre las dos densidades larvarias no se encontraron diferencias significativas (p = 0,16), siendo de 1,14 ± 0,01 mm de longitud) el tamaño de las alas en los mosquitos que fueron criados en condiciones de baja densidad larvaria y de 1,02 ± 0,01 mm de longitud (Fig. 4 A) los criados en condiciones de alta densidad larvaria. No se encontró diferencias significativas (p = 0.199) en el promedio de la longitud de las alas de la población de Ae. albopictus que se mantuvieron en baja y alta densidad larvaria, siendo de 1,05 ± 0,01 mm y 1,04 ± 0,01 mm, respectivamente (Fig. 4 B).
Discusión
Duración del ciclo larvario
Al comparar la duración del ciclo larvario entre las dos especies estudiadas a bajas densidades larvarias, se encontró que ambas poblaciones presentaron un tiempo de desarrollo muy similar entre sí (6 y 7 días). Este tiempo es informado por Vargas (1998), quien plantea que el estado larvario con frecuencia comprende de 4 a 10 días, en condiciones favorables de temperatura entre 25-29 °C.15
La duración del ciclo larvario fue altamente sensible a la condición de alta densidad debido a que la población de Ae. aegypti como la de Ae. albopictus presentaron un retraso en el proceso de pupación, que se extendió en algunos casos hasta los días 17 y 18 del experimento. Estos resultados concuerdan con lo reportado en estudios anteriores, donde se observó una prolongación del tiempo de desarrollo larvario en condiciones de alta densidad.16
Este retardo de las larvas en condiciones de alta densidad pudiera deberse a la competencia por alimento.17 La relación de este retraso con las reservas nutricionales tiene un efecto negativo sobre los últimos estadios larvales, los cuales son claves para que las larvas obtengan una masa corporal crítica que le permita transitar al estado de pupa. Cuando este recurso se ve disminuido o no se encuentra disponible, las larvas llegan a esta etapa de la metamorfosis con un tamaño más reducido.18 Otros factores dependientes de la densidad se consideran responsables de tener un impacto negativo en el desarrollo larvario en condiciones de alta densidad larvaria, como la disponibilidad del alimento, el incremento de materiales de desecho, producto de la descomposición de larvas muertas, exuvias y productos de la excreción de las larvas como nitrógenos en forma de amoniaco, los cuales pueden ser responsables del retraso en el crecimiento larvario.19
Supervivencia larvaria
Las dos especies mostraron porcentajes similares de supervivencia larvaria en competencia intraespecífica en cada una de las densidades larvarias, aunque en las larvas que se criaron en condiciones de alta densidad se observó una disminución en el tiempo de supervivencia.
Este resultado podría estar asociado al fenómeno del canibalismo. Aunque este no es de esperarse en larvas que son filtradoras de detritus como ocurre en especies de Aedes, se ha reportado con anterioridad tanto en Aedes aegypti20 como en Aedes albopictus,21) en estudios asociados a ambientes larvarios con baja disponibilidad de alimento, lo que conduce a una considerable pérdida de individuos. Algunos estudios sobre larvas de Aedes triseriatus (Say, 1823) mostraron canibalismo entre individuos de esta especie y su vulnerabilidad, lo que se constató en los cuatro estadios larvales.22
Supervivencia y talla corporal de hembras adultas
La supervivencia de hembras adultas en competencia intraespecífica mostró que a baja densidad larvaria no existió diferencias significativas entre ambas especies. Sin embargo, en alta densidad larvaria, se evidenció que Ae. aegypti presentó una menor supervivencia que Ae. albopictus. Esto sugiere que la competencia larvaria en condiciones de alta densidad tiene efectos significativos sobre la longevidad de hembras adultas en Ae. aegypti. Otros estudios también encontraron efectos negativos en la supervivencia de Ae. Aegypti, pero no en Ae. albopictus,7 por lo que el autor sugiere que puede ser un fenómeno con insuficiente evidencia, específico de cada especie y condición. Inferimos que Ae. aegypti tiene la posibilidad de dosificar sus puestas en diferentes criaderos y por lo general estos son de baja densidad larvaria. En consecuencia, si la hembra resultante se sometió a estrés competitivo en condiciones de alta densidad larvaria, pudiera verse afectada su longevidad.
Estos resultados concuerdan con algunos estudios de campo23 y de laboratorio.7 Lo anterior pudiera deberse a una posible competencia por el alimento, lo que coincide con lo informado por Brack y otros.23
Diversos autores,24 en colonias de mosquitos que experimentaron altas densidades larvarias, demostraron la existencia de una disminución en la talla de los adultos, lo que evidencia una relación entre la supervivencia y el tamaño corporal del adulto.
En nuestro estudio no se hallaron diferencias significativas al comparar el largo del ala de ambas especies en condiciones de bajas y altas densidades larvarias. Estos resultados no coinciden con estudios realizados por otros autores que plantean afectación de la talla del adulto, independientemente de la especie.25 Podemos destacar que en el presente estudio se apreció una tendencia a la disminución de la longitud del ala de Ae. aegypti en altas densidades, resultado que pudiera estar asociado al tamaño de muestra o a la prueba estadística utilizada.
Nuestros resultados no coinciden con algunos estudios donde se evidencia que la alta densidad larvaria tiene un efecto negativo en el tamaño corporal del mosquito en su fase adulta.26
La talla de los mosquitos tiene una base genética. No obstante, las modificaciones en su hábitat larvario determinadas por la alta densidad poblacional pueden generar variaciones en su tamaño.27 Los adultos con menor talla pueden estar influenciados por la disponibilidad de alimento en la etapa inmadura debido al estrés competitivo que experimentan en su estado larvario.24 Mientras que los mosquitos criados a bajas densidades poseen mayores reservas calóricas, su tamaño es mayor en la fase adulta que los criados a altas densidades y con reservas deficientes.28
Existen posibles repercusiones sobre la capacidad vectorial de Ae. aegypti frente al virus del dengue (VDEN). Una marcada disminución en el tamaño corporal de las hembras adultas (generado por el efecto de la competición bajo condiciones de alta densidad larvaria) podría estar modulando componentes tan importantes dentro de la capacidad vectorial como la tasa de picadura y dispersión.29
Varios estudios demostraron que las hembras más pequeñas tienden a alimentarse con más frecuencia de sangre, unido a una mejor dispersión que hembras con mayor talla, lo que promueve una mayor oportunidad de contacto del mosquito con un hospedero, hecho que podría potenciar la transmisión del virus.28 Algunos estudios también mostraron resultados similares en hembras de Ae. albopictus que experimentaron en su etapa larvaria efectos competitivos, disminuyeron su tamaño y resultaron más susceptibles de ser infectadas y de diseminar el VDEN que hembras de mayor tamaño.29
La tasa de supervivencia de las hembras adultas disminuyó bajo condiciones de altas densidades, lo que puede asociarse con el tamaño de los mosquitos. Esta asociación permite entender la dinámica de un sistema compensatorio entre el tamaño y la supervivencia. A pesar de que los mosquitos sobreviven menos tiempo, determinado por su pequeño tamaño, pudieran tener la capacidad para alimentarse más a menudo de un hospedero y la habilidad de recorrer mayores distancias, lo cual puede estar determinado por su pequeño tamaño.29
Conclusiones
El efecto de la relación larvaria intraespecífica bajo condiciones de alta densidad presentada en los hábitats de Ae. aegypti y Ae. albopictus tuvo un efecto negativo en cada una de las variables del ciclo de vida evaluadas. Estas alteraciones observadas son importantes dentro de los componentes de la capacidad vectorial con implicaciones en la epidemiología de las enfermedades transmitidas por estos mosquitos.
Los resultados obtenidos a nivel de laboratorio nos pueden ayudar a dar respuestas al comportamiento que pudieran presentar estos mosquitos en su entorno natural. Este panorama, en última instancia, podría estar generando un aumento en los patrones de transmisión del VDEN, así como un mayor riesgo de infección para la población humana.

