










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Investigaciones Biomédicas
versión On-line ISSN 1561-3011
Rev Cubana Invest Bioméd v.17 n.3 Ciudad de la Habana sep.-dic. 1998
Instituto de Ciencias Básicas y Preclínicas "Victoria de Girón"
Efecto de la privación de agua sobre el bostezo espontáneo en la rata
Dr. José M. Anías Calderón, Dr. Ariel Santana Felipe y Dra. Araceli Chia MenaRESUMEN
Con el objetivo de comprobar si la hormona arginina-vasopresina (AVP) tiene alguna influencia sobre el bostezo espontáneo en ratas adultas de la cepa Wistar se efectuaron 2 series experimentales: a) estudio del efecto de la privación de agua (WD) sobre el bostezo y b) estudio del efecto de la WD sobre el bostezo en animales en ayuno durante 96 h (el ayuno suprime el bostezo espontáneo a las 24 h en los animales estudiados). Se observó que la WD no afecta de manera significativa el bostezo en los animales con alimentación normal. En los animales en ayuno se observó la supresión del bostezo en el grupo control (ayuno y agua ad libitum) mientras que en el grupo experimental (WD y ayuno) se evitó la supresión que sobre el bostezo provoca el ayuno. Se infiere de estos resultados que la AVP participa en la regulación del bostezo en las condiciones estudiadas; la forma en que lo hace queda fuera del alcance de este estudio.Descriptores DeCS: BOSTEZO/fisiología; ARGIPRESINA; PRIVACION DE AGUA; RATAS WISTAR.
En diversas investigaciones efectuadas sobre el bostezo se han estudiado en relación con esta conducta el efecto de diferentes formas de estrés,1-5 la acción de la privación alimentaria6 y la repercusión de la hipofisectomía.7 Todos estos estudios están relacionados con los mecanismos de liberación de la ACTH, hormona que resulta potente inductora del bostezo.8,9
Es conocido que en la regulación de la liberación de la ACTH participan la hormona liberadora de corticotropina (CRH), su principal estimulador, la hormona oxitocina (OT) y la hormona arginina-vasopresina (AVP).10-12 La OT tiene un potente efecto inductor del bostezo,13-15 y el factor CRH aunque no ha sido utilizado para inducir el bostezo, necesariamente ha sido involucrado en todas las manipulaciones que influyen sobre esta conducta, como referimos en el párrafo anterior. Sin embargo, el efecto de la AVP sobre el bostezo no ha sido confirmado hasta el momento, aunque esta hormona es notablemente modificada por la privación alimentaria16,17 y el estrés.18,19
La AVP tiene una amplia distribución en el cerebro, por lo que se han adjudicado funciones de neurotransmisor.10 No obstante su sitio de síntesis principal es el núcleo paraventricular (PVN) donde también se sintetizan OT y CRH.10,20 Ya que el PVN ha sido reconocido como estructura importante en la generación del bostezo15,21 es de esperar que la AVP tenga alguna influencia en la conducta.
Otro aspecto de coincidencia es que la AVP es fuertemente influenciada por las hormonas sexuales,22,23 de manera similar a como ocurre con el bostezo.24,25 Tomando en cuenta todo lo expuesto anteriormente y con el objetivo de comprobar si la AVP tiene alguna influencia sobre la frecuencia de aparición del bostezo, en este trabajo estudiaremos el efecto que tiene la privación de agua, que estimula la secreción de AVP,26,27 sobre el bostezo.
MÉTODOS
Se efectuaron 2 series experimentales diferentes: a) estudio del efecto de la privación de agua (WD) sobre la frecuencia de aparición del bostezo espontáneo en ratas con la alimentación habitual, y b) estudio del efecto de la WD en ratas sometidas al ayuno durante 96 h, procedimiento que ha demostrado que suprime el bostezo espontáneo (trabajo en publicación).En dichas series experimentales se utilizaron ratas adultas (250-300 g) de la cepa Wistar, cada animal una vez utilizado fue desechado. Para mejor observación del bostezo espontáneo registramos la frecuencia de aparición del mismo en 80 animales en 3 ocasiones, con el objetivo de seleccionar a los que con más frecuencia y estabilidad bostezaran. Todos los animales usados fueron cuidados y utilizados según metodología establecida por el comité sobre cuidados y uso de animales de laboratorio del consejo científico nacional (USA). En todos los casos la frecuencia de aparición del bostezo fue monitoreada por 2 observadores entrenados.
Las observaciones se efectuaron en el horario de 15:30 a 16:30 h. Los animales para la observación fueron colocados en recipientes de cristal transparentes, circulares de 18 cm de diámetro y 9 cm de alto, cubierto con una lámina de plástico que permitía la entrada de aire sin dificultad. Dichos recipientes fueron colocados sobre una mesa en un local aislado de ruidos a una temperatura entre 20-22 oC En cada observación se controlaron 6 animales (3 experimentales y 3 controles).
- Estudio del efecto de la WD sobre la frecuencia de aparición de bostezo espontáneo en ratas con alimentación normal: a un grupo de 8 animales se les sometió a la WD con alimento ad libitum durante 96 h. Otro grupo de 8 animales sirvió como grupo control y no fue sometido a ningún tipo de privación. En ambos grupos de animales se observó la frecuencia de aparición del bostezo en los siguientes momentos: 24 h antes de iniciar la serie experimental, a las 48,72 y 96 h de haber iniciado la WD en el grupo experimental.
- Estudio del efecto de la WD sobre la frecuencia de aparición del bostezo en ratas sometidas al ayuno: un grupo de 8 animales fue sometido a la privación total de líquidos y de alimentos durante 96 h. Otro grupo de 8 animales sólo fue sometido a ayuno pero con libre acceso al agua. En ambos grupos se observó la aparición del bostezo espontáneo en los momentos indicados en sección (a).
RESULTADOS
La WD no afectó la frecuencia de aparición del bostezo de manera significativa al comparar estos resultados con el grupo control. Tampoco hubo diferencias significativas al comparar la frecuencia de aparición del bostezo antes de iniciar el experimento, con respecto al resto de los registros realizados en los diferentes momentos de observación (fig. 1).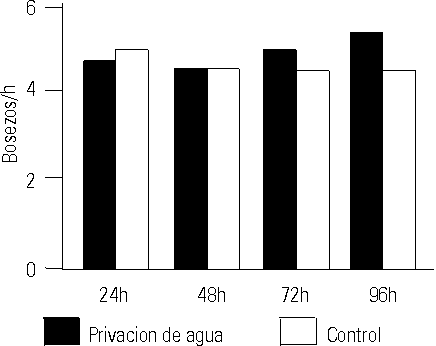
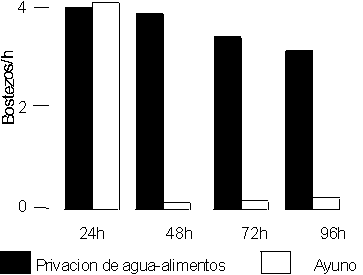
En los animales sometidos al ayuno se observó la supresión del bostezo a las 24 h de estar bajo este régimen, situación que se mantuvo durante todas las horas de observación del experimento. En los animales sometidos a la privación de líquido y de alimento, el bostezo se mantuvo a frecuencias similares que las que se observaron en ellos antes del experimento. Al comparar los privados de alimento y líquido con los animales privados sólo de alimento existieron diferencias altamente significativas entre ambos grupos en todos los momentos del experimento, p < 0,01 (fig. 2).
DISCUSIÓN
Los resultados obtenidos demuestran que la WD no deprime el bostezo, sino por el contrario lo preserva incluso ante condiciones como el ayuno que provoca la supresión del bostezo.6 Es conocido que durante la WD los niveles de la AVP se incrementan tanto en plasma como en diversas regiones del cerebro,26,27 entre ellas el PVN y SO.28 Este incremento de AVP relacionado con la preservación del bostezo sugiere que la AVP debe participar de alguna forma en la generación de dicha conducta, ya que el régimen de ayuno que suprime el bostezo provoca decrecimientos significativos de AVP en PVN y SO.16,17En otro trabajo donde se compararon ratas privadas de alimento (FD) con ratas WD y con ratas privadas simultáneamente de agua y de alimento (FWD), procedimientos que duraron 60 h, se observó que en las 3 condiciones se incrementaron los niveles de corticosterona plasmática, mientras que la ACTH y las catecolaminas no cambiaron. Sin embargo, la concentración plasmática de AVP fue normal en los FD mientras que en WD y en WFD se incrementó significativamente. Además, la AVP mRNA se incrementó en PVN por WD, y FD produjo ligero decrecimiento y previno el efecto de WD. El CRF mRNA en el PVN fue reducido en un 27 % por FD mientras que WD lo disminuyó en un 67 %, pero la FWD produjo decrementos similares a los ocurridos por FD. En la hipófisis anterior los niveles de POMC mRNA no cambiaron por la WD, pero se incrementaron significativamente por FD y FWD.29
Como puede apreciarse el procedimiento descrito en el trabajo referido fue similar al usado por nosotros, por lo que es de esperar que los cambios hormonales referidos hayan ocurrido similarmente en nuestros experimentos, lo que nos sugiere que los cambios observados en el bostezo se deben a la acción de la AVP.
En estudios efectuados en ratas WD durante 48 h, se demostró que se produce una intensa activación en el sistema magnocelular AVP del PVN, y una disminución de las células corticotrofas de la hipófisis, por lo que se infirió que en estas circunstancias el sistema AVP no desempeña una importante función en la regulación de la secreción de ACTH.27 Lo que nos hace pensar que en nuestros resultados no se debieron a un incremento en la acción directa de la ACTH.
En otro trabajo utilizando WD durante 60 h, se comprobó esto y se planteó que se debía a una disminución en la actividad del sistema parvicelular del PVN y a una caída de la actividad de los receptores para AVP en la hipófisis. Concluyeron en concordancia con otros autores que la relación entre la secreción de AVP y CRS en el PVN tiene una importante función en la modulación de los receptores AVP de la hipófisis y en la caída de la sensibilidad los glucocorticoides que de manera sostenida se secretan en el estrés crónico.28 Esto debió influir en nuestros resultados.
Es interesante que durante el estrés crónico la administración de antagonistas para los receptores AVP1 es seguida de un incremento en las conductas estereotipadas,30 lo que indica que la AVP puede modificar conductas motoras, además de las influencias que tiene sobre conductas integrativas como la atención y el aprendizaje.10
En relación con el eje hipotálamo adrenal la adrenalectomía no alteró la expresión de los receptores AVP en septum, hipocampus y núcleo arcuato9 e incrementó AVP mRNA en la región parvicelular del PVN siendo este efecto dependiente de la activación de los receptores AVP tipo 2 (glucocorticoideos), los autores no encontraron cambios en la región magnocelular, ni con la adrenalectomía, ni con la administración de glucocorticoides.25 La administración de dosis suprafisiológicas de corticosterona deprimió la expresión de los receptores AVP en septum e hipocampus.9
De todo lo planteado podemos especular que la AVP podría influir sobre el bostezo modificando el balance magnocelular (AVP)-paravicelular (CRF/AVP) en el PVN o quizás actuando en otras regiones del cerebro.
Otra posible explicación de mi resultado estaría en la oxitocina, hormona que induce el bostezo,13,15 y se ha reportado que se incrementa su liberación por la WD,30,31 por lo que pudiera esto ser la causa de la preservación del bostezo ante el ayuno. Por otro lado se ha sugerido que los donadores de óxido nítrico inyectados en ventrículos laterales o en el PVN inducen bostezo y erección peneana por activación de la transmisión oxcitocinérgica en el núcleo paraventricular, y se ha dicho que el óxido nítrico puede ser un neuromodulador en la liberación de AVP.
En el análisis de los neurotransmisores involucrados en la regulación de la AVP, se ha reportado que microinyecciones de adrenalina y dopamina en el núcleo PVN incrementan significativamente AVP en plasma.32 Se plantea que los receptores dopaminérgicos del PVN facilitarían la secreción de AVP.33 Este pudiera ser un mecanismo que explicaría la inducción del bostezo por microinyección de apomorfina en PVN.21
En conclusión, los resultados obtenidos por la WD sugieren que la AVP desempeña una función importante en la regulación del bostezo en las condiciones de nuestro estudio. No podemos concluir si su efecto es directo o es mediado por otras de las hormonas afines entre las cuales la oxitocina debe tener especial consideración, o si su acción es por vía del eje hipofisario, o en otras regiones del SNC. Son necesarios otros estudios para mejor comprensión de nuestros resultados.
SUMMARY
To confirm if arginine-vasopressin (AVP) hormone has any influence on spontaneous yawing in Wistar adult rats, two experimental series were performed: a) study of effect of water deprivation (WD) on yawing, and b) study of WD effect on yawing in fasting animals over 96 h. (fasting suppress spontaneous yawing within 24 h. in study animals). We observed that WD not damage significantly the yawing in animal given a normal feeding. In fasting animals, suppression of yawing was observed in controls (fasting and water ad libitum) while in experimental group (WD and fasting) suppression of yawing from fasting, was avoided. From these results, we infer AVP is involved in yawing regulation under study conditions, and mechanism of action is beyond reach of this study.Subject headings: YAWING/ physiology; ARGIPRESSIN; WATER DEPRIVATION; RATS, WISTAR.
REFERENCIAS BIBLIOGRÁFICAS
- Bourson A, Moser PC. The effect pre and post operative procedure on physostigmine and apomorphine-induced yawning rats. Pharmacol Biochem Behav 1989;34:915-7.
- Neumann BG, Troncone LRP, Braz S, Tufik S. Modification on dopaminergic and cholinergic system induced by the water tank technique: analysis through yawning behavior. Arch Intern Parmacodin Ther 1990;308:32-8.
- Nunes JGP, Tufik S. Validation of the modified multiple plataform method of paradoxical sleep deprivation in rats. Sleep Res 1994;23:419-21.
- Tufik S, Troncone LRP, Braz S, Silva-Filho AR, Neumann BG. Does REM sleep deprivation induce subsensitivity of presinaptic acetylcholine receptors in the rat brain. Eur J Pharmacol 1987;140:215-9.
- Tufik S, Nathan CL, Neumann B, Hipolide DC, Lobo LL, Medeiros R. et al. Effects of stress on drug induced yawning: constant vs. intermittent stress. Physiol Behav 1995;58:181-4.
- Nasello AG, Tieppo CA, Felicio LF. Apomorphine-induced yawning in the rat: Influence of fasting and time of day. Physiol Behav 1995;57:967-71.
- Serra G, Collu M, Loddo S, Celasio G, Gessa GL. Hypophysectomy prevents yawning and penile erection but not hipomotility induced by apomorphine. Pharmacol Biochem Behav 1983;19:917-9.
- Ferrari W, Gessa GL, Vargiu L. Behavioural effects induced by intracisternally injected ACTH and MSH. Ann NY Acad Sci 1963; 104:330-43.
- Gessa GL, Pisano N, Vargiu L, Grabal F, Ferrari W. Stretching and yawning movements after intracerebral injection of ACTH. Rev Can Biologie 1967;26:229-36.
- Le Moal M, Mormede P, Stinus L. The behavioral neuroendocrinology of arginine vasopressin, adrenocorticotropic hormone and opioids. En: Nemeroff CB. CRC Press, 1992;365-71.
- Kjaer A. Vasopressin as a neuroendocrine regulator of anterior pituitary hormone secretion. Acta Endocrinol 1993;129:489-96.
- Kjaer A, Kniggi U, Bach FW, Warberg J. Histamine and stress-induced secretion of ACTH and beta-endorphin: involvement of corticotropin-releasing hormone and vasopressin. Neuroendocrinol 1992;56:419-28.
- Argiolas A, Melis MR, Gessa GL. Intraventricular oxytocin induces yawning and penile erections in rats. Eur J Pharmacol 1985;117:395-6.
- Melis MR, Stancampiano R, Argiolas A. Hippocampal oxytocin mediates apomorphine induced penile erection and yawning. Pharmacol Biochem Behav 1992;42:61-6.
- Melis MR, Argiolas A, Gessa GL. Oxytocin-induced penile erection and yawning: site of action in the brain. Brain Res 1986;415: 98-104.
- Arlette J, Burlet M, Jhanwar-Uniyal M, Chapleur-Chateau M, Burlet CR, Leibowitz SF. Effect of food deprivation and refeeding on the concentration of vasopressin and oxitocyn in discrete hypothalamic sites. Pharmacol Biochem Behav 1992;43:897-905.
- Ogasa T, Hashimoto K, Suermaru S, Kageyama J, Ota Z. The magnocellular arginine-vasopressin mRNA responds differently to food deprivation between the supraoptic and paraventricular nuclei of the hypothalamus in adrenalectomized rats with low corticosterone replacement. Brain Res 1992;583:45-53.
- Ivanyi T Wiegant VM, De Wied D. Differential effects of emotional and physical stress on the central and peripheral secretion of neurohypophysial hormones in male rats. Life Sci 1991;48:1309-16.
- Onaka T, Yagi K. Effects of novelty stress on vasopressin and oxytocin secretion by the pituitary in the rat. J Neuroendocrinol 1993;5:365-9.
- Makuio S, Smith MA, Gold PW. Increased expression of corticotropin-releasing hormone and vasopressin- -messenger ribonucleic acid (mRNA) in the hipothalamic paraventricular nucleus during repeated stress: association with reduction in glucocorticoid receptor mRNA levels. Endocrinology 1995;136:3299-3309.
- Melis MR, Argiola A, Gessa GL. Apomorphine induced penile erection and yawning: site of action in the brain. Brain Res 1987;415:98-104.
- De Vries GJ, Al-Shamma HA. Sex differences in hormonal responses of vasopressin pathways in the rat brain. J Neurobiol 1990;21:686-8.
- De Vries GT, Buijs RM, Sleiter AA. Gonadal hormone actions on the morphology of vasopressinergic inervation of adult rat brain. Brain Res 1984;298:141-3.
- Berendsen HG, Nickolson UJ. Androgenic influences on apomorphine induced yawning in rats. Behav Neurol Biol 1986;33:123-8.
- Holmgren B, Urba-Holmgren R. Sex hormone influence on yawning behavior. Acta Neurobiol Exp 1980;40:515-9.
- Hauger RL, Aguilera G. Regulation of pituitary corticotropin releasing hormone (CRH) receptors by CRH: interaction with vasopressin. Endocrinology 1993;133:1708-14.
- Aguilera G, Lightman SL, Kiss A. Regulation of the hypothalamic-pituitary-adrenal axis during water deprivation. Endocrinology 1993;132:241-8.
- . Regulation of pituitary ACTH secretion during chronic stress. Front Neuroendocrinol 1994;15: 321-50.
- Kiss A, Jezova D, Aguilera G. Activity of the hypothalamic pituitary adrenal axis and sympathoadrenal system during food and water deprivation in the rat. Brain Res 1994;663:84-92.
- Chen X, Herbert J. Alterations in sensitivity to intracerebral vasopressin and the effects of a V1 receptor antagonist on cellular autonomic and endocrine responses to repeated stress. Neuroscience 1995;64:687-97.
- Fenelon VS, Theodosis DT, Poulain DA. Fos synthesis in identified magnocellular neurons varies with phenotype, stimulos, location in the hypothalamus and reproductive state. Brain Res 1994;662:165-77.
- Yamaguchi K, Hama H. Evaluation for roles of brain prostaglandins in the catecholamine-induced vasopressin secretion in conscious rats. Brain Res 1993;607:149-53.
- Yamaguchi K, Hama H, Watanabe K, Adachi C. Effect of dopamine injection into the anteroventral third ventricular region and the paraventricular nucleus on vasopressin secretion in concious rats. Acta Endocrinol 1992;127:420-4.
Dr. José M. Anías Calderón. Instituto de Ciencias Básicas y Preclínicas "Victoria de Girón". Ave. 146 y 31 No. 3102, Playa, Ciudad de La Habana 16, CP 11600, Cuba.

