










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Investigaciones Biomédicas
versión impresa ISSN 0864-0300
Rev Cubana Invest Bioméd vol.33 no.2 Ciudad de la Habana abr.-jun. 2014
ARTÍCULO ORIGINAL
Análisis del efecto de la sal en el desarrollo de obesidad
Analysis of the effect of salt intake on the development of obesity
Joel Cerna CortésI, María Irene Díaz RevalII, Víctor Hugo Cervantes KardaschI, Alina Marín CárdenasIII, Víctor M. Hernández Escalante III, Sergio Adrián Montero CruzI
I Facultad de Medicina, Universidad de Colima, Colima, Colima, México.
II Centro universitario de investigaciones biomédicas, Universidad de Colima, Colima, Colima México.
III Facultad de Medicina de la Universidad Autónoma de Yucatán, Merida, Yucatan, Mexico.
RESUMEN
Introducción: el transporte de la glucosa y de muchos aminoácidos en el intestino es realizado por el cotransportador SGLT1 únicamente si esta unido al ion sodio. La sal aporta un ion sodio por cada molécula que se consume y en los humanos su ingesta comúnmente es de diez veces más de la cantidad necesaria y generalmente se acompaña de dietas ricas en carbohidratos. Este trabajo se planteo pensando en que una estrategia simple para reducir de peso sería el disminuir la cantidad de sal en los alimentos.
Objetivo: estudiar el efecto que tiene la sal en la dinámica de la absorción de glucosa y el efecto de una dieta rica en carbohidratos y sal en el desarrollo de obesidad en ratas Wistar.
Métodos: para corroborar la hipótesis se evaluó el efecto de la sal en la dinámica de la absorción de la glucosa en el intestino realizando curvas de tolerancia a la glucosa con sal y sin sal. También se analizó si una dieta rica en carbohidratos y sal favorece el desarrollo de obesidad en ratas Wistar.
Resultados: los experimentos mostraron que la ingesta de sal no influye en la dinámica de la absorción intestinal de la glucosa, ni en el desarrollo de obesidad en la rata Wistar.
Conclusión: el sodio que de manera natural recircula desde el citoplasma de los enterocitos hacia la luz del intestino mantiene saturado al cotransportador de la glucosa SGLT1 y garantiza en todo momento el transporte de la glucosa que se ingiere en la dieta.
Palabras clave: Transporte de glucosa en intestino, dieta elevada en sal, obesidad, transportador SGLT1.
ABSTRACT
Introduction: Intestinal transport of glucose and many amino acids is performed by the SGLT1 cotransporter only when the latter is bound to the sodium ion. Salt contributes a sodium ion per molecule ingested. Human salt intake is often tenfold the required amount, and is generally accompanied by a carbohydrate-rich diet. The present study is based on the assumption that reducing the amount of salt in foods is a simple weight-loss strategy.
Objective: Study the effect of salt on glucose absorption dynamics and the effect of a diet rich in carbohydrates and salt on the development of obesity in Wistar rats.
Methods: To corroborate the hypothesis, an evaluation was conducted of the effect of salt on intestinal glucose absorption, based on glucose tolerance curves with and without salt. An analysis was also made of whether a diet rich in carbohydrates and salt leads to the development of obesity in Wistar rats.
Results: Experiments showed that salt intake does not influence intestinal glucose absorption or the development of obesity in Wistar rats.
Conclusion: Sodium naturally recirculating from the cytoplasm of enterocytes to the intestinal lumen keeps the SGLT1 glucose cotransporter saturated and at all times ensures the transport of the glucose ingested in the diet.
Keywords: intestinal glucose transport, salt-rich diet, obesity, SGLT1 transporter.
INTRODUCCIÓN
Según la organización Mundial de la Salud (OMS) la obesidad se define como una acumulación anormal o excesiva de grasa originada por factores tales como la herencia genética; el comportamiento del sistema nervioso, endocrino y metabólico; y el tipo o estilo de vida que se lleve. En conjunto puede haber dos causas principales: mayor ingesta de calorías que las que el cuerpo gasta y/o menor actividad física que la que el cuerpo necesita. Se ha establecido que los hombres con más de un 25 % de grasa corporal y las mujeres con más de un 30 % son obesos y de acuerdo a este criterio se ha reportado que alrededor del mundo existen 300 millones de personas obesas1. El tejido graso produce proteínas bioactivas conocidas como adipocitocinas2 tales como el factor de crecimiento epidermal que se une a la heparina (HB-EGF), la leptina, el factor de necrosis tumoral alfa (TNF-a), el inhibidor del activador del plasminógeno tipo 1 (PAI-1), la resistina y la adiponectina. La expresión de todas estas adipocitocinas (a excepción de la adiponectina) se incrementan con la acumulación de grasa visceral contribuyendo al desarrollo de enfermedades crónicas como las cardiacas, la diabetes, la hipertensión arterial, los eventos cerebrovasculares, la insuficiencia renal y algunas formas de cáncer2-8.
El adenosín trifosfato (ATP) es la principal molécula energética para realizar las funciones fisiológicas del cuerpo humano y puede generarse a partir de tres fuentes nutricionales que son los carbohidratos, las grasas y las proteínas. Si los carbohidratos en una dieta son abundantes y superan los requerimientos fisiológicos y la capacidad de almacenarlos en el hígado en forma de glucógeno, entonces el excedente será precursor de la biosíntesis de lípidos favoreciendo el incremento de la cantidad de grasa corporal9. La glucosa es la principal fuente de energía para el metabolismo celular y como carbohidrato, generalmente se ingiere en forma de almidón que es un polímero, el cual es degradado por las enzimas amilasa (presente en la saliva y liberada por el páncreas) y maltasa (liberada por el páncreas en el duodeno). La molécula de glucosa ingerida en la dieta es transportada desde la luz del intestino delgado hacia el torrente sanguíneo por el co-transportador de sodio y glucosa SGLT10. El transporte de la glucosa a través de la membrana intestinal se divide en dos etapas. En primer lugar, el transporte activo de los iones sodio desde el interior de las células epiteliales hacia el torrente sanguíneo, provoca el descenso de la concentración intracelular del ion. Esta reducción induce el paso de sodio, desde la luz intestinal al interior de la célula epitelial a través del borde en cepillo, gracias a una difusión facilitada. El sodio se combina primero con la proteína de transporte SGLT1, la cual para llevar a cabo su función debe unir a la molécula de glucosa, de modo que tanto el sodio como la glucosa se transportan juntos hacia el citoplasma del enterocito. Una vez allí, unas proteínas de transporte y enzimas facilitan la difusión de la glucosa hacía el espacio paracelular a través de la membrana basolateral, y de allí a la sangre11.
La cantidad de carbohidratos que un individuo ingiere y absorbe por el intestino por día, varía con los hábitos alimenticios de la población y con su nivel económico. Si no incluimos a los esquimales, que prácticamente no comen carbohidratos, el resto del mundo tiene una dieta en la que el 50 al 80 % de las calorías son aportadas por los glúcidos12. Los alimentos ricos en proteínas, como la carne, le leche, los huevos, tienen un costo mayor que el maíz, el trigo, el arroz o las papas, que son ricos en carbohidratos. De ese modo, los sectores más pudientes de una sociedad comen, proporcionalmente, más proteínas12.
El presente trabajo de investigación se fundamentó en los siguientes hechos: 1) que el transporte de glucosa y muchos aminoácidos en el intestino es dependiente de sodio10. 2) que el consumo de sal (cuya composición química es un ion cloro y un ion sodio) por los humanos generalmente es 10 veces mayor de la cantidad necesaria13 y que la alimentación rica en carbohidratos es común en la población y puede producir obesidad14. Como objetivos de la investigación se planteó estudiar el efecto que tiene la sal en la dinámica de la absorción de glucosa y el efecto de una dieta rica en carbohidratos y sal en el desarrollo de obesidad en ratas Wistar.
MÉTODOS
Evaluación del efecto de la sal sobre las concentraciones de glucosa que se absorben desde la luz intestinal hacia el torrente sanguíneo de la rata
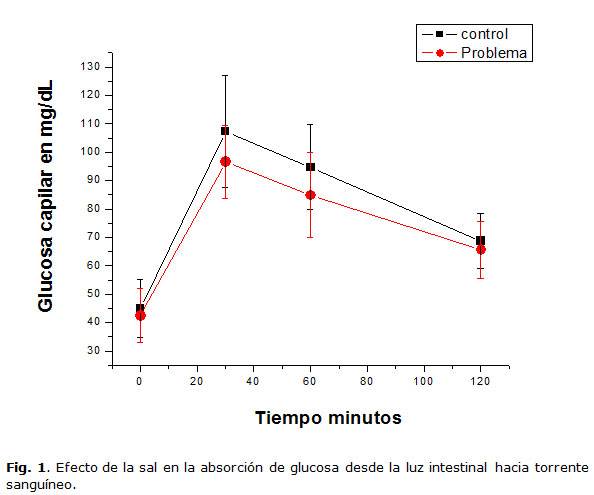
Dos grupos de 15 ratas Wistar de un peso de entre 200 y 250 gramos, se sometieron a un ayuno de 12 horas; Posteriormente se midió la glucosa capilar (lectura en el tiempo 0 minutos) para ello, se cortó el extremo de la cola de la rata y se tomó una gota de sangre que fue colocada en un glucómetro de la marca contour TS. Luego a cada una de las ratas de ambos grupos, se le administró por sonda oral 0.8 mililitros por cada 200 gramos de peso de una solución de glucosa (Técnica Química S.A. Cat. No. 61040) a una concentración de 1 g/Kg (1.55 molar), lo cual correspondió a una curva de tolerancia adaptada del humano a la rata. En el grupo control la solución de glucosa no contenía sal, mientras que en el grupo problema la solución de glucosa contenía sal (sigma aldrich Cat. No. 57653) a una concentración equimolar a la de la glucosa de 1.55 molar. Posteriormente se procedió a medir la glucosa capilar en los tiempos: 30 min, 60 min y 120 min.
Determinación del efecto a largo plazo del consumo elevado de sal en una dieta rica en carbohidratos sobre el peso en ratas Wistar
Para analizar el efecto del sodio en una dieta rica en carbohidratos sobre el peso corporal, se utilizó una dieta con la siguiente composición en nutrientes: Proteína cruda 16 %, Grasa cruda 6 %, Carbohidratos 20 %. La medición del peso se realizó en dos grupos de 7 ratas, el grupo control se alimentó con la dieta arriba indicada y el grupo problema con la misma dieta pero adicionada de sal al 7 %. El alimento fue adquirido de la empresa Nestlé México, S. A. de C. V. como croquetas, las cuales fueron trituradas utilizando un molino metálico casero, Posteriormente, la cantidad de sal que correspondía al 7 % del peso del alimento triturado (para los animales problema) fue diluido en una cantidad de agua cuyo volumen correspondía al 50 % del peso del alimento triturado (proporción porcentual volumen/peso). La cantidad de sal adicionada al alimento del 7 por ciento fue de una tercera parte del peso que correspondía a los carbohidratos de la dieta. Sin embargo, dado que el peso molecular de la glucosa (de 180.1 gramos/mol) es de aproximadamente tres veces mayor al peso molecular de la sal (NaCl con un peso molecular de 58 gramos/mol), en la dieta, la sal y la glucosa tuvieron concentraciones equimolares. Para la obtención de los pellets de alimento, el polvo alimenticio (obtenido de la trituración de las croquetas) se mezcló con el agua de manera homogénea, luego la mezcla se hizo pasar por una jeringa a la cual previamente se le trozo la parte superior con unas tijeras y de esta manera se obtuvieron pellets los cuales fueron secados a temperatura ambiente. Las ratas de cada grupo fueron marcadas y pesadas al inicio del experimento. El grupo control de ratas se alimentó con la dieta rica en carbohidratos, mientras que el grupo de ratas problema, se alimentó con la dieta rica en carbohidratos pero adicionada de sal al 7 %. Las ratas fueron pesadas antes del tratamiento y pasadas nueve semanas de someterlas al esquema de alimentación y se procedió a obtener la diferencia de peso de cada animal.
Determinación del efecto a largo plazo del consumo elevado de sal en una dieta rica en carbohidratos sobre el grosor del panículo adiposo
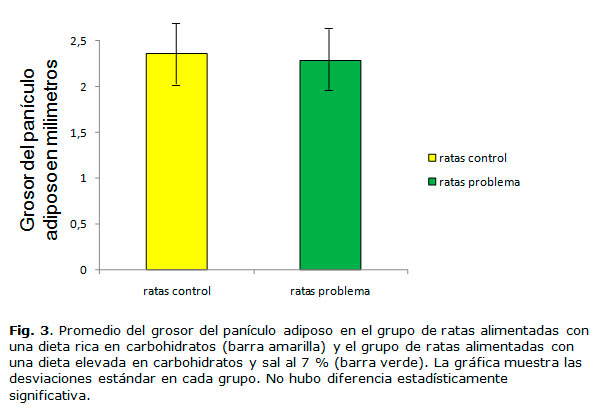
La medición del panículo adiposo en el grupo de ratas alimentadas con la dieta elevada en sodio y carbohidratos con respecto al grupo control, se realizó utilizando un plicómetro de la marca DYNATRON de la casa comercial Dynatonics Corporation. Cada grupo de ratas estuvo formado por siete animales (n=7). La medición se realizó después de nueve semanas de tratamiento.
RESULTADOS
Para corroborar nuestra hipótesis de que la sal que ingerimos en la dieta incrementa la eficiencia en el transporte de la glucosa desde la luz intestinal hasta el torrente sanguíneo, se realizaron curvas de tolerancia a la glucosa en dos grupos de 15 ratas. Uno de los grupos correspondió al grupo control y el otro al grupo problema. Ambos lotes de animales fueron sometidos a un ayuno de 12 horas, después del cual se tomó lectura de la concentración de glucosa en sangre; posteriormente se procedió a aplicar el tratamiento. A cada animal del grupo control se le administró por vía oral una solución de glucosa a una concentración 1.55 molar (1 g/kg, administrándola a un volumen de 0.8 mililitros por cada 200 gramos de peso), mientras que a los animales grupo problema se les administró también por vía oral la solución glucosada arriba descrita pero adicionada con sal a una concentración igual a la de glucosa (1.55 M). Luego de la administración de los tratamientos se realizó la medición de glucosa en sangre, a los 30 minutos, a los 60 minutos y a los 120 minutos. En la gráfica de la figura 1 podemos observar que el patrón de absorción de la glucosa en intestino fue muy similar entre ambos grupos y que no hay diferencia estadísticamente significativa. Es importante mencionar que se realizaron tres experimentos como éste de manera independiente y en todos ellos se observó el mismo resultado.
De manera paralela a la realización de los experimentos arriba descritos procedimos a evaluar el efecto de una dieta rica en carbohidratos y el consumo abundante de sal en el desarrollo de obesidad. Para ello a un grupo control de ratas se alimentó con la dieta rica en carbohidratos, mientras que a un grupo de ratas problema se le alimentó con la dieta rica en carbohidratos pero adicionada con sal al 7 %. Pasadas nueve semanas de tratamiento, las ratas de cada grupo se pesaron y se obtuvo la diferencia de peso de cada animal, el promedio de la diferencia de peso en cada grupo de animales, se comparó gráficamente, como puede apreciarse en la figura 2. Los valores de los promedios de la diferencia de peso en ambos grupos fueron prácticamente iguales. Se obtuvo un promedio de 41.45 gramos para el grupo de ratas control y de 41.52 gramos para el grupo de ratas problema. El valor de la desviación estándar para los datos de la diferencia de peso en el grupo de ratas control fue de 4.67 gramos, mientras que para el grupo de ratas tratadas con dieta hipersódica fue de 10.5 gramos. Por los valores casi iguales en los promedios y las desviaciones estándar considerables, era de esperar que no hubiera diferencia estadística significativa, se procedió entonces a realizar una prueba T de Student para confirmarlo. Como podemos observar en la figura 2, la diferencia estadística entre ambos grupos fue de p=0.98, por lo que se concluye que para este análisis no existe diferencia estadísticamente significativa.
Para finalizar el estudio se procedió a realizar la medición del panículo adiposo en el grupo de ratas alimentadas con la dieta elevada en sodio y en el grupo control al concluir nueve semanas de tratamiento. Cada grupo de ratas estuvo formado por siete animales (n=7). La medición se realizó después de dos meses de tratamiento y se utilizó un plicómetro. El promedio en el grosor del panículo adiposo en el grupo control fue de 2.36 mm y el grupo de ratas problema de 2.29 mm, la desviación estándar en el grupo problema fue de 0.37 mm y en el grupo control de 0.39 mm. Al realizar el análisis de la prueba T de Student, se observó que no había diferencia estadísticamente significativa (figura 3).
DISCUSIÓN
Durante el presente proyecto de investigación se abordó un análisis sobre el efecto que tiene el consumo de sal en el desarrollo de obesidad en ratas Wistar. La propuesta se planteó tomando en consideración que el transporte de glucosa desde la luz intestinal hacia el torrente sanguíneo es llevado a cabo por una proteína llamada SGLT1 que co-transporta al ion sodio y a la molécula de glucosa hacia el interior del enterocito para después pasar al torrente sanguíneo; también se consideró este planteamiento tomando en cuenta que la Organización Mundial de la Salud que establece que el consumo normal de sal (formada por un ion sodio y un ion cloruro) debe ser de solo 2 gramos por día y que la ingesta de sal en humanos es de 10 veces mayor que la cantidad necesaria13. En este contexto, esta propuesta de investigación, se planteó pensando en que una estrategia simple para reducir de peso sería el disminuir la cantidad de sal en los alimentos a niveles normosódicos de 2 gramos por día.
En la determinación del efecto de la dieta rica en carbohidratos y sal en el desarrollo de obesidad en la rata Wistar, se empleó una concentración de sal al 7 % ya que a esa concentración se alcanzó una relación equimolar entre las moléculas de cloruro de sodio y de glucosa, presentes en la formulación. Se verificó que a esa concentración salina fuera tolerable para su consumo. El peso de las ratas que se alimentaron con una dieta rica en carbohidratos y sodio durante nueve semanas de tratamiento se comparó con el grupo control al cual se le administró la misma dieta sin sal, el resultado mostró que no hay diferencia estadísticamente significativa en la ganancia de peso entre ambos grupos. A pesar de que el tiempo en que se realizó la determinación fue corto, no se consideró conveniente prolongar más el tratamiento debido a que el resultado de las curvas de tolerancia a la glucosa indicaron que la absorción de esta molécula en intestino es independiente de la cantidad de sal ingerida. Algo que es interesante señalar es que el consumo abundante de sal en el grupo problema incrementó considerablemente la ingesta de agua y disminuyó la del consumo de alimento, lo cual sugiere que la rata regula la homeostasis de sodio controlando su sed y alimentación15. Desafortunadamente no se midió la diferencia de la cantidad de líquidos y alimentos consumidos en cada grupo.
Con respecto a los resultados de la curva de tolerancia a la glucosa, se evaluó de qué manera la sal que se ingiere en solución junto a la glucosa favorece su absorción en el intestino. Es importante mencionar que la solución de glucosa que se administró tenía una concentración y un volumen por kilogramo de peso iguales a los que se aplican en los humanos. Los resultados de los experimentos donde se administró por vía oral la solución de sal y de glucosa en concentraciones equimolares (1.55 molar) con respecto al grupo control (donde la solución glucosada administrada no contenía sal), sugieren que existe una saturación del receptor SGLT1 por la cantidad de iones sodio que recirculan de manera natural desde el interior de los enterocitos hacía la luz intestinal15 ya que la dinámica de absorción fue similar en el grupo problema con respecto al grupo control. Es importante mencionar que se realizaron tres experimentos independientes donde observamos los mismos resultados y que no era posible incrementar al doble el número de moléculas de sal con respecto a la cantidad de moléculas de glucosa administradas debido a que la solución estaba al límite de la saturación. De acuerdo a lo reportado en la literatura, si se ingiere un volumen grande de agua, el agua bebida se mezclará con los jugos digestivos y como estos, en su mayoría, son isotónicos, la solución contenida en la luz intestinal se hará hipotónica. Al ser el intestino muy permeable al agua, habrá un pasaje de agua desde la luz a la sangre, hasta que el líquido intestinal quede isotónico. Por el contrario, si se consume sal, el líquido intestinal se hará hipertónico y el flujo osmótico será desde la sangre a Ia luz. Establecida, como primer paso, la isotonía del contenido intestinal con el plasma y el líquido extracelular, vendrán ahora Ios mecanismos de transporte que llevarán al Na+, al agua, a la glucosa, los aminoácidos, a los ácidos grasos y todas las otras sustancias desde la Iuz a la sangre16, 17. En el intestino existe una cantidad de sodio que de manera natural recircula y que es de aproximadamente de 20 a 30 gramos15. Se ha reportado que si se inhibe esta recirculación se impide la absorción de la glucosa y de aminoácidos en el intestino17, 18. Los presentes resultados sugieren que esta cantidad de sodio en recirculación garantizaría en todo momento la absorción de la glucosa en el intestino, independiemente de la ingesta de sal proveniente de la dieta.
Agradecimientos
Queremos agradecer el apoyo del estudiante Fernando Obledo Benicio por el apoyo técnico brindado en la aplicación de la sonda oral, a Salvador Elisea Quintero y Julio Villegas Leyva quienes son responsables del bioterio y nos ayudaron con la supervisión de los animales y al Doctor Valery Melnikov por los comentarios realizados el trabajo.
REFERENCIAS BIBLIOGRÁFICAS
1 World Health Organization. Controlling the global obesity epidemic. [Acceso: 8 de mayo de 2013]. Disponible en: http://www.who.int/nut/obs.htm.
2 Spiegelman BM, Choy L, Hotamisligil GS, Graves RA, Tontonoz P. Regulation of adipocyte gene expresión in differentiation and sindromes of obesity/diabetes. J Biol Chem 1993; 268: 6823-6826.
3 Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM. Positional cloning of the mouse obese gene and its human homologue. Nature 1994; 372: 425-32.
4 Hotamisligil GS, Shargill NS, Spiegelman BM. Adipose expression of tumor necrosis factor-a: direct role in obesity-linked insulin resistance. Science 1993; 259: 87-91.
5 Shimomura I, Funahashi T, Takahashi M, Maeda K, Kotani K, Nakamura T, et al. Enhanced expression of PAI-1 in visceral fat : Possible contributor to vascular disease in obesity. Nat Med 1996; 2: 800-3.
6 Maeda K, Okubo K, Shimomura I, Mizuno K, Matsuzawa Y, Matsubara K, et al. analysis of an expression profile of genes in the human adipose tissue. Gene 1997; 190: 227-35.
7 Steppan CM, Bailey ST, Bhat S, Brown EJ, Banerjee RR, Wright CM, et al. The hormone resistin links obesity to diabetes. Nature 2001; 409: 207-12.
8 Maeda K, Okubo K, Shimomura I, Funahashi T, Matsuzawa Y, Matsubara K. cDNA cloning and expression of a novel adipose-specific collagen-like factor, apMI (adipose most abundant gene transcript 1). Biochem Biophys Res Commun 1996; 221: 286-289.
9 Roden M, Bernroider E. Hepatic glucose metabolism-its role in health and disease. Best Pract Res Clin Endocrinol Metab 2003; 17(3):365-383.
10 Reuss L: One-hundred years of injury: the mechanism of glucose absorption in the intestine. Annu Rev Physiol 2000;62:939.
11 Ferraris RP, Diamond J. Regulation of intestinal sugar transport. Physiol Rev 1997;77(1):257-302.
12 Depósitos de documentos de la FAO. Departamento de agricultura. Nutrición humana en el mundo en desarrollo. [Acceso: 8 de mayo de 2013]. Disponible en: http://www.fao.org/DOCREP/006/W0073S/w0073s0d.htm
13 Reunión Técnica de la OMS sobre la Reducción del Consumo de Sal en la Población. El cloruro sódico alimentario: efectos adversos. París. ISBN 978 92 4 159537 7 (Clasificación de la NLM: QU 145), 2006.
14 Flowers MT, Ntambi JM. Stearoyl-CoA Desaturase and its Relation to High-Carbohydrate Diets and Obesity. Biochim Biophys Acta 2009; 1791(2): 85-91.
15 Guyton AC, Hall JE. Tratado de fisiología médica. 11ª ed. Madrid: Elsevier; 2006.
16 Zeuthen T, Stein WD. Cotransport of salt and water in membrane proteins: membrane proteins as osmotic engines. J Membr Biol 1994;137(3):179-95.
17 Montoreano R. Manual de Fisiología y biofísica para estudiantes de medicina. [Acceso: 8 de mayo de 2013]. Disponible en: http://www.fundabiomed.fcs.uc.edu.ve/cap51.pdf
18 Field M. Regulation of active ion transport in the small intestine. Ciba Found Symp. 1976;(42):109-27.
Recibido: 5 de julio de 2013.
Aceptado: 5 de agosto de 2013.
Joel Cerna Cortés . Facultad de medicina de la Universidad de Colima. Av. Universidad #333. Colonia Las Víboras. C.P. 28040. Colima, Col., México. Tel: 312 3161099. Fax (52) 312 3161099. Email: joelcerna@ucol.mx

{kind=link}
{kind=link}
{kind=link}