










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl vol.32 no.2 La Habana abr.-jun. 2015
REPORT
Protective capacity of vaccine candidates against dengue virus type 2 based on the capsid protein and the oligodeoxynucleotide 39M
Capacidad protectora de candidatos vacunales contra el virus dengue serotipo 2 basados en la proteína de la cápsida y el oligodesoxinucleótido 39M
Ernesto Marcos1, Lázaro Gil1, Lisset Hermida1, Alienys Izquierdo2, Gerardo Guillén1, María Guadalupe Guzmán2, Laura Lazo1, Iris Valdés1, Mayling Álvarez2, Edith Suzarte1, Rosa Ramírez2, Jorge Castro1, Yusleidi Pérez1, Karem Cobas1, José Ángel Silva1, Yaremis Romero1, Karelia Cosme1, Emma Brown1, Aracelys Blanco1, Jamilet Miranda1, Yasser Perera1
1 Centro de Ingeniería Genética y Biotecnología. Ave. 31 e/ 158 y 190, Cubanacán, Playa, PO Box 6162, CP 10600, La Habana, Cuba.
2 Tropical Medicine Institute Pedro Kourí, IPK. Autopista Novia del Mediodía, Km 6½, La Lisa, CP 11400, La Habana, Cuba.
ABSTRACT
Vaccine formulations of the chimeric DIIIC-2 protein with different ODNs displaying adjuvant capacity were tested in mice against dengue virus serotype 2. DIIIC-2 comprises domain III from the envelope protein fused to the N-terminal region of the capsid protein. One of the ODNs, named ODN 39M, combines CpG motifs described as immunostimulatory active in mice, monkeys and human cells. The ODN sequence determined the immune response pattern generated, since ODN 39M was the only ODN inducing a Th1 response similar to that induced by the viral infection in mice. Hence, ODN 39M was aggregated with either the capsid protein of dengue-2 (C-2) or the DIIIC-2 protein, and both formulations were adjuvanted in alum. Upon administration in two mice models and in non-human primates, both formulations induced protection after the viral challenge. This was the first demonstration of the protective capacity induced by the vaccine formulation 39M-DIIIC-2 in non-human primates and in transgenic mouse models which develop dengue-like clinical symptoms. Moreover, viremia levels were reduced in non-human primates with the 39M-C-2 formulation. This was the first evidence ever on the induction of a functional immune response against dengue virus by vaccination in non-human primates, regardless the presence of neutralizing antibodies. Moreover, this was the first work describing protection in monkeys with a formulation that is suitable for human use and combines recombinant proteins from dengue virus with alum. This research granted the 2014 Award of the Cuban National Academy of Sciences.
Keywords: Dengue-2 virus, capsid protein, nonhuman primates, protection.
RESUMEN
Se evaluaron preparaciones de la proteína quimérica DIIIC-2 con diferentes oligodesoxinucleótidos (ODN) con actividad adyuvante, contra el virus del dengue serotipo 2 en ratones. DIIIC-2 comprende el dominio III de la envoltura del virus dengue serotipo 2 fusionado al extremo N-terminal de la proteína de la cápsida del mismo serotipo. Uno de los ODN, el 39M, combina en una misma molécula regiones CpG descritas como inmunológicamente activas en células de ratones, monos y seres humanos. La secuencia del ODN determinó el patrón de respuesta inmune generado, pues solo el ODN 39M indujo un patrón de respuesta Th1 semejante al que se induce tras la infección viral en ratones. Basado en este hallazgo, se seleccionó el ODN 39M y se agregó con las proteínas DIIIC-2 y la de la cápsida del dengue serotipo 2 (C-2), ambas formulaciones adyuvadas en alúmina. Ambas formulaciones indujeron protección frente al reto viral en dos modelos animales de ratones y en primates no humanos. Fue la primera vez que se demuestra la capacidad protectora de la preparación 39M-DIIIC-2 tanto en ratones transgénicos que desarrollan síntomas similares al dengue como en primates no humanos. Además, la reducción de los niveles de viremia alcanzados en primates no humanos con la preparación 39M-C-2 evidenció por primera vez, en este modelo animal, la funcionalidad de una respuesta inmune independiente de anticuerpos neutralizantes contra el virus dengue. Además, este fue el primer reporte de protección en monos con una formulación adecuada para ser administrada a seres humanos, que combina a proteínas recombinantes del virus dengue con alúmina. Este trabajo mereció el Premio Anual de la Academia de Ciencias de Cuba para el año 2014.
Palabras clave: virus Dengue 2, proteína de la cápsida, primates no humanos, protección.
INTRODUCTION
Dengue virus (DENV) infection is a major emerging disease of tropical and subtropical countries, transmitted by the bite of infected mosquitoes. Each of the four DENV serotypes causes a spectrum of clinical symptoms ranging from mild fever to potentially fatal manifestations of dengue shock syndrome.
Our group has developed subunit vaccine candidates based on the capsid protein of DENV-2 (C-2) and a fusion of this protein with the domain III of the envelope protein (E) of the homologous serotype (DIIIC-2). The resulting recombinant proteins were efficiently produced in Escherichia coli and then properly purified and folded. When presented as a particulate aggregate, incorporating random oligodeoxynucleotides of 45 bases (45b-ODNs), DIIIC-2 induced antiviral and neutralizing antibodies, cell-mediated immunity (CMI) and significant protection in mice [1].
In the last years a high number of ODNs with adjuvant capacity have been identified. In 2001, Krug et al. identified the ODN 2216 (5´-GGGGGACGATCGTCGGGGGG-3´), which has the capacity to induce IFNα/β/γ secretion in human peripheral blood mononuclear cells (PBMC) and mice splenocytes [2]. Verthelyi et al. identified the ODN K3 (5´-ATCGACTCTCGAGCGTTCTC-3´) which showed the induction of proliferation of humans B cells and the stimulation of IgM and IL-6 secretion [3]. In addition, the adjuvant capacity of the preparation IC31, a combination of a polycationic peptide and the ODN1a (5´-ICICICICICICICICICICICICIC-3´) has evaluated [4]. ODN1a (named PolyIC) is a synthetic ODN of 26 bases composed by 13 copies of the dinucleotide Inosine/Cytosine (IC) joined by a phosphodiester bond. The preparation IC31 was able to potentiate the specific cellular and humoral immune response induced in mice by an ovalbumin peptide and murine tyrosinase [4].
Here we present results on the adjuvant effect of a modified version of the ODN m2216, the ODN PolyIC (without chemical modifications) and the 39M ODN, formed by the fusion of the ODNs m2216 and K3, on the recombinant protein DIIIC-2. After the selection of the most immunogenic variant in mice, its ability to induce a protective response was evaluated in mice and monkeys for aggregates of DIIIC-2 with C-2 and the selected ODNs. This research granted the 2014 Award of the Cuban National Academy of Sciences.
RESULTS
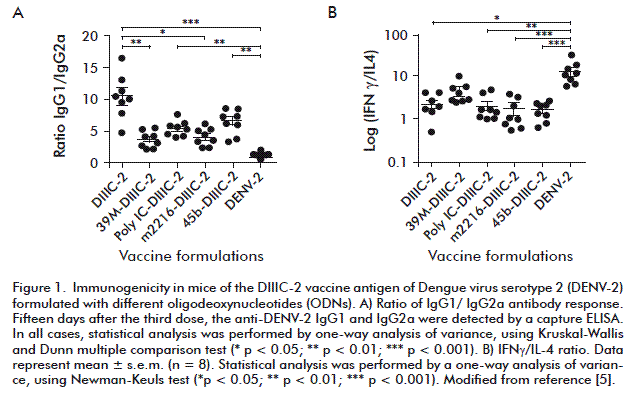
The anti-DENV-2 immunoglobulin (Ig)G1/IgG2a ratio was assessed in mice after the aggregation of the protein with the three different ODNs (m2216, poly IC and 39M) (Figure 1). A group of animals receiving the non-aggregated DIIIC-2 was included as a control (without ODNs). As additional controls, three groups were tested: one received a DIIIC-2 protein aggregated with 45b ODN (45b-DIIIC-2), a placebo formulation (negative control) and the last one was inoculated with 105 plaque-forming units (p.f.u.) of infective DENV-2 (positive control). No statistical differences were observed in the antibody responses among the groups of animals inoculated with either of DIIIC-2 aggregates (p > 0.05) (Figure 1A). However, only the IgG1/IgG2a ratios from the animals receiving 39M-DIIIC-2, m2216-DIIIC-2 or infective DENV-2 (positive control) were statistically lower than those observed in the group immunized with the non-aggregated DIIIC-2 (p < 0.05).
On the other hand, culture supernatants from mock-treated or DENV-2-infected splenocytes from immunized animals were tested by ELISA to measure the concentrations of IFN-γ and interleukin (IL)-4 (Figure 1B). The IFN-γ/IL-4 ratios were higher in animals inoculated with 39M-DIIIC-2 or DENV-2, with significant differences as compared to the other groups (p < 0.05). These results were in accordance with the low ratios of IgG1/IgG2a observed in DIIIC-2 plus 39M and DENV-2 immunized animals.
Taking into account these results a second experiment was performed to evaluate the protective capacity induced by the protein aggregates containing the ODN 39M. Four groups were included, the animals immunized with: 1) protein 39M-DIIIC-2; 2) C-2-39M; 3) placebo formulation (negative control); 4) infective DENV-2 (positive control).
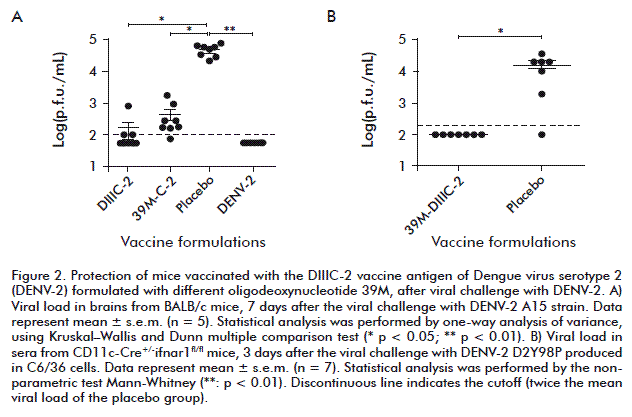
One month after the last dose, protection was assessed using the mouse model of dengue encephalitis. Animals were intracranially inoculated with 50 median lethal doses (LD50) of the live, neuroadapted DENV-2 strain A15. Eight animals from each immunized group were euthanized to measure viral load in the brain. As shown in Figure 2A, the highest viral titers (104.5 p.f.u./mL) were detected in the brain of animals from the placebo group. No brain infection was observed in the positive control group.
The statistical analysis revealed significant differences between the C-2 and DIIIC-2 immunized animals (p < 0.05) or DENV-2 group (p < 0.01) as compared to placebo, respectively.
Therefore, the protective capacity of the formulation 39M-DIIIC-2 was evaluated in an immunocompromised mouse model CD11c+-Cre+/-ifnar1fl/fl, where after the inoculation of non-mouse-adapted dengue viral strains results in systemic dengue infection (Figure 2B). Viral load was highly significantly reduced in mice immunized (lower than 102.3 p.f.u./mL; p < 0.01) in respect to the placebo group (104.2 p.f.u./ mL) was observed.
On the basis of the preclinical studies in mice, the DIIIC-2 aggregates were evaluated in non-human primates. Green monkeys were divided into three groups, receiving: 1) 39M-DIIIC-2 formulation; 2) 39M-C-2; 3) placebo (alum plus ODN 39M). Administrations were performed once every 2 months for 6-months.
To assess the protective efficacy of the recombinant vaccine candidate, all animals were challenged by subcutaneous inoculation of 103 p.f.u. of DENV- 2. Serum samples were collected after challenge and viremia was quantified using Vero cells. All the placebo animals developed viremia with a mean duration of four days and maximum viral titers of 102.5, 101.4 and 102.4 p.f.u./mL (Figure 6A in reference [5]). On the contrary, two out of three animals in the group receiving DIIIC-2 plus 39M were totally protected, whereas the animal M2140 (partially protected) developed a brief, low-level viremia (titer lower than 10 p.f.u./mL) that lasted only 1 day (Figure 6B in reference [5]).
In turn, variable levels of viremia were detected in the group immunized with 39M-C-2 (Figure 3B in reference [6]). The animal M2072 had five days of viremia with a maximum virus titer of 101.9 p.f.u./mL, whereas the monkey M2126 exhibited three days of viremia with maximum virus titer of 101.6 p.f.u./ mL. The monkey M2048 was completely protected. Statistical analysis of mean viremia using a nonparametric Mann-Whitney test showed high statistical differences between this group and placebo (p = 0.0077).
DISCUSSION
In this study, we immunized mice with formulations containing ODNs [2, 3] that had been previously reported to have adjuvant capacity in mice, monkeys and human cells. In addition, we evaluated the novel ODN 39M, which contains CpG motifs and has potential immunostimulatory properties.
As expected, our results revealed the influence of the ODN sequence on the immunogenicity of DIIIC-2, as indicated by the subclass of IgG elicited and by the secretion of IFN-γ in mice. The addition of ODNs to the formulations shifted the humoral immune responses toward a Th1 phenotype, despite using alum as adjuvant. This effect was more pronounced with the formulations containing the ODNs 39M and m2216. In turn, immunization with the formulation containing the ODN 39M also induced a high-level secretion of IFN-γ and resulted in the highest IFN-γ/IL-4 ratio. These findings indicate that the overall magnitude of both, humoral and cellular immune responses is affected by the ODN used for protein aggregation.
The protection assay in BALB/c mice was then carried out with the 39M-DIIIC2 and 39M-C2 formulations. In both cases, there was a significant reduction in virus titers in the brain of challenged mice. The formulation 39M-DIIIC2 was also evaluated in an immunocompromised mouse model CD11c+-Cre+/- ifnar1fl/fl. This animal model has the disadvantage that use immunocompromised animals with a limited capacity of develop an efficient immune response after the vaccination. Noteworthy, the main advantage of this model is that it is possible to generate a systemic infection in mice, with a non-neuroadapted dengue strain. There was a significant reduction in virus titers in sera of challenged mice.
The successful results obtained in mice with 39M-DIIIC-2 and 39M-C-2 aggregates encouraged us to conduct experiments in monkeys using these two formulations. The immunogenicity of DIIIC-2 in monkeys was assessed by a protection assay. Two monkeys were fully protected, whereas the third one only had a 1 day with low-titer viremia. This was the first work describing protection with a formulation that is suitable for human use and combines a recombinant protein from DENV with alum. In our previous studies, protection was only provided by proteins emulsified in Freund’s adjuvant, whereas the formulations containing alum did not conferred protection [7, 8]. This was also reported by Simmons et al. [9], showing no protection in monkeys receiving the protein MBP-domain III formulated with alum. Conversely, a promising subunit vaccine candidate based on the E protein and developed by Hawaii Biotech provided solid protection when combined with ISCOMATRIX and protein NS1 in a tetravalent formulation [10]. Nevertheless, when alum was used without any other Th1-prone adjuvancity in a monovalent formulation, none of the immunized monkeys was protected [10].
The formulation 39M-C-2 induced a reduction of viral load after the viral challenge. Capsid protein does not contain a high quantity of cell-mediated immunity (CMI) epitopes for monkeys. In fact, a very recent study revealed in detail that the dengue capsid is not the most immunodominant region for CMI in humans, since it contains one region immunologically identified by 4 % of responders as compared to other viral antigens recognized by high magnitude T-cell responses [11].
Both, the capsid protein and domain III from the envelope protein are serotype specific viral fragments, so, this supports the development of a tetravalent vaccine formulation, able to protect against the four dengue serotypes.
CONCLUSIONS
The experiments conducted in this work using mice and monkeys provide the proof of concept of a subunit vaccine based on the capsid protein and the ODN 39M, which is effective in both animal models using alum as adjuvant. Further studies should carefully measure the duration of the functional immune response. This issue should be addressed in upcoming experiments in non-human primates and humans, using a tetravalent formulation.
REFERENCES
1. Valdes I, Bernardo L, Gil L, Pavon A, Lazo L, Lopez C, et al. A novel fusion protein domain III-capsid from dengue-2, in a highly aggregated form, induces a functional immune response and protection in mice. Virology 2009;394:249-58.
2. Krug A, Rothenfusser S, Hornung V, Jahrsdorfer B, Blackwell S, Ballas ZK, et al. Identification of CpG oligonucleotide sequences with high induction of IFN-alpha/ beta in plasmacytoid dendritic cells. Eur J Immunol 2001;31:2154-63.
3. Verthelyi D, Ishii KJ, Gursel M, Takeshita F, Klinman DM. Human peripheral blood cells differentially recognize and respond to two distinct CPG motifs. J Immunol 2001;166:2372-7.
4. Schellack C, Prinz K, Egyed A, Fritz JH, Wittmann B, Ginzler M, et al. IC31, a novel adjuvant signaling via TLR9, induces potent cellular and humoral immune responses. Vaccine 2006;24:5461-5472.
5. Gil L, Marcos E, Izquierdo A, Lazo L, Valdes I, Ambala P, et al. The protein DIIIC-2, aggregated with a specific oligodeoxynucleotide and adjuvanted in alum, protects mice and monkeys against DENV- 2. Immunol Cell Biol. 2015;93(1):57-66.
6. Gil L, Izquierdo A, Lazo L, Valdes I, Ambala P, Ochola L, et al. Capsid protein: evidences about the partial protective role of neutralizing antibody-independent immunity against dengue in monkeys. Virology. 2014;456-457:70-6.
7. Hermida L, Bernardo L, Martin J, Alvarez M, Prado I, Lopez C, et al. A recombinant fusion protein containing the domain III of the dengue-2 envelope protein is immunogenic and protective in nonhuman primates. Vaccine 2006;24:3165-71.
8. Bernardo L, Izquierdo A, Alvarez M, Rosario D, Prado I, Lopez C, et al. Immunogenicity and protective efficacy of a recombinant fusion protein containing the domain III of the dengue 1 envelope protein in non-human primates. Antiviral Res 2008;80:194-9.
9. Simmons M, Porter KR, Hayes CG, Vaughn DW, Putnak R. Characterization of antibody responses to combinations of a dengue virus type 2 DNA vaccine and two dengue virus type 2 protein vaccines in rhesus macaques. J Virol 2006;80:9577- 85.
10. Clements DE, Coller BA, Lieberman MM, Ogata S, Wang G, Harada KE, et al. Development of a recombinant tetravalent dengue virus vaccine: immunogenicity and efficacy studies in mice and monkeys. Vaccine 2010;28:2705-15.
11. Weiskopf D, Angelo MA, de Azeredo EL, Sidney J, Greenbaum JA, Fernando AN, et al. Comprehensive analysis of dengue virus-specific responses supports an HLA-linked protective role for CD8+ T cells. Proc Natl Acad Sci USA. 2013;110:E2046-53.
Received in January, 2015.
Accepted in May, 2015.
Ernesto Marcos. Centro de Ingeniería Genética y Biotecnología. Ave. 31 e/ 158 y 190, Cubanacán, Playa, PO Box 6162, CP 10600, La Habana, Cuba.


{kind=link}
{kind=link}