










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Producción Animal
versión On-line ISSN 2224-7920
Rev. prod. anim. vol.30 no.3 Camagüey set.-dic. 2018
GENÉTICA Y REPRODUCCIÓN
Efectos sobre la fertilidad del cerdo de dos candidatos vacunales recombinantes basados en la GnRH
Effects of Two GnRH-based Recombinant Vaccine Candidates on Pig Fertility
Roberto Basulto Baker*, Gerardo Guillén Nieto**, Lesvia Calzada Aguilera*, Jesús Junco Barranco*, Nelson Izquierdo Pérez***, Osvaldo Reyes Acosta**, Hilda Garay Pérez**, Ricardo Bringas Pérez**, Eulogio Pimentel Vázquez*, José A. Bertot***, Rafael Avilés***, María Dolores Castro Santana*, Eddy Bover Fuentes*, Franklin Fuentes Aguilar*
* Centro de Ingeniería Genética y Biotecnología (CIGB), Camagüey, Cuba
** Centro de Ingeniería Genética y Biotecnología (CIGB), La Habana, Cuba
*** Facultad de Ciencias Agropecuarias, Universidad de Camagüey Ignacio Agramonte Loynaz, Camagüey, Cuba. roberto.basulto@cigb.edu.cu
RESUMEN
En este trabajo se describe, por primera vez, la obtención de la fimbria híbrida K88ab-GnRH, la fusión de la GnRH a la proteína P64k de N. meningitidis (P64k-GnRH) y su evaluación como candidatos vacunales para controlar la fertilidad en mamíferos. Veinte cerdos machos híbridos fueron asignados al azar a cuatro grupos: placebo e inmunizados con K88ab-GnRH, P64k-GnRH y con un péptido análogo de GnRH (GnRHm1), unido a un epítopo T cooperador del toxoide tetánico (TT) (control positivo), respectivamente. Los cerdos se inmunizaron con 9-10 semanas de edad, en un esquema de 2 dosis y se sacrificaron 16 semanas después. K88ab-GnRH, P64k-GnRH y GnRHm1-TT indujeron niveles de testosterona en suero, mayor, similar y menor, comparados con el placebo, respectivamente. En el grupo K88ab-GnRH los cerdos disminuyeron (P 0,01) el largo y peso de los testículos y el peso de los epidídimos, comparado con el placebo y ninguno llegó a eyacular. En el grupo P64k-GnRH los cerdos disminuyeron el peso de los testículos (P 0,05), y sólo uno llegó a eyacular. Los testículos de los cerdos inmunizados con K88ab-GnRH y P64k-GnRH mostraron daños estructurales, funcionales y afectación de la espermatogénesis. Las glándulas sexuales accesorias del grupo P64k-GnRH estaban normales a diferencia de K88ab-GnRH, en las que se observó fibrosis intersticial. Los daños provocados por K88ab-GnRH y P64k-GnRH en los órganos diana evaluados, resultaron inferiores en todos los casos, a las afectaciones que generó el péptido GnRHm1-TT.
Palabras clave: GnRH, K88ab, P64k, inmunocastración, proteína portadora.
ABSTRACT
This paper describes for the first time the generation of the K88ab-GnRH hybrid fimbriae, the fusion of N. meningitidis P64k protein (P64k-GnRH), and its evaluation as vaccine candidates to control fertility in mammals. Twenty hybrid male pigs were randomly distributed in four groups: placebos and immunized with K88ab-GnRH, P64k-GnRH and a GnRH analogue (GnRHm1), linked to a tetanus toxoid (TT) T-helper epitope (positive control), respectively. The pigs were immunized at 9-10 weeks of age, using a two-dose scheme, and were sacrificed sixteen weeks later. K88ab-GnRH, P64k-GnRH, and GnRHm1-TT induced higher, similar, and lower testosterone levels in the serum, compared to the placebo, respectively. In the K88ab-GnRH group, the pigs underwent a reduction in testicle size and weight (P 0.01), and the weight of epididymes compared to the placebo; none of them was able to ejaculate. In the P64k-GnRH group, the pigs had a reduction in testicle weight (P 0.05), and only one of them was able to ejaculate. The testicles of the pigs immunized with K88ab-GnRH and P64k-GnRH showed structural and functional damage; spermatogenesis was also affected. The accessory sexual glands of the P64k-GnRH group were normal, in contrast to K88ab-GnRH, where interstitial fibrosis was observed. The damage caused by K88ab-GnRH and P64k-GnRH in the target organs evaluated were in all cases lower than the affectations caused by the GnRHm1-TT peptide.
Keywords: GnRH, K88ab P64k, immunocastration, carrier protein.
INTRODUCCIÓN
La hormona liberadora de gonadotropinas (GnRH) es una molécula estructuralmente conservada y clave en el control de la reproducción de los mamíferos (Millar, Pawson, Morgan, Rissman y Lu, 2008). La inmunocastración, basada en provocar una respuesta inmune contra la GnRH, constituye una alternativa a la castración quirúrgica, más apropiada, menos traumática y reversible (Andersson, Brunius, Zamaratskaia y Lundström, 2012). Sin embargo, la GnRH es un hapteno y un auto antígeno, por lo que para obtener anticuerpos contra esta molécula se debe, entre otras estrategias, generar una o múltiples copias de péptidos análogos de GnRH que puedan ser vistos como no propios por el sistema inmune y acoplarlos a proteínas transportadoras como el toxoide tetánico, toxoide de la difteria o sus epítopos sintéticos T cooperadores (Fuentes et al., 2017). Otra de las estrategias empleadas es la de fusionar la GnRH a la fimbria P y expresarla en E. coli K-12 (Van der Zee et al., 1995) o a la fimbria de Dichelobacter nodosus y expresarla en Pseudomonas aeruginosa (Mott y Reilly, 1994).
En los animales de granja, la vacunación contra la GnRH podría ser una alternativa para reducir la agresividad y prevenir la conducta sexual en animales no castrados, la preñez y cambios en el olor y sabor de la carne en cerdos, equinos, bovinos y ovino-caprinos (Stevens et al., 2005; Janett, Stump, Burger y Thun, 2009; Han et al., 2015; Mancini, Menozzi y Filippo, 2017; Yamsakul et al., 2017). En mascotas y especies de vida silvestre una vacuna contra la GnRH podría evitar la superpoblación y el comportamiento sexual no deseado (Basulto, et al., 2003; Levy, Friary, Miller, Tucker y Fagerstone, 2011).
En este trabajo se describe, por primera vez, la clonación de la GnRH de mamíferos en el gen que codifica para la subunidad mayor de la fimbria K88ab y en el gen para la dihidrolipoamida dehidrogenasa (lpdA) o proteína P64k de la bacteria N. meningitidis. Además, su evaluación como candidatos vacunales para la inmunocastración de mamíferos, en cerdos machos prepúberes como modelo animal.
MATERIALES Y MÉTODOS
Cepas bacterianas, plasmidios y medios de cultivo
Se emplearon cepas de E. coli K-12 y medios de cultivo, suplementados con ampicilina ![]() para la selección de los plasmidios. Se utilizaron los plasmidios pK203 (Fig. 1A), los cuales contienen los genes estructurales para la biosíntesis y expresión de la fimbria K88ab (Bakker et al., 1992), insertados en el sitio Hind III del vector pUC19 y el pM-6 (Silva et al., 1992), que comprende el gen codificante para la lpdA o proteína P64k de N. meningitidis (Guillén et al., 1998), bajo el promotor triptófano, clonado en el vector pUC-18, obtenidos en los Centros de Ingeniería Genética y Biotecnología de Camagüey y La Habana.
para la selección de los plasmidios. Se utilizaron los plasmidios pK203 (Fig. 1A), los cuales contienen los genes estructurales para la biosíntesis y expresión de la fimbria K88ab (Bakker et al., 1992), insertados en el sitio Hind III del vector pUC19 y el pM-6 (Silva et al., 1992), que comprende el gen codificante para la lpdA o proteína P64k de N. meningitidis (Guillén et al., 1998), bajo el promotor triptófano, clonado en el vector pUC-18, obtenidos en los Centros de Ingeniería Genética y Biotecnología de Camagüey y La Habana.
Diseño y síntesis del péptido GnRHm1-TT
El péptido GnRHm1-TT (C-QHWSYPLRPGGGQYIKANSKFIGITEL-NH2), que incluye la variante modificada de la GnRH de mamíferos (Bringas, Basulto, Reyes y de la Fuente, 1998), se obtuvo en el CIGB Habana, mediante síntesis en fase sólida con el empleo de la química Fmoc/tBu (Fields y Noble, 1990).
Clonación de ADN
Fue digerido pK203 con la endonucleasa de restricción Eco RI y religado con T4 ADN ligasa, con el fin de disminuir su talla al eliminar el fragmento Eco RI (2) - Eco RI (3) (Fig. 1A). El plasmidio resultante pK333 contiene los genes faeA-H del determinante genético K88ab (Fig. 1A). La digestión de pK333 con Bss HII (Fig. 1A) permitió reemplazar la región hipervariable de la subunidad fimbrial faeG (Fig. 1B), por el adaptador sintético de 70 pares de bases que codifica para la GnRH (Fig. 1C) y generar el plasmidio pK365. Fue digerido pM6 con Mlu I para introducir los oligonucleótidos codificantes para GnRH (5´-CGCGCAGCACTGGTCCTATGGACTGCGCCCTGGAGA-3´ y 3´-GTCGTGACCAGGATACCTGACGCGGGACCTCTGCGC-5´) en el gen de la lpdA. El plasmidio obtenido se nombró p64k-GnRH.
Expresión y purificación de la fimbria K88ab-GnRH y de la proteína recombinante P64k-GnRH
El precipitado con las células de E. coli transformadas con pK365 se resuspendió en tampón fosfato salino (PBS, por sus siglas en inglés) 1X (137 mM NaCl; 2,7 mM KCl; 10 mM Na2HPO4; 1,8 mM KH2PO4; pH 7,2) e incubó durante 20 min a 60° C, se centrifugó y tomó con cuidado el sobrenadante. La ruptura de las bacterias transformadas con p64k-GnRH se logró con un homogenizador ultrasónico (Labsonic U, B. Braun, Alemania). El sedimento, en PBS 1X, se precipitó con sulfato de amonio al 40 y 60 %, centrifugó, se resuspendió y dializó contra PBS 1X, durante 48 h . En ambos casos las proteínas de interés se conservaron a -20 C hasta que fue requerido.
Detección inmunológica de la fimbria K88ab y de la proteína P64k
La fimbria K88ab se detectó con un sistema ELISA que emplea el anticuerpo monoclonal CBC-K88a1 (Castro et al., 2004). La expresión de la proteína P64k se detectó mediante la inmuno-identificación de colonias y Western-blot con el anticuerpo monoclonal anti P64k AcM 114-HRP (Guillén et al., 1998).
Determinación de proteínas
La determinación de las proteínas totales se realizó por el método de Lowry. Las proteínas K88ab-GnRH y P64k-GnRH se cuantificaron mediante electroforesis de proteínas (SDS-PAGE) y densitometría óptica.
Animales
Un total de veinte cerdos machos híbridos fueron asignados al azar a cuatro grupos experimentales. Los cerdos fueron alojados en instalaciones propias para ellos y manejados de acuerdo al manual de crianza porcina de Cuba.
Inmunización de los cerdos contra GnRH
En la elaboración de las vacunas se empleó 0,4 mg de cada antígeno. En todos los casos las proteínas fueron resuspendidas en PBS 1X y emulsificadas con el adyuvante completo de Freund (ACF) (1:1), momentos antes de la vacunación. Los cerdos entre 9 y 10 semanas de edad y un peso corporal promedio de 11,5 kg, fueron inmunizados intramuscularmente en el cuello con 2 mL de vacuna y recibieron una segunda dosis en un intervalo de 8 semanas con la misma composición, pero esta vez emulsificada con adyuvante incompleto de Freund (AIF). Un grupo de animales fue vacunado solamente con PBS 1X y ACF o AIF y se utilizó como placebo. Otro se vacunó con GnRHm1-TT y se utilizó como control positivo. Los restantes se inmunizaron con K88ab-GnRH y P64k-GnRH, respectivamente.
Evaluaciones in vivo y post mortem
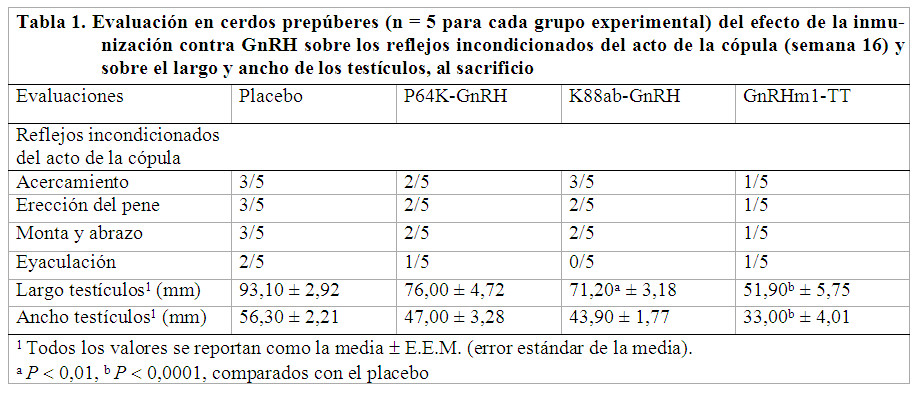
A partir de la primera inmunización y cada 4 semanas se realizó la medición de los testículos con un pie de rey. En la semana 16 se tomaron muestras de sangre, por punción del seno retro-orbital, para la extracción del suero y posterior determinación de los niveles de testosterona (Fuentes et al., 2017). También se comprobó el desarrollo de los reflejos incondicionados del acto de la cópula: acercamiento y preparación sexual, erección del pene, monta y abrazo, búsqueda para la introducción del pene en la vagina, fricción, eyaculación y desmonta. Estos reflejos se evaluaron de bien (1) cuando se evidenciaron de manera convincente, y de mal (0), cuando no se produjeron, al menos en una de las tres evaluaciones realizadas para cada reflejo. La estimulación de los verracos se realizó con una cerda en celo natural. Los cerdos se sacrificaron al finalizar la semana 16, según la norma cubana. Los epidídimos, próstatas, vesículas seminales y glándulas bulbo-uretrales fueron extirpados y pesados. Los testículos fueron medidos y pesados. Para el examen histológico se les tomaron muestras de las secciones transversales y sagitales. La tinción se realizó por el método de hematoxilina y eosina.
Análisis estadístico
Se utilizó un ANOVA de una vía y la prueba de comparaciones múltiples de Tukey, en el programa GraphPad Prism 6 para Windows, versión 6.01. Valores de P 0,05 se consideraron estadísticamente significativos.
RESULTADOS Y DISCUSIÓN
La inserción de la GnRH en pK365 disminuyó en 81,3 % la producción de la fimbria K88ab, comparado con pK333; sin embargo, en pM6 no afectó la producción de la proteína P64k (estos datos no se muestran).
En todos los cerdos se observó que, después de la primera inmunización, sus testículos siguieron creciendo, al igual que los del grupo placebo. Sólo después de concluido el esquema de inmunización se observó que los testículos de los cerdos vacunados contra GnRH disminuyeron su tamaño.
Los cerdos inmunizados con K88ab-GnRH tuvieron un valor medio de testosterona de 16,71 ± 8,53 nmol/L, mayor comparado con el placebo (8,91 ± 3,71 nmol/L). Los grupos P64k-GnRH y GnRHm1-TT tuvieron valores de testosterona de 8,47 ± 5,23 nmol/L y 1,18 ± 0,92 nmol/L, similar y menor comparados con el placebo, respectivamente.
La reducción de los niveles séricos de testosterona debe haber inducido la disminución significativa del tamaño y peso de testículos y glándulas accesorias de los animales inmunizados contra GnRH. El grupo K88ab-GnRH tuvo un valor medio de testosterona mayor, comparado con el placebo. En este grupo, tres cerdos con valores de testosterona > 10,40 nmol/L, lograron los reflejos de acercamiento, y dos de ellos la monta, erección y penetración de la vagina de la cerda (Tabla 1). Los dos restantes tuvieron valores de testosterona 7,50 nmol/L, que coinciden con los animales que no lograron desarrollar los reflejos incondicionados de la cópula (Tabla 1). Dos de los cerdos que se inmunizaron con P64k-GnRH tuvieron erección y monta, y sólo uno eyaculó. En el grupo GnRHm1-TT cuatro cerdos tuvieron valores de testosterona ![]() 0,95 nmol/L, de ellos, tres
0,95 nmol/L, de ellos, tres ![]() 0,07 nmol/L; o sea, por debajo del nivel de castración para esta especie animal (0,4 nmol/L), resultando inmunocastrados. Sólo uno tuvo un valor de 4,79 nmol/L, coincidiendo con el único cerdo de este grupo experimental que desarrolló todos los reflejos primarios de la cópula (Tabla 1).
0,07 nmol/L; o sea, por debajo del nivel de castración para esta especie animal (0,4 nmol/L), resultando inmunocastrados. Sólo uno tuvo un valor de 4,79 nmol/L, coincidiendo con el único cerdo de este grupo experimental que desarrolló todos los reflejos primarios de la cópula (Tabla 1).
En el grupo K88ab-GnRH tres cerdos mostraron el reflejo de acercamiento y excitación sexual, dos de estos tuvieron erección y monta de la cerda, pero ninguno llegó a eyacular. En el grupo GnRHm1-TT sólo uno mostró todos los reflejos incondicionados de la cópula (Tabla 1). Un cerdo del grupo P64k-GnRH y otro de GnRHm1-TT se mostraron agresivos en presencia de la cerda en celo y no mostraron el reflejo de acercamiento y excitación sexual. Los cerdos inmunizados contra GnRH mostraron disminución de los reflejos incondicionados de la cópula, más notable en el grupo GnRHm1-TT, en correspondencia con la reducción del peso y tamaño de los testículos y niveles de testosterona. Los animales con atrofia testicular no pudieron desarrollar los reflejos primarios de la cópula.
K88ab-GnRH disminuyó significativamente el largo y peso de los testículos y el peso de los epidídimos. P64K-GnRH sólo disminuyó (P 0,05) el peso de los testículos. GnRHm1-TT produjo una disminución significativa del largo, ancho y peso de los testículos y del peso de los epidídimos, vesículas seminales, glándulas bulbouretrales y próstatas, cuando se compararon todos con el grupo placebo (Tabla 1, Fig. 2). Entre los grupos K88ab-GnRH y P64K-GnRH no existieron diferencias significativas (P > 0,95). Por su parte, GnRHm1-TT disminuyó significativamente el largo de los testículos, comparado con K88ab-GnRH y P64K-GnRH, respectivamente y el ancho de los testículos, comparado con P64K-GnRH.
Los testículos de los cerdos inmunizados con K88ab-GnRH mostraron eliminación parcial (1/5) y completa (2/5) del parénquima funcional de los túbulos seminíferos (Fig. 3A). En los menos afectados se observaron escasos nemaspermos (3/5). Se encontraron casos de ligera fibrosis en los tabiques interlobulillares de los testículos, la que pudo provocar la disminución del diámetro de los túbulos seminíferos. En epidídimos se observó presencia mínima (2/5), moderada (1/5) y ausencia (2/5) de espermatozoides y fibrosis intersticial en las glándulas sexuales accesorias. En los cerdos inmunizados con P64K-GnRH se observó una disminución del diámetro de los túbulos seminíferos, en los que la mayoría de estos (3/5) contenían sólo células de Sertoli, presencia mínima (1/5), moderada (1/5) y ausencia (3/5) de espermatozoides en epidídimos y glándulas sexuales normales. Los testículos del grupo GnRHm1-TT mostraron una marcada disminución del diámetro de los túbulos seminíferos y atrofia, iniciada con degeneración vacuolar de las espermátidas, seguido de los espermatocitos, hasta alcanzar la completa eliminación del parénquima funcional de los túbulos seminíferos (4/5). Sólo en un cerdo se observaron ocasionalmente túbulos con espermatogonias. En epidídimos no observamos secreción espermática (4/5). En las vesículas seminales se observó fibrosis, en glándulas bulbouretrales engrosamiento de las trabéculas y fibrosis y en las próstatas abundante tejido fibrótico, con reducción de las glándulas tabulares secretoras. En el grupo placebo, los túbulos seminíferos de los 3 cerdos evaluados, mostraron varios tipos de células germinales y espermatozoides (Fig. 3B), espermatogénesis evidente y glándulas sexuales histológicamente normales. Ninguno de los grupos experimentales mostró alteraciones histológicas a nivel de las células de Leydig.
Este estudio estuvo dirigido a evaluar el posible uso de la fimbria K88ab y la proteína P64k, como portadoras de la GnRH, en el control de la fertilidad en animales domésticos y mascotas.
Una simple bacteria K88 fimbriada contiene varios cientos de miles de copias de FaeG en su superficie. Esta subunidad es empleada como portadora de péptidos derivados de la bacteria N. gonorrhoeae, del virus de la fiebre aftosa y virus de la inmunodeficiencia humana (Bakker, van Zijderveld, van der Veen, Oudega y de Graaf, 1990) y de péptidos derivados de la influenza y la somatostatina (Thiry, Clippe, Scarcez y Petre, 1989). La inserción de la GnRH en K88ab, puede haber provocado un cambio en la estructura conformacional de esta fimbria, lo que pudiera disminuir su reconocimiento por parte del anticuerpo monoclonal contra K88ab.
El alto peso molecular y la inmunogenicidad de P64k la convierten en una proteína portadora adecuada para inmunógenos débiles covalentemente acoplados (González et al., 2000) o en construcciones quimeras (González et al., 1997; Guillén et al., 1997). La inserción de GnRH en el plasmidio p64k-GnRH no afectó los niveles de expresión de la proteína P64k de N. meningitidis en E. coli.
Los posibles efectos farmacológicos provocados por estos candidatos vacunales fueron evaluados en cerdos prepúberes. En el análisis de los resultados aquí obtenidos debemos tener en cuenta que en ambas construcciones genéticas se insertó la secuencia que codifica para la GnRH de los mamíferos sin realizarle ninguna modificación en su composición aminoacídica. Van der Zee et al. (1995) y Mott y Reilly (1994) han fusionado la GnRH a las fimbrias P y a D. nodosus, respectivamente, pero hasta se conoce ninguno ha evaluado el efecto farmacológico de estas fimbrias híbridas en cerdos.
En los grupos P64k-GnRH y K88ab-GnRH, se obtuvo, respecto al placebo, una reducción del peso medio de los testículos de 40,4 y 52,7 %, (Fig. 2), respectivamente, debido principalmente a la disminución del diámetro de los túbulos seminíferos (Fig. 3A). Estos resultados fueron superiores a los obtenidos por Zeng et al. (2002). Por su parte, la reducción del peso de los testículos con K88ab-GnRH fue similar a la obtenida en cerdos con el esquema estándar de vacunación con Improvac® (Andersson et al., 2012). GnRHm1-TT, como era de esperar, tuvo una reducción mayor (73,9 %) del peso de los testículos (Fig. 2), comparable con los resultados obtenidos por Einarsson et al. (2011) y Yamsakul et al. (2017).
La neutralización de la acción de la GnRH endógena (Junco et al. 2008; Thompson, 2000) requiere de la unión directa de una suficiente cantidad de anticuerpos neutralizantes de la acción de esta hormona. En cerdos, no fue posible establecer un sistema tipo ELISA para determinar los títulos de anticuerpos anti GnRH; sin embargo, en otras especies animales se demostró la respuesta de anticuerpos anti GnRH con el mismo candidato vacunal GnRHm1-TT (Basulto et al., 2003; Junco et al., 2007). Al considerar la analogía de estos resultados nos permite inferir que, en cerdos, se pudiera producir un fenómeno similar.
El resultado obtenido con la GnRH en las proteínas portadoras K88ab y P64k fue inesperado, pues al contener mayor cantidad de epítopos T cooperadores era de esperar una mayor respuesta funcional que al usar un solo epítopo T cooperador de toxoide tetánico. Esto puede deberse a que estas proteínas también contienen epítopos B que pueden desviar la respuesta inmune o que se creen efectos estéricos que impidan una adecuada presentación antigénica. Este resultado también puede deberse a que la GnRH no contiene el cambio aminoacídico usado en la combinación GnRHm1-TT.
CONCLUSIONES
Los candidatos vacunales K88ab-GnRH y P64k-GnRH provocaron en cerdos prepúberes transformaciones estructurales y funcionales en testículos y epidídimos compatibles con la infertilidad. Sin embargo, el efecto biológico provocado por estos candidatos, que incluyeron la GnRH natural de los mamíferos, resultó menos homogéneo y efectivo, comparado con el péptido GnRHm1-TT, que incluye la variante modificada de esta hormona.
REFERENCIAS BIBLIOGRÁFICAS
ANDERSSON, K.; BRUNIUS, C.; ZAMARATSKAIA, G. y LUNDSTRÖM, K. (2012). Early Vaccination with Improvac®: Effects on Performance and Behaviour of Male Pigs. Animal 6(1), 87-95.
BAKKER, D.; VAN ZIJDERVELD, F. G.; VAN DER VEEN, S.; OUDEGA, B. y DE GRAAF, F. K. (1990). K88 Fimbriae as Carriers of Heterologous Antigenic Determinants. Microb Pathogen, 8 (1), 343-352.
BAKKER, D.; WILLEMSEN, P. T. J.; WILLEMS, R. H.; HUISMAN, T. T.; MOOI, F. R.; OUDEGA, B. et al. (1992). Identification of Minor Fimbrial Subunits Involves in Biosynthesis of K88 Fimbriae. J Bacteriol, 174(20), 6350-6358.
BASULTO, R.; MILANÉS, C.; ROJAS, A.; FUENTES, F.; IZQUIERDO, N.; BERTOT, J. et al. (2003). Efectos de la inmunocastración contra GnRH sobre la estructura y función testicular en perros adultos. Biotecnol Apl, 20(1), 20-24.
BRINGAS, R.; BASULTO, R.; REYES, O. y DE LA FUENTE, J. (1998). Vaccine for the Reversible Immunocastration of Mammals. Patent Cooperation Treaty (PCT) WO 98/27111.
CASTRO, M. D.; CAMPAL, ANA; LEÓN, L.; MIRANDA, A.; JUNCO, J.; ARTEAGA, N. et al. (2004). Inmunodominancia del epítope "a" de las fimbrias K88. Rev Prod Anim, 16(1), 59-66.
EINARSON, S.; BRUNIUS, C.; WALLGREN, M.; LUNDSTROM, K.; ANDERSON, K.; ZAMARATSKAIA, G. et al. (2011). Effects of Early Vaccination wiht Improvac® on the Development and Function of Reproductive Organs of Male Pigs. Anim Reprod Sci, 127(1-2), 50-55.
FIELDS, G. B. y NOBLE, R. L. (1990). Solid Phase Peptide Synthesis Utilizing 9-Fluorenylmethoxycarbonyl Amino Acids. Int J Pept Protein Res, 35(3), 161-214.
FUENTES, F.; JUNCO, J.; CALZADA, L.; BOVER, E.; SERRADELO, J. A.; HERNÁNDEZ, E.; PIMENTEL, E.; BASULTO, R.; REYES, O.; GARAY, H. y GUILLÉN, G. (2017). The Influence of Different Peptide Combinations to Increase the Immunogenicity of the Gonadotrophin Releasing Hormone Vaccine for Prostate Cancer Treatment. J Experiment Therap Oncol, 12 (1), 87-94.
GONZÁLEZ, G.; ÁLVAREZ, A.; CABALLERO, E.; VIÑA, L.; GUILLÉN, G. y SILVA, R. (2000). P64k Meningococcal Protein as Immunological Carrier for weak Immunogens. Scand J Immunol, 52(1), 113-116.
GONZÁLEZ, G.; PARDO, O.; SÁNCHEZ, B.; GARCÍA, J.; BEAUSOLEIL, I.; MARINELLO, P. et al. (1997). Induction of Immune Recognition of Self-Epidermal Growth Factor II: Characterization of the Antibody Response and the Use of a Fusion Protein. Vaccine Res, 6(4), 91-100.
GUILLÉN, G.; ÁLVAREZ, A.; SILVA, R.; MORERA, V.; GONZÁLEZ, S.; MUSACCHIO, A. et al. (1998). Expression in Escherichia coli of the lpdA Gene, Protein Sequence Analysis, and Immunological Characterisation of the P64K Protein from Neisseria meningitidis. Biotechnol Appl Biochem, 27(1), 189-196.
GUILLÉN, G.; DUARTE, C.; ÁLVAREZ, A.; et al. (1997). System for the Expression of Heterologous Antigens as Fusion Proteins. PCT Patent WO 97/26359.
HAN, X.; GU, L.; XIA, C.; FENG, J.; CAO, X.; DU, X. et al. (2015). Effect of Immunization against GnRH on Hypothalamic and Testicular Function in Rams. Theriogenol, 83(4), 642-649.
JANETT, F.; STUMP, R.; BURGER, D. y THUN, R. (2009). Suppression of Testicular Function and Sexual Behavior by Vaccination against GnRH (Equity) in the Adult Stallion. Anim Reprod Sci, 115(1-4), 88-102.
JUNCO, J.; BASULTO, R.; FUENTES, F.; BOVER, E.; CASTRO, M. D.; REYES, O. et al. (2008). Induction of Androgenic Ablation and Inhibition of Prostate Hormone-Sensitive Tumors by Immunization with the GnRHm1-TT Synthetic Peptide Adjuvated in VSSP. Biotecnol Apl, 25(2), 174-176.
JUNCO, J.; PESCHKE, P.; ZUNA, I.; EHEMANN, V.; FUENTES, F.; BOVER, E. et al. (2007). Immunotherapy of Prostate Cancer in a Murine Model using a Novel GnRH Based Vaccine Candidate. Vaccine, 25 (1), 8460-8468.
LEVY, J. K.; FRIARY, J. A.; MILLER, L. A.; TUCKER, S. J. y FAGERSTONE, K. A. (2011). Long-Term Fertility Control in Female Cats with GonaCon™, a GnRH Immunocontraceptive. Theriogenol, 76 (1), 1517-1525.
Mancini, M. C.; Menozzi, D. y Filippo, A. (2017). Immunocastration: Economic implications for the pork supply chain and consumer perception. An assessment of existing research. Livestock Sci., 203 (1), 10-20.
MILLAR, R. P.; PAWSON, A. J.; MORGAN, K.; RISSMAN, E. F. y LU, Z. L. (2008). Diversity of Actions of GnRHs Mediated by Ligand-Induced Selective Signalling. Front Neuroendocrin, 29(1), 17-35.
MOTT, M. R. Y REILLY, W. (1994). Double Immunogold Labelling Demonstrating Expression of Recombinant Genes for Production of an Anti-Fertility Vaccine. Micron, 25(6), 539-545.
SILVA, R.; SELMAN, M.; GUILLÉN, G.; HERRERA, L.; FERNÁNDEZ, J.; NOVOA, L. et al. (1992). Nucleotide Sequence Coding for an Outer Membrane Protein from Neisseria meningitides and use of Said Protein in Vaccine Preparations. European Patent Office, Publication number EP 0474313 A2.
STEVENS, J. D.; SOSA, J. M.; DEAVILA, D. M.; OATLEY, J. M.; BERTRAND, K. P.; GASKINS, C. T. et al. (2005). Luteinizing Hormone-Releasing Hormone Fusion Protein Vaccines Block Estrous Cycle Activity in Beef Heifers. J. Anim. Sci., 83(1), 152-159.
THIRY, G.; CLIPPE, A.; SCARCEZ, T. y PETRE, J. (1989). Cloning of DNA Sequences Encoding Foreing Peptides and their Expression in the K88 pili. Appl Environmental Microbiol, 55(4), 984-993.
THOMPSON, D. L. (2000). Immunization against GnRH in male species (comparative aspects). Anim. Reprod. Sci., 60 (1), 459-469.
VAN DER ZEE, A.; NOORDEGRAAF, C. V.; VAN DEN BOSCH, H.; GIELEN, J.; BERGMANS, H.; HOEKSTRA, W. et al. (1995). P-fimbriae of Escherichia coli as Carriers for Gonadotropin Releasing Hormone: Development of a Recombinant Contraceptive Vaccine. Vaccine, 13(1), 753-758.
YAMSAKUL, P.; PATCHANEE, P.; YANO, T.; BOONMA, T.; SOMKERT, C. y SATHANAWONGS, A. (2017). Gonadotropin-Releasing Hormone Immunization and Castration in Male Pigs: Effects on Growth, Hormonal Levels, Antibody Titer Response, Testicular Function, Back Fat, and Consumers' Sensory Perceptions. Korean J. Vet. Res., 57(1), 23-29.
ZENG, X. Y.; TURKSTRA, J. A.; MELOEN, R. H.; LIU, X. Y.; CHEN, F. Q.; SCHAAPER, W. M. et al. (2002). Active Immunization against Gonadotrophin-Releasing Hormone in Chinese Male Pigs: Effects of Dose on Antibody Titer, Hormone Levels and Sexual Development. Anim Reprod Sci, 70(1), 223-233.
Recibido: 10-3-2018
Aceptado: 16-3-2018
Roberto Basulto Baker, Centro de Ingeniería Genética y Biotecnología (CIGB), Camagüey, Cuba. roberto.basulto@cigb.edu.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}