










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
La industria porcina destaca entre las más promisorias para dar respuesta a las necesidades de alimentos que demanda la humanidad. Su producción mantuvo una tendencia de incremento hasta 2016, año en el que se produjo una disminución de 0,9 % de la tasa anual de crecimiento. Este viraje, entre otras, se debió a pérdidas económicas asociadas a patógenos respiratorios y gastrointestinales cuya presentación va en ascenso. También a estas causas se suman otras mermas, por lo general asociadas a un deficiente manejo. Precisarlas es esencial para adoptar medidas que reduzcan su impacto negativo (Rodríguez et al., 2020).
Las enteropatías (colibacilosis y salmonelosis) son una causa importante y frecuente de mortalidad en crías y precebas en los sistemas intensivos de crianza porcina (Barreto et al., 2020a), pese a lo cual persisten (Barreto, Rodríguez y Campal, 2020b). Un problema que se agrava con el uso de antibióticos, tanto de forma profiláctica como terapéutica, ya que los mismos afectan la microbiota intestinal, pobre en las crías y alterada luego del destete en las precebas (Rodríguez et al., 2020; Barreto y Rodríguez, 2021). Esta opción impone un riesgo a la salud de los consumidores y del entorno, y, además, potencia la selección y propagación de cepas antibiótico-resistentes (Espinosa et al., 2019). Solo los tratamientos que estabilicen una microbiota normal son efectivos al respecto. En este sentido destacan los basado en el empleo de prebióticos, probióticos y microorganismos eficientes (EM) (Barreto, Rodríguez y Campal, 2020b).
El receptor de hidrocarburos de arilo (AHR) es un factor de transcripción (dependiente de ligandos) que se expresa en las células inmunes, epiteliales, endoteliales, estromales y en tejidos. Una de sus funciones más relevantes y que se aviene al caso, es su papel como sensor central de diversos componentes de la dieta. Una interacción que condiciona la homeostasis intestinal y favorece la sinergia hospedero - microbiota, a la par que estimula una respuesta inmune local más efectiva (Stockinger et al., 2014; Mahringer et al., 2018; Avilla et al., 2020).
El eje AHR-IL-22 (eje aryl hydrocarbon - interleucina -22) en el intestino desempeña un papel importante en la defensa del huésped contra los patógenos microbianos. La señalización del AHR, a través de IL-22, inhibe la inflamación y la colitis en el tracto gastrointestinal de los ratones. La modulación de las vías de AHR es una estrategia terapéutica atractiva (Monteleone et al., 2011; Murray et al., 2016; Boule et al., 2018; Cervantes-Barragan y Colonna, 2018; Ehrlich et al., 2018). Varias evidencias indican que los indoles de la microbiota intestinal pueden alterar las respuestas a los factores estresantes agudos. Los derivados del indol pueden inducir IL-22 a través del AHR, lo que inicia un programa de reparación epitelial (Powell et al., 2020), que de conjunto con el empleo de prebióticos y probióticos, resultaría de gran utilidad para el tratamiento y control de las enfermedades gastrointestinales en cerdos.
El presente trabajo tiene como objetivo analizar las interacciones entre el AHR y la microbiota (favorecida por el uso de probióticos) en una mejoría de los parámetros de salud y productivos en cerdos (crías y precebas).
DESARROLLO
La producción porcina requiere especialistas veterinarios con conocimiento y las habilidades prácticas necesarias para el correcto desempeño de esa labor. Existen explotaciones que, en aras de un mayor rendimiento económico, no satisfacen los requerimientos mínimos básicos en cuanto a instalaciones, alimentación balanceada y manejo zootécnico veterinario (Barreto et al., 2020a) que asociados a las pérdidas por concepto de mortalidad tienen un impacto económico negativo considerable (Barreto, Rodríguez y Campal, 2020b; Rodríguez et al., 2020), por lo que disminuyen las posibilidades de cumplir los parámetros productivos establecidos para los cerdos al final del ciclo productivo.
Las enteropatías en los cerdos
Las enteropatías resultan muy frecuentes, y son un serio problema de salud en las unidades de cría y preceba porcinas ya que influyen significativamente en la mortalidad al ser la primera de sus causas (Rodríguez et al., 2020; Bárzaga, Alcolea y Rosell, 2021). En tales casos, múltiples enteropatógenos, a los que están expuestos estos animales desde el nacimiento, encuentran la brecha para afectar a las categorías más sensibles. Dos de los principales agentes bacterianos involucrados son Escherichia coli, en sus diversos patotipos, algunos de los cuales son zoonóticos y Salmonella. El patotipo enterotoxigénico de E. coli (ECET) ocasiona grandes pérdidas económicas, y E.coli productoras de toxina Shiga (ECST) provoca las diarreas postdestete, aunque ya se han aislado cepas híbridas ECET/ECST, tanto en casos de diarreas neonatales como pos destete. Las cepas del patotipo ECET provocan la mayoría de los casos y brotes de colibacilosis en los sistemas intensivos porcinos de todo el planeta (Barreto et al., 2020a; Guillén y Ríos, 2020). Razón por la que esta especie Escherichia figura entre los 40 patógenos que afectan a los cerdos con un síndrome gastrointestinal que tiene como dianas a crías lactantes y cerditos luego del destete (Barreto, Rodríguez y Campal, 2020b; Guillén y Ríos, 2020).
Por su parte, las salmonelosis septicémicas en cerdos son ocasionadas mayormente por Salmonella serovar Choleraesuis y, en menor grado, por Salmonella serovar Typhimurium; ambos zoonóticos. En los cuadros diarreicos por lo general se invierte su orden. También constituyen una importante causa de pérdidas en la producción porcina por concepto de tratamientos e incumplimientos con los compromisos asociados a ventas de carne y subproductos (Pastrana, Mogollón y Rincón, 2014; Barreto y Rodríguez, 2012; Barreto et al., 2020a).
Salmonella constituye una de las causas más frecuentes de gastroenteritis porcina en los países industrializados. Este agente llega al consumidor a través de alimentos de origen porcino, y con ello las zoonosis asociadas. Los productos de origen porcino por años han sido una importante causa de salmonelosis en Camagüey. Vale destacar que los cerdos pueden estar infectados por una cantidad inmensa de serovares, pero sólo son clínicamente afectados por S. Choleraesuis, S. Typhisuis y S. Typhimurium (Rodríguez et al., 2011; Barreto y Rodríguez, 2012; Pastrana, Mogollón y Rincón, 2014).
La tendencia por años para prevenir estos problemas han sido la suplementación de antibióticos en los piensos o la administración de los mismos al animal. Sin embargo, el empleo de antibióticos como profilácticos y promotores del crecimiento o como terapia no resuelve el problema y lo complica al alterar la microbiota intestinal, clave en el desarrollo y ulterior estado de salud del animal (Barreto, Rodríguez y Campal, 2020b). Por el contrario, el uso de prebióticos y probióticos, tanto en su forma comercial pura o como mezclas de los mismos como es el caso de los microorganismos eficientes (EM -del inglés efficient microorganims) o microorganismos autóctonos multipropósito (MAM) contribuyen al desarrollo de una microbiota en crías y al restablecimiento de la misma luego del estrés del destete (Barreto et al., 2015).
El AHR y las enteropatías porcinas
Un receptor es una molécula o una estructura polimérica que se localiza en la superficie o dentro de una célula que reconoce y se une a un compuesto endógeno. Los sitios de unión son estructuras tridimensionales que forman bolsas o ranuras en la superficie de la proteína que permiten interacciones específicas con compuestos conocidos como ligandos, que son moléculas de forma complementaria al sitio de unión a la proteína. Los receptores poseen un sistema efector (también denominadas vías de transducción de señales) (Rang et al., 2016; Riviere y Papich, 2018; Katzung y Trevor, 2019; Visovsky et al., 2019).
Un agonista es una sustancia que actúa activando o desbloqueando receptores celulares que provocan cambios y acciones en la función celular del cuerpo (respuesta) (Rang et al., 2016, Riviere y Papich, 2018; Katzung y Trevor, 2019; Visovsky et al., 2019), por lo que actúa como ligando de los receptores.
En los últimos 15 años se ha revelado que el AHR está involucrado en varios procesos fisiológicos como la homeostasis celular, la proliferación y diferenciación celular, la embriogénesis, la carcinogénesis, la inflamación y la inmunidad del huésped (Rademacher et al., 2018). Estudios recientes ponen de manifiesto las funciones moleculares del AHR en el sistema inmunológico durante el estado de equilibrio y durante la infección y la inflamación (Zhu et al., 2018). La capacidad del intestino para regenerarse mediante la proliferación y diferenciación de las células madre de las criptas sigue siendo vital para reparar el epitelio dañado y facilitar las respuestas al patógeno (Powell et al., 2020).
El AHR reside en el citoplasma y tras la unión del ligando, se traslada al núcleo donde heterodimeriza con el translocador nuclear del AHR (ARNT). El trímero AHR: ligando: ARNT se une a los elementos de respuesta de dioxinas (DRE) o de respuesta a xenobióticos (XRE) en las regiones reguladoras de los genes diana de AHR, que incluyen la monooxigenasa Cyp1a1 dependiente del citocromo P450, el represor AHR (AHRR) y la interleucina IL-22 (Fig. 1).
También se han informado vías de señalización de AHR no canónicas, ya sea a nivel genómico por asociación con otros factores de transcripción como el factor nuclear potenciador de las cadenas ligeras kappa de las células B activadas (NF-κB), o a nivel no genómico (por ejemplo, a través de la liberación de la tirosina quinasa c-SRC) (Gutiérrez-Vásquez et al., 2018; Lamas et al., 2018; Wang et al., 2020). Además de los xenobióticos (que incluyen el agonista prototípico de AHR 2,3,7,8-tetraclorodibenzo-p-dioxina (TCDD) se han identificado ligandos de AHR derivados de la dieta, muchos de los cuales son subproductos del metabolismo del aminoácido triptófano (Trp) (Hubbard et al., 2015; Wang et al., 2020).
El metabolismo del Trp por la microbiota intestinal genera ligandos agonistas de AHR que apoyan el desarrollo y mantenimiento de células linfoides innatas de tipo 3 intestinales. La señalización de AHR también es necesaria para el mantenimiento de linfocitos intra-epiteliales productores de IL-22. La IL-22 participa en la cicatrización de heridas de la mucosa y en la producción de péptidos anti microbianos por parte de las células epiteliales intestinales. El eje AHR-IL-22 en el intestino desempeña un papel importante en la defensa del huésped contra los patógenos microbianos, al mismo tiempo que garantiza la resistencia a la enfermedad (Cervantes-Barragan y Colonna, 2018; Ehrlich et al., 2018; Boule et al., 2018) (Fig. 2).

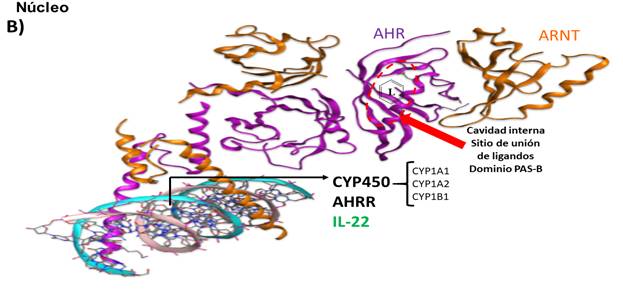
Fig. 1 Representación esquemática de la arquitectura estructural de la interacción de los monómeros AHR y ARNT para la formación de la región relacionada con la dimerización y unión de ligandos para el heterodímero funcional AHR: ARNT modelado por SwissModel (A), activación de genes y síntesis de proteínas tras la unión a los DRE o XRE (B). L: ligando para AHR.

Fig. 2 Representación esquemática de la mediación de los receptores y ligandos en la modulación de la respuesta inflamatoria al nivel intestinal (Fuente: Czerucka, 2021).
El papel clave de AHR en la microbiota intestinal e inmunidad del cerdo
El tracto gastrointestinal (TGI) ha desarrollado mecanismos de defensa frente a agentes ambientales adversos a los que está expuesto por vía oral (alérgenos, contaminantes, patógenos, etc.). Los microorganismos intestinales influyen en el desarrollo y en la función del sistema inmunológico. La rotura de este equilibrio con su anfitrión puede dar lugar a desregulación inmunológica y contribuir a la aparición de trastornos inflamatorios y autoinmunes crónicos (Álvarez et al., 2021). El AHR está involucrado en la regulación de las respuestas pro-inflamatoria y de tolerancia pues en las células inmunes intestinales el AHR estimula la producción de IL-22 (Bessede et al., 2014), e inhibe la inflamación inducida por la colitis experimental, sugiriendo esto que el AHR juega un papel clave en la resolución de la inflamación intestinal (Basson et al., 2021) (Fig. 2). El efecto supresor lo ejerce mediante la producción de la citocina antiinflamatoria IL-10 a través de c-SRC asociado a AHR (Zhu et al., 2018).
¿Por qué son importantes las IL-17 e IL-22?
Es importante la IL-22 para lograr la homeostasis a nivel intestinal. Se induce rápidamente en la mucosa del intestino en respuesta a IL-23, y a través de la activación del AHR para restaurar los daños debidos a patógenos o a dietas indebidas luego del destete. Se ha demostrado que la IL-22 juega un papel protector durante la infección con algunos patógenos, incluyendo Klebsiella pneumoniae, Citrobacter rodentium, Enterococcus resistente a la vancomicina y Plasmodium chabaudi. Uno de los mecanismos postulados por los que la IL-22 mejora la función de barrera de la mucosa es a través de la inducción de proteínas antimicrobianas a este nivel. Una función que explica en parte su papel en la contención de comensales al nicho intestinal. Por ejemplo, en ratones, el tratamiento con IL-22 disminuyó la abundancia de Escherichia coli de una manera dependiente de la dosis, lo que se correlacionó con la disminución de los niveles de endotoxina sérica (Basson et al., 2021).
La función principal de IL-17 (también inducida en respuesta a IL-23) es orquestar el reclutamiento de neutrófilos al sitio de la infección mediante la inducción de quimiocinas CXC (CXC porque tienen un aminoácido intermedio entre las dos primeras de cuatro cisteínas, como CXCL1 y CXCL8) (Soler, 2021), y mediante la mejora de granulopoyesis, que explica su función protectora durante la infección con una variedad de patógenos (Fig. 2).
Ligandos y metabolitos de triptófano de la dieta que interaccionan con el AHR
Los ligandos de AHR se clasifican en dos clases fisiológicas: "xenobióticos" y "endobióticos". Se designa el término xenobiótico para aquellos compuestos que se encuentran en un organismo y que no se producen dentro de ese organismo. Su presencia en el organismo es "ajena" o de una fuente ajena ("xeno"). Las fuentes comunes de ligandos xenobióticos incluyen por ejemplo las Dioxinas y sus congeneres. Por el contrario, reservamos el término ligando endobiótico para indicar cualquier ligando AHR que se produce fácilmente en un sistema biológico dado, incluso dentro del tracto gastrointestinal. Algunos ligandos endógenos ampliamente estudiados son 6-formilindolo [3,2- b ]carbazoles (FICZ), éster metílico del ácido 2- (1 ′ H -indol-3-carbonil) -tiazol-carboxílico (ITE), índigo, indirrubina, y bilirrubina (Avilla et al., 2020).
El agonista de AHR prototípico de alta afinidad, la 2,3,7,8-tetraclorodibenzo-p-dioxina xenobiótica (TCDD), muestra una afinidad 10 veces mayor por el AHR de ratón en comparación con el AHR humano. Por el contrario, los metabolitos del indol derivados de la dieta tienen una mejor afinidad por el AHR humano, como una posible consecuencia de la evolución (Hubbard et al., 2015; Murray et al., 2016; Lamas et al., 2018).
Entre los ligandos endógenos de AHR se encuentran una serie de productos metabólicos del triptófano como la indirrubina, quinurenina y 6-formilindolo [3,2-b] carbazol (FICZ). Además de los metabolitos del triptófano, el Lumichrome, un metabolito de la riboflavina, se identifica como ligando AHR endógenos en ratas ya en la década de 1980. Además, los productos de degradación del hemo, la bilirrubina y su precursor metabólico, la biliverdina, se reconocen como ligandos endógenos del AHR para la transformación AHR directamente activa y la transcripción de isoenzimas CYP. En el modelo de ratón con hiperhomocisteinemia, se ha encontrado que la lipoxina A4 (un metabolito del ácido araquidónico) aumenta notablemente la actividad de AHR y regula al alza la expresión de CD36. Estos metabolitos, junto con los metabolitos del triptófano, tienen un impacto importante en la homeostasis al regular la actividad de AHR (Kawajiri y Fujii-Kuriyama, 2017; Hattori et al., 2018; Larigot et al., 2018; Wang et al., 2020; Furue et al, 2021; Goya-Jorge et al., 2021; Kou, 2021).
El Trp es un aminoácido esencial que se obtiene a través de la dieta. La microbiota intestinal (Lactobacillus reuteri y Allobaculum) podrían catabolizar el triptófano en derivados de indol que son agonistas de AHR y en respuesta hay aumento en la producción de IL-22 que protege contra la inflamación en el colon (colitis), ya que los enteropatógenos bacterianos no son capaces de degradar a estos derivados (Marsland, 2016) (Fig. 2 y 3).
Agonistas de AHR derivados de Trp
Los ligandos de AHR derivados del Trp incluyen a compuestos como el FICZ (6-formilindolo [3,2-b] carbazol), que es un ligando agonista muy potente de AHR endógeno, pero que requiere el aminoácido triptófano (Trp) derivado de la dieta. Resulta de la conversión del Trp por fotooxidación dependiente de UV o estrés oxidativo mediado por H2O2 (Wincent et al., 2009). El siguiente ligando agonista endógeno de AHR identificado es la indirrubina (2- (2-oxo-1H-indol-3-iliden) -1H-indol-3-ona), pero su síntesis requiere del triptófano (Trp) derivado de la dieta. Es el resultado del metabolismo de Trp por la microbiota intestinal comensal y los hepatocitos del huésped. Se demostró que la indirrubina es más potente para activar el AHR humano que la 2,3,7,8-tetraclorodibenzo-p-dioxina (TCDD), el agonista del AHR prototípico, y es también un agonista más potente del AHR humano que el AHR murino (Adachi et al.,2001; Hubbard et al., 2015). El tercer compuesto de este grupo lo constituye el ITE (éster metílico del ácido 2- (1 'H-indol-3'-carbonil) -tiazol-4-carboxílico). Se ha planteado como hipótesis que se obtiene de la conversión gástrica de las glucobrasicinas, una familia de metabolitos altamente concentrados en las verduras crucíferas (como el brócoli y las coles de Bruselas), o de una reacción de condensación entre dos aminoácidos, el triptófano y la cisteína. ITE es un potente agonista de AHR tanto ¨in vitro¨ como ¨in vivo¨ y, a diferencia de la 2,3,7,8-tetraclorodibenzo-p-dioxina (TCDD), no induce toxicidad (Henry et al., 2010; Hubbard et al., 2015; Lamas et al., 2018). Por último, la L-quinurenina (Beta-Antraniloil-L-Alanina) fue el primer subproducto del metabolismo del Trp generado a través de la “vía de la quinurenina” enzimática. El modo de acción de la L-quinurenina en AHR aún no está claro. De hecho, podría ser un pro-ligando de AHR de baja afinidad que se convierte lentamente en compuestos de alta afinidad que actúan como agonistas de AHR a concentraciones subnanomolares (Bessede et al., 2014; Hubbard et al., 2015; Seok et al., 2018; Wang et al., 2020) (Fig. 3).

Fig. 3: Señalización de la formación de metabolitos derivados del triptófano. (Fuentes: www.elsitioporcino.com ; nutriNews, la revista de nutrición animal).
La solución conveniente para restaurar los daños debidos a patógenos o a dietas indebidas consiste en propiciar a través del alimento una microbiota adecuada durante la etapa de cría; luego, ayudar a restablecer la misma al destete, mediante formulaciones alimenticias líquidas que favorezcan el rápido restablecimiento de las microvellosidades, para lo cual son útiles los prebióticos y probióticos, los cuales constituyen opciones factibles a cualquier unidad especializada en crías y precebas porcinas (Barreto, Rodríguez y Campal, 2020b).
Un prebiótico es un sustrato que es selectivamente utilizado por microorganismos del hospedador y confiere beneficios para la salud. Este concepto sería aplicable a diferentes sustancias, incluyendo hidratos de carbono, polifenoles, ácidos grasos poliinsaturados, etc. (Álvarez et al., 2021). Por su parte, los probióticos pueden definirse como ¨microorganismos vivos que, cuando se administran en cantidades adecuadas, confieren un beneficio para la salud del hospedador¨. Los microorganismos comercializados como probióticos incluyen levaduras (Saccharomyces, Kluyveromyces) y bacterias de diferentes géneros (Lactobacillus, Streptococcus, Enterococcus, Pediococcus, Bifidobacterium, Propionibacterium, Bacillus), tienen beneficio demostrado para una condición de salud concreta (p. ej., diarrea aguda) (Álvarez et al., 2021).
Los probióticos deben presentar propiedades científicamente probadas tales como: 1) inhibición de los patógenos intestinales y extra intestinales; 2) inhibición de toxinas derivadas de patógenos y de los propios alimentos; 3) aumento de la capacidad de absorción de nutrientes, y 4) producción de sustancias con efectos bioactivos para el hospedero (Delgado, Barreto y Rodríguez, 2014). La alimentación adecuada con suplementación de nutrientes ricos en Trp o en sus derivados aquí mencionados, al actuar sobre el AHR tienen un papel estabilizador de la microbiota y del mejoramiento de la respuesta inmunitaria frente a las infecciones causadas por agentes patógenos en estas etapas de desarrollo y crecimiento, que, de conjunto con el empleo de prebióticos, prebióticos y MAM mejoraran la salud y los rendimientos productivos del rebaño.
CONCLUSIONES
Queda evidenciada la importancia los receptores AHR para restaurar los daños que pueden deberse a patógenos o a dietas indebidas en cerdos luego del destete por medio del aumento de IL-22, y propiciar el restablecimiento y estabilidad de la microbiota intestinal con el uso de los probióticos para promover respuestas favorables en los indicadores productivos y de salud de las crías y precebas porcinas

