










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Las temperaturas altas y la salinidad de los suelos son situaciones de estrés que afectan la productividad de los cultivos a nivel mundial. Para enfrentar estas problemáticas en la actualidad, se trabaja en la búsqueda de soluciones que permitan minimizar los daños que estos tipos de estrés provocan en las plantas, para lo cual se pudieran obtener y utilizar cultivares tolerantes e introducir tecnologías amigables con el medio ambiente.
Una de las tecnologías que se vienen desarrollando en los últimos años se basa en el uso de reguladores del crecimiento. Si bien en el mercado internacional existe una gran diversidad de estos compuestos, dentro de ellos, los brasinoesteroides son particulares, dada su función dual como estimuladores del crecimiento y protectores de las plantas contra el estrés. Los análogos de BR, sintetizados en Cuba, han demostrado ser productos más baratos, con utilidad práctica para incrementar los rendimientos agrícolas de 10 % a 25 % en diferentes especies, particularmente en condiciones no óptimas de producción.1
Existen diversos informes, en la literatura internacional, que demuestran la capacidad de los brasinoesteroides de inducir tolerancia a las plantas ante estrés de temperaturas altas y de salinidad;2),(3) sin embargo, no existía información acerca del papel de la homeostasis de brasinoesteroides en la tolerancia de las plantas al estrés por temperaturas altas. Por otra parte, la información que existe acerca de los mecanismos fisiológicos y bioquímicos asociados a la protección inducida por los brasinoesteroides en plantas sometidas a estrés salino no es suficiente y en el caso de la inducida por los análogos espirostánicos de brasinoesteroides es muy escasa. Además, de forma general, se desconoce si los brasinoesteroides y sus análogos utilizan los mismos mecanismos o no para inducir protección a las plantas ante condiciones de estrés.
Para dar respuesta a esta problemática y contribuir al conocimiento de la inducción de tolerancia en las plantas por los brasinoesteroides y sus análogos, se realizó el siguiente trabajo cuyo objetivo central fue, no solo demostrar el papel de la homeostasis de brasinoesteroides en la termotolerancia de plántulas de tomate y su vínculo con el estrés oxidativo inducido por la temperatura alta; sino además, determinar la tolerancia inducida por los brasinoesteroides y sus análogos; además de, profundizar en los mecanismos fisiológicos y bioquímicos asociados a la tolerancia a estrés salino inducida por los brasinoesteroides y sus análogos en plantas de arroz.
Métodos
Termotolerancia basal de diferentes genotipos de tomate y efecto de la aplicación exógena de brasinoesteroides y sus análogos
Con el objetivo de determinar el papel de la homeostasis de brasinoesteroides en la termotolerancia de plántulas de tomate (Solanum lycopersicum L.), se ejecutó un experimento donde se utilizaron diferentes genotipos caracterizados por cambios genéticos que originan niveles endógenos diferentes de la castasterona (CS). Así, se utilizaron plantas del mutante enano d x con el gen biosintético d bloqueado y que resulta deficiente de CS (concentraciones no detectables); del mutante enano abs1, que resultó de una mutación del dominio quinasa del receptor tBRI1 de los BR, el cual tiene alterada la sensibilidad a los BR y posee un alto nivel endógeno de la CS (18,56 × 10-7 mg kg-1 MFA). Además, se utilizaron plantas transgénicas (35SD), que se caracterizan por la acumulación de CS (5,75 × 10-7 mg kg-1 MFA) y la reversión del enanismo del mutante d x ya que sobreexpresan constitutivamente el gen d, bajo el control del promotor del virus del mosaico de la coliflor (CaMV35S). Las plantas del genotipo 35SD fueron generadas previamente mediante la transformación del mutante d x con la construcción pGB1421.4 Los genotipos d x y 35SD constituyen líneas isogénicas del cultivar de tomate Ailsa Craig (AC), un genotipo no alterado genéticamente en la biosíntesis y reconocimiento de los BR, por lo cual posee un nivel normal de la CS (2,17 × 10-7 g kg-1 MFA).
La caracterización de la termotolerancia basal se realizó colocando semillas desinfectadas de cada uno de los genotipos en potes plásticos estériles que contenían medio Murashige y Skoog suplementado con glucosa 1,0 % y solidificado con agar 0,8 % (m/v). Las plantas crecieron en condiciones controladas, a una temperatura de 25 ºC, humedad relativa de 50 % y un fotoperíodo de 16 h (10-4 mol m-2 s-1, luz fluorescente). A los 15 d de edad, las plántulas de todos los genotipos se sometieron a una temperatura de 45 oC por (4, 5, 6, 7 y 8) horas en la oscuridad en una cámara para ensayo ambiental. Al finalizar los choques térmicos, los potes se colocaron inmediatamente a 25 oC durante una semana (período de recuperación) en las condiciones de crecimiento mencionadas anteriormente. Se empleó la sobrevivencia como indicador de termotolerancia de las plántulas y se determinó al final del período de recuperación, estimándose a partir de la evaluación del número de plántulas que mantienen el ápice vivo o la emergencia de sus primeras hojas verdaderas.
Para evaluar el efecto de la 24-epibrasinólida (EBL) en la termotolerancia, se aplicaron diferentes concentraciones (0, 10-8, 10-7, 10-6 mol L-1) al medio MS cuando las semillas ya estaban germinadas y el choque por temperatura alta de 45 oC durante siete horas se impuso según se describió anteriormente. La termotolerancia se estimó a partir de la sobrevivencia como ya se ha descrito antes.
Para combinar el interés teórico con el práctico se ejecutaron experimentos con el genotipo Amalia, cultivo extendido en Cuba. Para ello, primeramente, las plántulas se sometieron a tratamientos de temperatura de 45 oC por 2 h, 3 h, 4 h y 5 h, siguiendo la metodología descrita anteriormente. Posteriormente, se evaluó la influencia de la EBL y dos análogos de la castasterona (A y B), sintetizados en Cuba, en la sobrevivencia y el crecimiento de plántulas del cultivar Amalia al choque de 45 oC durante 4 h. El análogo A corresponde a la formulación conocida como MH-5 que tiene como ingrediente activo al compuesto 25(R) 2(, 3(, 5( trihidroxi-espirostan-6-ona; mientras B corresponde a la formulación conocida como Biobras-6 y su ingrediente activo es la 25(R) 2(, 3( dihidroxi-5( espirostan-6-ona.
Las condiciones de crecimiento de las plántulas, la forma de aplicación, el rango de concentraciones de la 24-epibrasinólida y los dos análogos, la imposición del choque por temperatura alta, así como la estimación de la termotolerancia fueron similares a las ya descritas.
Estrés oxidativo en plántulas de diferentes genotipos de tomate tratadas con brasinoesteroides y sus análogos y sometidas a estrés por temperatura alta
Para evaluar la influencia que el choque por temperatura alta y la EBL ejercen en la estabilidad de las membranas, se determinó el eflujo de iones (EI) de las plántulas de los diferentes genotipos (AC, d x , abs1, 35SD), al finalizar el tratamiento de 45 oC durante 7 h, solamente con la concentración de EBL más efectiva. Los análisis de la peroxidación lipídica y la actividad de la enzima peroxidasa se realizaron antes de la imposición del tratamiento térmico, una vez finalizado este y al final de período de recuperación. La peroxidación lipídica se estimó mediante la determinación del contenido de malondialdehído (MDA) como se describió previamente.5
El ensayo de la actividad peroxidasa, POX (EC 1.11.1.7) se realizó a través del monitoreo del incremento de la absorbancia de la purpurogalina a 420 nm cada 10 s en un intervalo de 1 min.6 La concentración de proteínas solubles totales se determinó por el método de Bradford,7 utilizando la albúmina de suero bovino (1 g L-1) como estándar.
En el cultivo Amalia, también se evaluó el efecto de la EBL y los dos análogos en el contenido de MDA y en la actividad de la enzima POX.
Tolerancia al estrés salino de plántulas de arroz inducida por el tratamiento a las semillas con 24-epibrasinólida y dos análogos espirostánicos de brasinoesteroides
Para determinar la influencia que ejercía el tratamiento a las semillas con BR y sus análogos en el crecimiento de plántulas de arroz en medio salino, se ejecutaron varios experimentos en condiciones semicontroladas. En el primer experimento, se utilizaron semillas de los cultivares de arroz J-104 e INCA LP-7, las cuales se embebieron, durante 24 h, en agua, NaCl o NaCl suplementado con diferentes concentraciones (1,0; 0,5; 0,05; 0,005 mg L-1) de BB-6 o BB-16. Las semillas se germinaron en la oscuridad durante 48 h y luego, se transfirieron a potes que contenían las mismas soluciones de ensayo. Los potes se colocaron en un cuarto de luces a 25 oC y un fotoperíodo de 16 h durante 7 d. Al finalizar este período, se evaluaron las longitudes y las masas secas de parte aérea y raíces. Posteriormente, se ejecutaron dos experimentos, en el primero, se utilizaron semillas del cv. J-104 y en el segundo, semillas de Ginés, cultivares sensible y tolerante a la salinidad, respectivamente. Las semillas de ambos cultivares se trataron, durante 24 h, con diferentes concentraciones (0,0, 0,01; 0,1; 1,0 µmol L-1) de EBL o BB-16. Al finalizar la imbibición, las semillas se pusieron a germinar y 48 h después, las semillas germinadas se transfirieron a potes que contenían solución nutritiva Hoagland diluida (1:1) suplementada o no con NaCl 100 mmol L-1 y se colocaron en un cuarto de luces durante 11 d y 13 d, para J-104 y Ginés, respectivamente. Al finalizar el experimento, se realizaron las evaluaciones de crecimiento descritas anteriormente.
Finalmente, se ejecutaron otros dos experimentos donde se utilizaron semillas del cv. INCA LP-7, las cuales, en el primer experimento, se trataron con soluciones de BB-16 o BB-25 a concentraciones similares a las descritas en los experimentos anteriores. En el segundo experimento, se utilizó la mejor concentración de cada uno de los productos. Para la germinación de las semillas y el crecimiento de las plántulas se procedió de manera similar a los experimentos anteriores, aunque en estos casos los potes permanecieron en el cuarto de luces durante 14 d. En ambos experimentos, se evaluaron los indicadores de crecimiento y en el segundo experimento, además, se determinaron las concentraciones de pigmentos y de prolina en las hojas.
Identificación de mecanismos fisiológicos y bioquímicos asociados a la tolerancia al estrés salino inducida por la aspersión foliar de brasinoesteroides y sus análogos
Se ejecutó un primer experimento para seleccionar las concentraciones de EBL8 y de BB-169 más efectivas en la inducción de tolerancia al estrés salino en las plantas de arroz. Se ejecutó un segundo experimento para evaluar las respuestas fisiológicas y bioquímicas inducidas por la aspersión foliar de los brasinoesteroides en plantas de arroz sometidas a estrés salino. Para esto, se utilizaron semillas de arroz (Oryza sativa L.) del cultivar J-104 considerada como susceptible al estrés salino10 y se sumergieron en agua destilada durante 24 h. Posteriormente, se colocaron en placas Petri; en la oscuridad, para propiciar la germinación y a las 48 h, las semillas germinadas se trasplantaron a recipientes plásticos que contenían suelo Gleysol Nodular Ferruginoso Petroférrico11 y se introdujeron en cestas plásticas que contenían agua corriente. Las cestas se colocaron dentro de un umbráculo para evitar el efecto de las precipitaciones. En el momento de emergencia de la tercera hoja verdadera, se realizó la aspersión foliar con EBL (2 µmol L-1) o BB-16 (0,1 µmol L-1). Se asperjaron 240 plantas por tratamiento y se dejaron 240 como tratamiento control. En ese momento, las plantas se dividieron en dos grupos de 120 plantas, uno se colocó en cestas a las cuales se les adicionó solución nutritiva Hoagland diluida y el otro se colocó en cestas que contenían solución nutritiva Hoagland diluida suplementada con NaCl 100 mmolL-1. A los 7 d, las plantas que estaban sometidas a tratamiento salino, se colocaron en solución nutritiva Hoagland diluida, durante 14 d, para evaluar la recuperación. A los 7 d del tratamiento salino y al final de la recuperación se evaluaron indicadores del crecimiento (longitud y masa seca de parte aérea y raíz, área foliar), indicadores de las relaciones hídricas, solutos orgánicos (prolina, glicina betaína, azúcares solubles totales y reductores), cationes inorgánicos (Na+, K+, Ca2+ y Mg2+), contenido de clorofilas y carotenoides, daño oxidativo (contenido de MDA y proteínas oxidadas) y actividad de enzimas antioxidantes (POX, POXWC, CAT, SOD, APX, MDHAR, DHAR, GR).
Resultados
Termotolerancia basal de diferentes genotipos de tomate y efecto de la aplicación exógena de brasinoesteroides y sus análogos
Se evidenció, por primera vez, que la termotolerancia basal se modifica poco cuando el contenido de castasterona (CS) de las plantas de tomate se eleva entre la deficiencia (d x , mutante biosintético) hasta una cantidad intermedia de 5,75 × 10-7 g kg MFA (35SD, línea transgénica que sobre expresa el gen d). A partir de este nivel de hormona, un incremento adicional de hasta 18,56 × 10-7 g kg MFA produjo un aumento de la termotolerancia (abs1, mutación del dominio quinasa tBRI1). Se demostró que la inducción de termotolerancia por la EBL fue pronunciada tanto en tejidos con deficiencia como en los que poseen un nivel más elevado de CS; aunque el mutante d x resultó muy sensible a la EBL, respondiendo a la menor concentración estudiada (10-8 mol L-1). En general, con la aplicación de EBL 10-6 mol L-1 se obtuvieron los mayores niveles de inducción de termotolerancia de las plántulas de estos genotipos12 (Tabla 1).
Tabla 1 Porcentajes de sobrevivencia de plántulas de los diferentes genotipos de tomate tratadas con y sin la EBL, crecidas en el medio MS y sometidas a 45 °C durante 7 h (medias ± error estándar)*
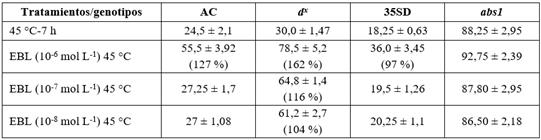
*Los números entre paréntesis corresponden a los porcentajes de incremento del indicador en los tratamientos con EBL con respecto a su correspondiente tratamiento control.
Resultados similares han sido expuestos por ) al demostrar la mayor inducción por la aspersión foliar de EBL a la concentración de 10-6 mol L-1.13
De igual manera, esta concentración resultó la más efectiva en estimular la sobrevivencia de plántulas de tomate cv. Amalia (Figura 1), incluso el incremento fue superior al informado con anterioridad14) y al observado en el genotipo Ailsa Craig (AC). En cuanto a los análogos A (MH-5) y B (BB-6), la mayor estimulación también se obtuvo a la concentración de 10-6 mol L-1, obteniéndose valores de sobrevivencia de 72 % y 58 % respectivamente; valores inferiores al obtenido con la aplicación de la EBL (85 %). Se debe destacar, que los resultados obtenidos confirman lo informado anteriormente, con relación a la termoprotección inducida por los análogos espirostánicos en este cultivo15 y sugiere la posibilidad futura del uso de estos productos para incrementar la termotolerancia de plántulas de tomate cv. Amalia.
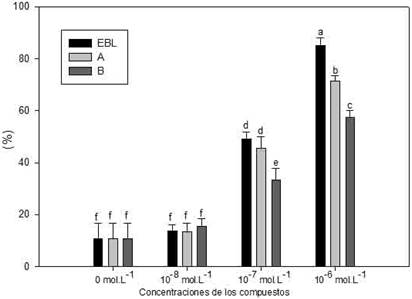
Fig. 1 Influencia de diferentes concentraciones de la 24-epibrasinólida (EBL) y los análogos A (MH-5) y B (BB-6) en la sobrevivencia de plántulas de tomate (cv. Amalia) sometidas al choque de 45 °C durante 4 h. Las barras representan los errores estándar de las medias y las letras las diferencias significativas entre tratamientos, según análisis de comparación de proporciones (p < 0,05).
Estrés oxidativo en plántulas de diferentes genotipos de tomate tratadas con brasinoesteroides y sus análogos y sometidas a estrés por temperatura alta
Se determinó que la temperatura incrementó el eflujo de iones de los diferentes genotipos de tomate (AC, d x , 35SD y abs1), inmediatamente después del tratamiento de choque térmico en la oscuridad; sin embargo, los genotipos con contenidos alterados de castasterona (d x , 35SD y abs1) no mostraron daño oxidativo. Esto sugiere que el efecto de la temperatura alta alteró primariamente la estabilidad de la membrana de las plántulas de estos genotipos sin que estas tuvieran daño oxidativo. Sin embargo, al final de la recuperación en la luz, se observó que, al aumentar el contenido de la CS, el daño oxidativo inducido por la temperatura alta se redujo. Así, las plántulas con ausencia de la CS (mutante d x ) se dañaron más y las que presentan el contenido más elevado (mutante abs1) se afectaron menos.
La EBL no propició la protección de las membranas de las plántulas de los genotipos AC y abs1 ante el daño oxidativo mediado por la temperatura alta; sin embargo, este compuesto ejerció un efecto protector en las plántulas de los genotipos d x y 35SD, siendo marcado en las primeras. De las actividades de las enzimas antioxidantes, las peroxidasas (POX) fueron las únicas que respondieron a la aplicación de la EBL, por lo que pudiera ser clave para las respuestas inducidas por los BR. La respuesta más pronunciada de la actividad POX se encontró cuando el tejido presenta deficiencia de CS, mientras que esta actividad no respondió al choque por temperatura alta en plántulas con un nivel elevado de CS. Al aplicarse la EBL, la actividad POX incrementó de manera pronunciada en el mutante d x ; sin embargo, en el mutante abs1, la EBL inicialmente incrementó la actividad POX, la cual fue disminuyendo durante la recuperación hasta igualarse al final del período.12
En el caso del cultivar Amalia, el comportamiento de los indicadores del metabolismo oxidativo (peroxidación lipídica y actividad de la enzima POX) ante la aplicación de los análogos A (MH-5) y B (BB-6) sugieren que ellos tuvieron efectos similares; aunque el análogo MH-5 proporcionó un mayor incremento del porcentaje de sobrevivencia; por lo que otros mecanismos, más que la reducción del daño oxidativo, podrían determinar la mayor capacidad de este análogo de incrementar la termotolerancia.
Tolerancia al estrés salino de plántulas de arroz inducida por el tratamiento a las semillas con 24-epibrasinólida y dos análogos espirostánicos de brasinoesteroides
Se demostró que el análogo BB-16 fue más efectivo que el BB-6 en estimular el crecimiento de plántulas de dos cultivares de arroz (J-104 e INCA LP-7) en medio salino, necesitándose una concentración inferior (0,01 µmol L-1) a las informadas para los brasinoesteroides naturales en la literatura internacional (de 1 µmol L-1 a 5 µmol L-1) por varios autores16),(17) en este cultivo.18
Posteriormente, se demostró que tanto la EBL como el BB-16 fueron capaces de revertir parcialmente la inhibición del crecimiento provocada por el NaCl en las plántulas de los cultivares de arroz J-104 y Ginés; siendo la EBL más efectiva que el BB-16 y se pudo comprobar que las plántulas del cultivar Ginés, tolerante a la salinidad, respondieron mejor a los tratamientos empleados.19 En otro estudio se comparó la efectividad de los análogos Biobras-16 y Biobras-25 en estimular el crecimiento de las plántulas del cv. INCA LP-7 en medio salino y se encontró que la modificación en las características estructurales que presenta el Biobras-25 favorecieron aún más el crecimiento de las plántulas de dicho cultivar, así como la concentración de pigmentos en las hojas.20
Identificación de mecanismos fisiológicos y bioquímicos asociados a la tolerancia al estrés salino inducida por la aspersión foliar de brasinoesteroides y sus análogos
Tomando como punto de partida, el establecimiento de las concentraciones más adecuadas a utilizar para la aspersión foliar con EBL8 o Biobras-169) con vistas a proteger a las plantas arroz de los efectos adversos que provoca el estrés salino; se ejecutaron experimentos que permitieron profundizar en las respuestas fisiológicas y bioquímicas asociadas a la protección inducida por cada uno de estos compuestos.
Se demostró que la concentración efectiva de BB-16 (0,1 µmol L-1) capaz de inducir protección fue 20 veces inferior a la utilizada para la EBL (2 µmol L-1) (Figura 2).
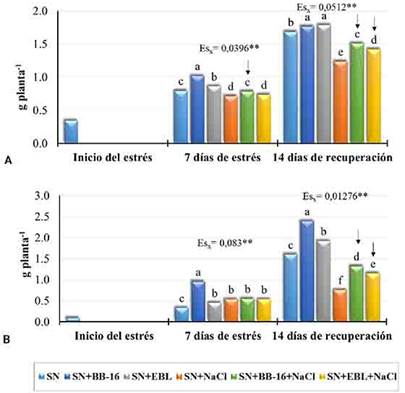
Fig. 2 Efecto de la aspersión foliar con BB-16 (0,1 µmol L-1) o EBL (2 µmol L-1) en la masa seca de plantas de arroz del cultivar J-104 sometidas a NaCl 100 mmol L-1 por 7 d. A. Parte aérea. B. Raíces. Medias con letras comunes no difieren significativamente entre sí según prueba de Rangos Múltiples de Tukey (p ≤ 0,05) n= 24.
En condiciones de salinidad, la estimulación del crecimiento inducida por la aspersión foliar con EBL o BB-16 está asociada a un mejor estado hídrico de las plantas, a un mejor balance de cationes como el K+, Mg2+y el Ca2+, a un incremento de la actividad de algunas enzimas antioxidantes (Figura 3) y a la disminución del daño oxidativo a lípidos y proteínas.21
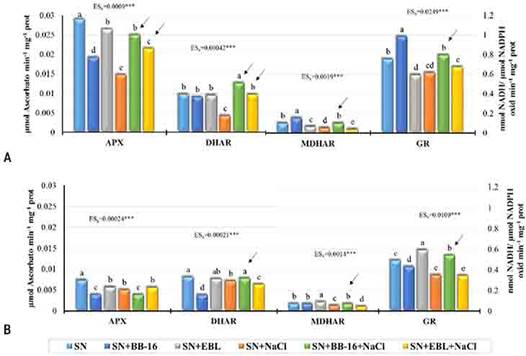
Fig. 3 Efecto de la aspersión foliar con BB-16 (0,1 µmol L-1) o EBL (2 µmol L-1) en la actividad específica de las enzimas del Ciclo del Ascorbato-Glutatión de plantas de arroz del cultivar J-104 sometidas a NaCl 100 mmol L-1 por 7 d. A. Final del período de estrés. B. Final del período de recuperación. APX: Ascorbato peroxidasas. DHAR: Deshidroascorbato reductasa. MDHAR: Monodeshidroascorbato reductasa. GR: Glutatión reductasa. Medias con letras comunes no difieren significativamente entre sí según prueba de Rangos Múltiples de Tukey (p ≤ 0,05) n= 9.
A pesar de que ambos brasinoesteroides protegen a las plantas jóvenes de arroz de los efectos adversos del estrés salino, se encontraron diferencias en la respuesta de las plantas a la aplicación de EBL o BB-16; lo cual se informa por primera vez. Por ejemplo, el BB-16, al finalizar la recuperación, incrementó significativamente la masa seca de la parte aérea y el área foliar con valores superiores al control sin sal. Sin embargo, con la aplicación de la EBL la longitud de las raíces fue superior al control sin sal. En las respuestas bioquímicas, se encontró que el BB-16 solo aumentó significativamente la prolina en hojas, mientras que la EBL aumentó este aminoácido en hojas y raíces. Este último, además, incrementó la glicina betaína en las raíces. Adicionalmente, en las enzimas antioxidantes se observó un efecto del BB-16, durante la recuperación, que no presentó la EBL.22 Estos resultados de la aplicación foliar del BB-16 durante la recuperación, sugieren un efecto a largo plazo de este análogo, lo que tendría influencia en la práctica productiva.
Las respuestas fisiológicas y bioquímicas obtenidas permitieron la elaboración de un modelo biológico (Figura 4) que sugiere, por primera vez, el modo de acción de la EBL en la inducción de tolerancia al estrés salino en plantas de arroz.22
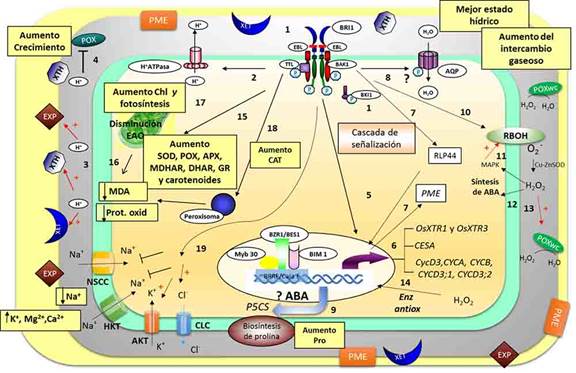
Fig. 4 Modelo de acción de la EBL en las células fotosintéticas de plantas jóvenes de arroz sometidas a estrés salino. 1) La EBL es reconocida por su receptor BRI1 y desencadena cambios conformacionales, heterodimerización con BAK1 y transfosforilación con lo que se activa totalmente el receptor. 2) La activación de BRI1 puede fosforilar a la H+ ATPasa tipo P con lo que se hiperpolariza la membrana y se acidifica el apoplasto. 3) El pH ácido activa enzimas modificadoras de la pared como las expansinas (EXP), las xiloglucano-endotransglicosilasas (XET) y las xiloglucano-transhidrolasas (XHT). 4) A su vez el pH ácido también inhibe las peroxidasas dependientes de pH (POX). 5) La activación del receptor por la EBL desencadena una cascada de señalización que termina en la asociación del factor de transcripción BZR1/BES1 con otros factores para el reconocimiento de promotores específicos en el ADN. 6) Este reconocimiento provoca la inducción de numerosos genes involucrados en el crecimiento y la expansión celular. 7) La interacción directa de RLP44 con BAK1 induce la expresión de genes de la pectinmetilesterasa (PME). 8) La activación de BRI1 a su vez regula la actividad de las acuaporinas (AQP), tal vez por fosforilación. 9) También se induce, por una vía dependiente de ABA, el gen P5CS, que codifica para la pirrolina-5-carboxilato sintasa, enzima clave en la biosíntesis de la prolina en condiciones de estrés. 10) Se incrementa la actividad de la NADPH oxidasa produciendo O2 - que es rápidamente dismutado por la Cu-Zn SOD a H2O2. 11) El peróxido activa las MAPK que a su vez inducen nuevas RBOH produciendo una amplificación de la señal. 12) Este H2O2 pudiera inducir la síntesis de ABA, produciendo una respuesta sistémica al estrés. 13) Las concentraciones de peróxido en el apoplasto pueden ser reguladas por peroxidasas asociadas a la pared celular (POXwc) que son inducibles por su sustrato. 14) El peróxido puede inducir la expresión de genes relacionados con la respuesta a estrés como enzimas antioxidantes. 15) El aumento de las principales enzimas antioxidantes como la SOD y las POX y las enzimas del ciclo del ascorbato-glutatión, unido al aumento de los carotenoides, pudieran provocar una disminución de las EAO en los cloroplastos. 16) Esto disminuyó el daño oxidativo en lípidos y proteínas. 17) Esta disminución de las EAO favorecería el aumento de las clorofilas y la fotosíntesis. 18) El aumento de la catalasa (CAT) puede contribuir a la eliminación del peróxido de hidrogeno y así también disminuir el daño oxidativo. 19) El mantenimiento de la integridad de la membrana plasmática por la disminución del daño oxidativo, unido a la regulación por los BR de la expresión y la actividad de transportadores de Na+ y K+ y las bombas de H+ pudiera conllevar a la disminución del Na+, así como el aumento del K+, Ca2+ y Mg2+. *Los recuadros en amarillo representan los resultados obtenidos en el trabajo
Conclusiones
La termotolerancia basal de genotipos de tomate estuvo asociada a concentraciones altas de castasterona endógena. Los brasinoesteroides provocan la activación de algunas enzimas antioxidantes e inhiben el estrés oxidativo inducido por el estrés de temperatura alta en tomate o el estrés salino en arroz para estimular la tolerancia de las plantas. Los análogos espirostánicos de brasinoesteroides, a determinadas concentraciones, protegen a las plantas de tomate y arroz ante condiciones de estrés por temperatura alta o salinidad, respectivamente; a pesar de que las respuestas fisiológicas y bioquímicas asociadas a dicha protección sean diferentes a la de los brasinoesteroides naturales.

