










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Las características de los componentes del complejo QRS están en dependencia de la forma en que se propaga la excitación por el tabique interventricular, las paredes libres y la base de cada ventrículo. En las especies del grupo A aparecen las tres ondas, en distintas derivaciones, que representan tres frentes definidos de activación ventricular (1,2,3).
En el grupo B, en el que se incluyen los bovinos, se obtienen dos ondas formando el complejo ventricular QRS, las cuales corresponden a la existencia de dos frentes de despolarización. La activación inicial es similar a la del grupo A, pero la propagación por las paredes libres, las bases de los ventrículos y el septum interventricular se produce en un solo golpe, como un estallido, en una dirección general ápico-basal y del ventrículo izquierdo al ventrículo derecho. Se origina así una sola onda, en lugar de la R y la S que se registran en los animales del grupo A (3).
Existen notables diferencias entre el ECG del hombre y el de los animales domésticos, que se deben, fundamentalmente, a la distinta situación del eje eléctrico cardíaco y de los puntos de derivación de la corriente, así como a la natural variación de las ondas, tanto en su dirección como en su amplitud, por cambios posturales y otras causas (4,5). Dentro del grupo A, el ECG del mono es similar en su morfología al del hombre, pero se diferencia en la duración de los segmentos e intervalos y en tener una frecuecia más elevada en la aparición de los complejos QRS. El objetivo de esta investigación fue determinar la morfología y la polaridad del complejo QRS del electrocardiograma del bovino Holstein, en diferentes puntos de la superficie corporal como referencia para el diagnóstico de alteraciones cardíacas.
MATERIALES Y MÉTODOS
Se utilizaron registros electrocardiográficos de 100 bovinos de la raza Holstein, clínicamente sanos y pertenecientes a diferentes grupos etarios de uno y de otro sexo: 10 terneras, 10 terneros, 20 novillas, 10 vacas lactantes, 25 sementales jóvenes y 25 adultos. La edad promedio de las terneras y de los terneros fue de 33 ±9 días, la de las novillas de 20 ± 3 meses, la de las vacas de 4 ± 2 años, la de los sementales jóvenes de 21 ± 4 meses y la de los adultos de 7 ± 3 años. Los terneros, las novillas y las vacas lactantes pertenecían al Distrito de Producción “Guayabal” de la Universidad Agraria de la Habana (UNAH); los sementales, al Centro Nacional de Inseminación Artificial “ROSAFÉ SIGNET, ubicado en Mayabeque.
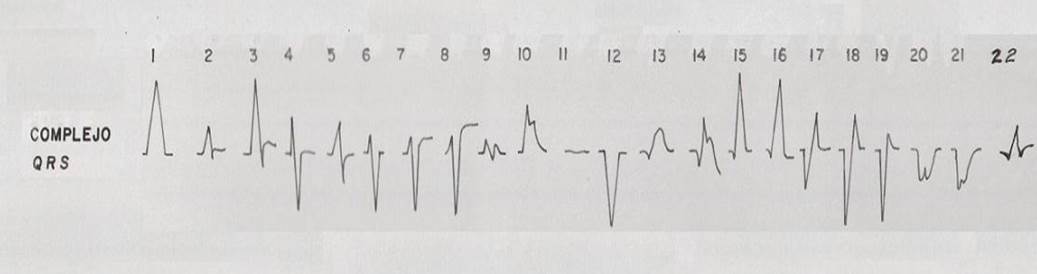
Los electrocardiogramas se efectuaron en 20 derivaciones, nueve bipolares y 11 monopolares y se exponen los datos de 18, por encontrarse dos de ellas (V1C yV2E) en la zona de transición y mostrar resultados muy similares aV2C (Tablas 1 y 2). En total, se observaron 1800 registros. El complejo QRS fue codificado de acuerdo a su morfología y se determinó la polaridad de cada una de las formas encontradas (Figura 1).
Se empleó un electrocardiógrafo portátil (HITACHI), calibrado con una señal de 1 mV / cm y una velocidad de corrida del papel de 25 mm/s. Durante el muestreo los animales se encontraban en estación, en un estado de correcto aplomo y en reposo sobre una manta de goma, como aislante de las corrientes parásitas.
Análisis estadístico
Se determinó la frecuencia de las formas del complejo QRS y de la polaridad en cada una de las 20 derivaciones anteriormente reportadas (6). Se realizó una comparación de proporciones para valorar si existían diferencias estadísticamente significativas entre la morfología de los registros obtenidos en derivaciones que originaran ondas con la misma polaridad.
Tabla 1 Puntos de colocación de los electrodos para obtener derivaciones bipolares en el bovino./ Electrode placement to obtain bipolar derivations in cattle.
| ELECTRODOS | ||
|---|---|---|
| Derivación | NEGATIVO (electrodo rojo o amarillo) | POSITIVO (amarillo o verde) |
| DI | Tercio medio del gran metacarpiano derecho (electrodo rojo). | Tercio medio del gran metacarpiano izquierdo (electrodo amarillo). |
| DII | Tercio medio del gran metacarpiano derecho (electrodo rojo). | Tercio medio del gran metatarsiano izquierdo (electrodo verde). |
| DIII | Tercio medio del gran metacarpiano izquierdo (electrodo amarillo). | Tercio medio del gran metatasiano izquierdo (electrodo verde). |
| IH | Sobre el quinto espacio intercostal derecho a la altura de la articulación escápulo humeral (electrodo rojo). | Por delante de la articulación escáspulo humeral izquierda (electrodo amarillo). |
| IIE | Sobre el quinto espacio intercostal derecho a la altura de la articulación escápulo humeral (electrodo rojo). | Tercio medio del gran metatarsiano izquierdo (electrodo verde). |
| IIH | Sobre el quinto espacio intercostal derecho a la altura de la articulación escápulo humeral (electrodo rojo). | Pliegue pregenual izquierdo (electrodo verde) |
| IIIE | Por delante de la articulación escápulo humeral izquierda (electrodo amarillo). | Tercio medio del gran metatarsiano izquierdo (electrodo verde). |
| IIIH | Por delante de la articulación escápulo humeral izquierda (electrodo amarillo). | Pliegue pregenual izquierdo (electrodo verde) |
| B-A | Sobre la séptima vértebra torácica. (electrodo rojo). | Apéndices xifoides del esternón, hacia el ápice del corazón (electrodo amarillo). |
Tabla 2 Puntos de colocación de los electrodos para obtener derivaciones monopolares pericordiales en bovino./ Electrode placement to obtain precordial monopolar derivations in cattle.
| ELECTRODOS | ||
|---|---|---|
| Derivación | REFERENCIA (VALOR CERO) | EXPLORADOR |
| V1H |
Amarillo en el metacarpo izquierdo. Rojo en el metacarpo derecho. Verde en el pliegue pregenual izquierdo. |
Por delante de la articulación escápulo humeral derecha. |
| V1C | Borde anterior de la extremidad derecha a nivel del olécranon. | |
| V2E |
Por delante del manubrio del esternón. Punto medio del segmento definido por las derivaciones V1C y V2C |
|
| V2C | Borde anterior de la extremidad izquierda a nivel del olécranon. | |
| V3C | Cuarto espacio intercostal izquierdo a nivel del olécranon, por detrás de la articulación del codo izquierdo. | |
| V4 | Entre V3C y V5. | |
| V5 | Apéndices xifoides del esternón. En el ápice del corazón. | |
| V6 | Cuarto espacio intercostal derecho a nivel del olécranon, por detrás de la articulación del codo derecho. | |
| V3H | Cuarto espacio intercostal izquierdo a la altura de la articulación escápulo humeral, por detrás de la articulación del hombro izquierdo. | |
| V3 | Cuarto espacio intercostal izquierdo, en el punto medio del segmento que une V3C y V3H. | |
| V10 | Sobre la séptima vértebra torácica. | |
RESULTADOS Y DISCUSIÓN
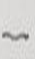
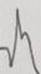
En la Tabla 3 se ilustran las 22 formas del complejo QRS que fueron encontradas, cada una de las cuales se asocia a un número para su codificación. Se puede observar que la frecuencia de la morfología y de la polaridad con que aparece cada una de las ondas del complejo QRS difiere entre las derivaciones bipolares estándares de extremidades, las de hombros y las monopolares pericordiales. En las bipolares estándares de extremidades se presenta la mayor diversidad en la polaridad, ya que aparecen +, +/-, +/-/+, - y -/+ y en la morfología, con 15 formas distintas en DI y 16 en DII e igual número en DIII. Debido a esto, estas derivaciones no son útiles como en el hombre (3,7,8) para diagnosticar alteraciones cardiovasculares, solo arritmias o trastornos en la conducción del impulso eléctrico (9-12). El complejo QRS, en los animales categorizados electrocardiográficamente en el grupo B, está constituido en todos los casos por la onda Q, como primer componente, y como segundo la R. De ahí que, tanto una onda como la otra, puede aparecer negativa o positiva, pues lo que determina su identificación es el orden con que aparecen y no su polaridad. Siempre la primera es la onda Q y la segunda la R y, si aparece una sola, se le denomina R (13). En los animales del grupo A y en los del B, el complejo QRS se modifica por diferentes factores que incluyen la disfunsión ventricular izquierda y derecha, la sobrecarga ventricular, el ejercicio y otros factores (14-17).
En las derivaciones de hombros, en IH las ondas Q resultaron siempre negativas y las R positivas; mientras, en IIE, IIH, IIIE y IIIH la onda Q adquiere signo positivo y la R negativo, lo que le proporciona gran estabilidad y confiabilidad para ser utilizadas con fines diagnósticos. Este hecho se debe a que se cumple la Ley de Kirchoff para los voltajes eléctricos en el circuito triangular formado por las tres derivaciones (18), en un caso IH, IIE, IIIE y en el otro IH, IIH, y IIIH. En este último triángulo, el electrodo positivo (color verde) se coloca en el pliegue pregenual izquierdo y no en el metatarso de ese lado. En la derivación Base-Ápice (B-A), reportada como muy estable con anterioridad (19), fue en la que con mayor frecuencia apareció la onda Q acompañando la onda R, en el 86 % de los casos con polaridad positiva y la R negativa apareció, además, sola en 14 %. El número máximo de formas diferentes del complejo QRS observadas en estas derivaciones bipolares fue bajo, a diferencia de las estándares de extremidades.
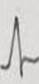
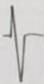
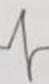
Tabla 3 Frecuencia de aparición de diferentes formas y polaridad del complejo QRS en derivaciones electrocardiográficas bipolares y monopolares, en 100 bovinos Holstein. /Frequency of appearance of different forms and polarity of the QRS complex in bipolar and monopolar electrocardiographic derivations in 100 Holstein cattle.
| Codificación. | 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 |
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Morfología |

|
|

|
|
|

|

|

|

|

|
|

|

|
|

|

|
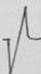
|
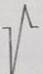
|

|
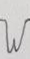
|

|

|
| Polaridad | + | +/- | +/- | +/- | +/- | +/- | +/- | +/- | +/-/+ | + | 0 | - | -/+ | -/+ | -/+ | -/+ | -/+ | -/+ | -/+ | - | - | -/+/- |
| DI | 37 | 1 | 1 | - | 4 | 2 | - | - | 3 | - | 11 | 18 | 2 | - | 4 | - | 7 | 2 | 6 | 1 | 1 | - |
| DII | 22 | - | 7 | - | 1 | 1 | - | - | 6 | 3 | 2 | 17 | 7 | 1 | 5 | 1 | 8 | - | 16 | 2 | 1 | - |
| DIII | 24 | 1 | 4 | - | 3 | 1 | - | - | 6 | 7 | 1 | 18 | 5 | 1 | 2 | - | 7 | - | 18 | 1 | 1 | - |
| IH | 49 | - | - | - | - | - | - | - | 1 | - | - | - | - | 45 | - | 4 | 1 | - | - | - | - | |
| IIE | - | - | 1 | 1 | 2 | 18 | - | - | - | - | 2 | 76 | - | - | - | - | - | - | - | - | - | - |
| IIH | - | - | 1 | 1 | 2 | 18 | - | - | - | - | 2 | 76 | - | - | - | - | - | - | - | - | - | - |
| IIIE | - | - | - | 3 | 1 | 20 | - | - | - | - | - | 76 | - | - | - | - | - | - | - | - | - | - |
| IIIH | - | - | - | 3 | 1 | 20 | - | - | - | - | - | 76 | - | - | - | - | - | - | - | - | - | - |
| Entre Base y Ápice (B-A) | - | - | - | - | - | 77 | 8 | 1 | - | - | - | 14 | - | - | - | - | - | - | - | - | - | - |
| V1H | 72 | - | - | - | - | - | - | - | - | - | - | - | - | - | 24 | 4 | - | |||||
| V2C | 75 | - | 2 | - | - | 1 | - | - | 2 | - | - | 3 | - | - | 15 | - | 1 | - | - | - | - | 1 |
| V3C | 3 | 1 | 2 | 1 | 6 | 22 | - | - | - | - | - | 64 | - | - | - | - | - | - | - | - | 1 | - |
| V4 | - | - | 1 | 1 | 2 | 41 | - | - | - | - | - | 55 | - | - | - | - | - | - | - | - | - | - |
| V5 | - | - | 1 | 2 | 3 | 49 | - | - | - | - | - | 45 | - | - | - | - | - | - | - | - | - | - |
| V6 | 2 | - | 19 | - | 12 | 32 | - | - | - | - | - | 35 | - | - | - | - | - | - | - | - | - | - |
| V3H | 68 | - | - | - | - | - | - | - | - | 1 | - | - | - | - | 25 | 5 | 1 | - | - | - | - | - |
| V3 | 45 | 1 | 1 | - | 1 | 1 | - | - | 1 | 3 | 8 | 12 | 5 | 1 | 12 | 2 | 5 | - | 1 | - | - | 1 |
| V10 | 49 | - | - | - | - | - | - | - | - | - | - | - | - | - | 42 | 9 | - | - | - | - | - | - |
En las derivaciones monopolares pericordiales V1H, V3H y V10, en el 100 % de los animales la onda Q fue negativa y la R positiva, mientras en V4 y V5 la Q resultó positiva y la R negativa. En las restantes derivaciones, V2C, V3C, V6 y V3 que pueden considerarse de transición por la posición que guardan respecto al dipolo originado por la distribución de cargas en los ventrículos, la polaridad de una y de otra onda fue muy variable. Mediante la derivación V3 se exploró sobre la superficie de los animales para determinar la línea isoeléctrica que separa los polos del vector eléctrico integral que origina el complejo QRS. Se encontró que en el punto medio del segmento que une las derivaciones V3C y V3H, ubicado en el cuarto espacio intercostal izquierdo,el complejo QRS fue nulo en el 8 % de los electrocardiogramas (Tabla 3). En esta última derivación, el 60 % del total de los complejos QRS estuvo constituido por una sola onda, por la R, 48 % positivas y 12 % negativas. Las ondas Q representaron el 32 %, con gran predominio de las negativas. La mayor diversidad en la morfología del complejo QRS se obtuvo, también, en V3 con 16 formas diferentes y la menor variación en V1H y V10 con tres formas distintas en cada una. Se detectaron diferencias estadísticamente significativas (p<0,05) entre la morfología obtenida en las derivaciones monopolares que tienen la misma polaridad, tanto en la base como en el ápice y entre la bipolar B-A que mide la diferencia de potencial entre estas dos regiones y las estándares de extremidades y las de hombro.
Como la derivación monopolar V3 se ubica en la región de transición del dipolo, de positivo a negativo o viceversa, la polaridad de Q y de R en esta región puede ser positiva, nula o negativa. Para ello, se tiene en cuenta que cuando el registro obtenido presenta dos componentes, se le denomina Q a la primera onda que aparece y R a la segunda, independientemente de sus respectivas polaridades y, si se registra una sola onda, corresponde a la R (13). Con este criterio, al valorar los datos de la Tabla 3, en V3 se registran 75 ondas R positivas en las formas codificadas como 1, 10, 13, 14, 15, 16, 17, 19 y 22, nulas 8 (forrma 11) y negativas 17, en las formas 2, 3, 5, 6, 9 y 12. La baja frecuencia de ceros y el predominio de ondas positivas (75 vs 17) indican que en la mayoría de los registros el electrodo explorador se colocó hacia la región positiva del dipolo, hacia la base del corazón. La anterior inferencia se confirma por el hecho de que el 100 % de las ondas R registradas en la derivación V3H, situada por encima de V3, son positivas, las cuales aparecen en las formas 1, 10, 15, 16, 17.
Uno de los segmentos más importante de la señal del ECG es el complejo QRS, que es originado por los potenciales que se generan cuando se despolarizan los ventrículos antes de su contracción (20). El centro eléctrico del corazón coincide con el centro del triángulo de Einthoven, de acuerdo con la teoría aceptada. Desde ese punto se puede fijar el inicio del eje eléctrico del QRS medio, que tiene en el hombre la dirección usual de arriba hacia abajo, de derecha a izquierda, de atrás hacia adelante (21,22). Marca el inicio de la contracción del ventrículo izquierdo, por lo que la detección de este evento tiene muchas aplicaciones clínicas (23). Tanto el incremento de su duración como su dispersión reflejan falta de homogeneidad del proceso y se asocian a un incremento de las arritmias ventriculares malignas (24-26). La prolongación de la conducción del impulso a través de los ventrículos produce prolongación del complejo QRS. Con frecuencia se produce esta alteración cuando uno o los dos ventrículos están hipertrofiados o dilatados, debido al trayecto más largo que debe recorrer el impulso (27-29). El ensanchamiento relevante del intervalo QRS se ha correlacionado con los niveles plasmáticos de ciertos fármacos (30) y, adicionalmente, su duración promedio en las derivaciones bipolares ha sido utilizada como indicador del rendimiento deportivo en equinos (31).
El tratamiento del corazón como un dipolo es muy útil para el diagnóstico de la ubicación aproximada de las áreas que puedan estar lesionadas, pues en tales circunstancias se afectan la dirección y el sentido del vector eléctrico integral que origina al complejo QRS, lo que se asocia también a modificaciones en la morfología de este complejo (32). El eje de QRS representa la suma vectorial de la despolarización ventricular y, por tanto, la mayor proporción se dirige al ventrículo izquierdo (33).
Las diferencias morfológicas del complejo QRS entre las epecies animales se deben al grado de penetración de las ramificaciones de Purkinje, lo que ha sido demostrado al estudiar su distribución en el miocardio de los mamíferos y de las aves (34-36), de lo que resulta que los animales pueden ser divididos o categorizados en dos grandes grupos: un grupo A, donde las fibras de Purkinje penetran hasta la mitad de la distancia del endocardio al epicardio y en el que se incluyen los primates, los caninos, los felinos y los roedores y un grupo B, en el que las fibras de Purkinje penetran completamente extendiéndose desde el endocardio hasta el epicardio en las paredes libres y en la base de los ventrículos. En este último grupo están comprendidos los equinos, los rumiantes, el cerdo y las aves (37).
En los animales categorizados electrocardiogáficamente como grupo A, el complejo QRS se compone de una onda principal R y dos depresiones situadas por delante y por detras de ella, que son la onda Q y la S, respectivamente. Cada uno de sus componentes refleja la parte de los ventrículos que se despolariza en el proceso de excitación ventricular; la onda Q corresponde a la actividad del Septum interventricular, la onda R a la propagación de la excitación por las paredes libres de los ventrículos y la onda S a la actividad de la base de estos (38,39), por lo que la secuencia de activación ventricular no acontece de la misma forma en todas las especies animales; ello está determinado por la diferente distribución del sistema de conducción cardíaca, o sea, el haz de His y sus ramas y las ramificaciones de Purkinje que penetran en las paredes ventriculares a diferentes profundidades (40,41).
En las investigaciones electrocardiográficas realizadas en búfalos (42) se señala que la morfología del complejo QRS se comporta muy variable, lo que puede manifestarse, también, dentro de una misma especie, ya que el patrón predominante de este complejo en la raza Jersey ha sido el del tipo QS, mientras que en la raza Red Dane ha sido el del tipo R o QR. Las observaciones de electrocardiogramas, correspondientes a 20 especies de vertebrados, revelaron que los procesos de cambio en las formas de onda del complejo QRS fueron los mismos que se han manifestado ontogénicamente en los embriones de pollos y en los fetos de rata (43). Se considera que la despolarización septal en el ganado bovino puede producir en el ECG registrado resultantes de muchas formas y de distintas amplitudes dentro de los límites fisiológicos. Esto está estrechamente relacionado con las propiedades conductivas de la masa corporal de los rumiantes atribuibles al volumen del rumen, lo que se acentúa más cuando existe repleción de este órgano o cuando el mismo es desplazado durante la gestación avanzada (44-46).
Durante el periodo de excitación, el miocardio se comporta como una serie de dipolos eléctricos conectados de una forma única denominada sincito. Esta actividad eléctrica del corazón es normalmente representada usando un vector variante en el tiempo, cuyas proyecciones pueden ser tomadas en la superficie del cuerpo (32). Varios factores pueden modificar la amplitud y el sentido en que se dirige este vector, por eso, se realiza el análisis vectorial del segmento ST y del complejo QRS con fines predictivos (47,48). En la infección chagásica aguda, 52,3 % de los exámenes electrocardiográficos presentaron alteraciones, las más frecuentes fueron alteraciones difusas de repolarización ventricular, bajo voltaje del QRS y desvío del eje cardíaco hacia la izquierda, lo que refleja señales de miocarditis difusa (49-51).
A partir de los resultados obtenidos se comprobó que la polaridad de las ondas, que constituyen el complejo QRS, es muy estable y la morfología varía muy poco cuando en las derivaciones bipolares los electrodos miden diferencias de potenciales entre los puntos de la base y del ápice del corazón, o el electrodo explorador se coloca en alguna de estas dos regiones en las derivaciones monopolares, lo que las convierte en una referencia importante para detectar alteraciones del sistema de conducción del impulso eléctrico en el corazón del bovino.

