










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
La resistencia a los antimicrobianos constituye un serio problema a nivel mundial. El uso indiscriminado de antibióticos en seres humanos, animales y la agricultura, unido al incremento de la población mundial, ejerce fuertes presiones sobre las comunidades de bacterias ambientales y contribuye a la difusión de sus genes de resistencia en los ambientes naturales.1,2 Esta situación ha facilitado la diseminación de bacterias gramnegativas multirresistentes, entre ellas bacterias productoras de BLEE.3,4) Los antibióticos son considerados contaminantes emergentes en los ecosistemas acuáticos y la mitad de los que consumen los seres humanos y los animales se excreta en forma activa a través de la orina y las heces, por lo que estos compuestos pueden llegar a acumularse en todas las matrices ambientales, principalmente las aguas superficiales, subterráneas, suelos y sedimentos.5
El agua constituye una importante ruta por la cual se introducen genes de resistencia bacterianos en los ecosistemas naturales, en donde las bacterias patógenas y no patógenas pueden servir como reservorio de genes de resistencia.6) Escherichia coli, miembro del grupo de los coliformes, es una especie bacteriana de la que se describen tanto cepas patógenas como no patógenas, por lo que es también utilizada como un indicador de la contaminación fecal en los ecosistemas acuáticos.7 Como patógeno, puede ser responsable de meningitis neonatal, infecciones urinarias recurrentes, septicemia y gastroenteritis, por lo que su presencia puede asociarse directamente al riesgo sanitario del empleo de aguas contaminadas con este microorganismo.8 En los últimos años, E. coli ha mostrado una elevada resistencia antibiótica tanto en cepas de origen clínico como ambiental, especialmente frente a los β-lactámicos.9)
Los β-lactámicos constituyen una familia numerosa y muy utilizada en la práctica clínica. Aunque la resistencia a esta familia está definida por distintos mecanismos, la producción de enzimas β-lactamasas es el principal. Las BLEE, constituyen una de las más difundidas entre las enterobacterias y particularmente en E. coli. La mayoría de las BLEE deriva de las β-lactamasas TEM-1, TEM-2 y SHV-1. Sin embargo, otra familia de BLEE, la CTX-M, ha iniciado una rápida expansión, aislándose en la actualidad con mayor frecuencia en diversas áreas epidemiológicas y en ambientes naturales.10
El aumento mundial de las enterobacterias resistentes a los antibióticos en los ambientes acuáticos hace que sea imperativo la vigilancia de la presencia y distribución de estas bacterias y de genes de resistencia procedentes de la clínica al medio ambiente. 11) En Cuba, se han realizado estudios de la resistencia de E. coli y sus mecanismos en la clínica;12 sin embargo, es poco lo que se sabe de la participación e influencia de los ecosistemas dulceacuícolas en la resistencia antibiótica y la diseminación de los mecanismos de resistencia de esta bacteria en esos ambientes. Por otra parte, los ecosistemas dulceacuícolas de La Habana están sometidos a un proceso antrópico acelerado, situación que ha provocado el deterioro de la calidad de sus aguas y la diseminación de bacterias resistentes a los antibióticos.13) El objetivo del presente trabajo fue evaluar la susceptibilidad antimicrobiana y la producción de enzimas BLEE de aislados de Escherichia coli procedentes de ecosistemas dulceacuícolas de La Habana.
Métodos
Aislamientos bacterianos
Se realizó un estudio descriptivo-longitudinal en el que se estudiaron un total de 43 aislados de E. coli procedentes de los ríos Almendares, Quibú y Luyanó de La Habana. Los aislamientos se realizaron entre septiembre de 2017 a enero de 2018 a partir de muestras de agua tomadas en diez estaciones de muestreo ubicadas en las zonas urbanas de los ríos capitalinos mencionados anteriormente. Los aislados evaluados forman parte la colección microbiana de la Facultad de Biología de La Universidad de La Habana.
Prueba de susceptibilidad a los antibióticos
Las pruebas de susceptibilidad a los antibióticos se llevaron a cabo mediante el método de difusión en disco descrito por Baüer-Kirby de acuerdo con las recomendaciones del Instituto de Estándares para el Laboratorio Clínico, 2020 (CLSI). 14) Se ensayaron los antibióticos ampicilina (10 µg), amoxicilina-ácido clavulánico (30/10 µg), ceftazidima (30 µg), cefotaxima (30 µg), aztreonam (30 µg), ceftriaxona (30 µg), cefepime (30 µg), piperacilina-tazobactam (100/10 µg), meropenem (10 µg), imipenem (10 µg) de la casa comercial Liofilchem, Italia. También se ensayaron contra antibióticos no β-lactámicos, incluyendo trimetropim-sulfametoxasol (1,25/23,75 µg), gentamicina (10 µg), amikacina (30 µg), cloranfenicol (30 µg), ciprofloxacina (5 µg), levofloxacina (5 µg), azitromicina (15 µg) y tetraciclina (30 µg) de la casa comercial Liofilchem, Italia. Se emplearon como cepas control Escherichia coli ATCC 25922, Staphylococcus aureus ATCC 25923 y Pseudomonas aeruginosa ATCC 27853.
Se calculó el índice de multirresistencia a los antibióticos (MAR) de acuerdo con los criterios establecidos por Krumperman.15 Se consideraron cepas multiresistentes aquellas que presentaron un índice MAR mayor que 0,5.15
Detección fenotípica de BLEE
Se realizó en todos los aislamientos que mostraron resistencia al menos a una de las cefalosporinas de tercera (cefotaxima (30 µg)/ceftazidima (30 µg)/ceftriaxona (30 µg)) y cuarta generaciones (cefepime (30 µg)) y el monobactámico aztreonam (30 µg), utilizando la prueba de discos combinados con inhibidor (cefotaxima (30 µg)-ácido clavulánico(10 µg)/ceftazidima (30 µg)-ácido clavulánico(10 µg)) de la casa comercial Liofilchem, Italia, de acuerdo con las recomendaciones del CLSI, 2020.15 Se incubó a 37 ± 1 ºC durante 18-24 h para realizar la lectura. La potenciación de la actividad de la cefalosporina en presencia del inhibidor indicó la producción de BLEE. Se midieron en milímetros los diámetros de las zonas de inhibición completa con un pie de rey. Las cepas control utilizadas fueron E. coli ATCC 25922 y Klebsiella pneumoniae ATCC 700603.
Además, se determinaron los patrones de resistencia de cada aislado ambiental mediante la lectura interpretada del antibiograma.16
Detección molecular de enzimas BLEE
Todos los aislados de E. coli caracterizados fenotípicamente como positivos para BLEE fueron analizados mediante reacción en cadena de la polimerasa (PCR). Se empleó el kit QIAGEN Multiplex PCR Master Mix. Para determinar la presencia de los tipos de genes bla CTX-M-1, bla CTX-M-2, bla CTX-M-9, bla OXA, bla TEM y bla SHV se utilizaron cebadores y condiciones descritas previamente17 (Tabla 1). El producto de amplificación se analizó por electroforesis en gel de agarosa 2 %, se tiñó con el fluorocromo SYBR (Invitrogen) (10 000X in DMSO, Lot 1942039) y se examinó a través de un transiluminador de luz ultravioleta (UV). La corrida electroforética se llevó a cabo a 100 V con solución de Tris Borato EDTA, pH= 8 (TBE). El patrón de peso molecular utilizado fue 100 pb Smart Ladder SF(Eurogentec).

Resultados
De los 43 aislados de E. coli estudiados, el 65 % fueron resistentes al menos a un antibiótico y el 35 % fueron sensibles a todos los antibióticos probados. Los β-lactámicos fueron la familia a la que presentaron mayor resistencia los aislados de E. coli y la de los fenicoles mayor susceptibilidad. Por otro lado, frente a los macrólidos y los aminoglucósidos mostraron el mismo porcentaje de resistencia (Fig. 1).

Fig. 1 Porcentajes de resistencia por familia de antibióticos de los aislamientos de E. coli aislados de los ríos de La Habana.
De los 15 aislados que fueron sensibles a todos los antibióticos, el 47 % de ellos presentaron una sensibilidad disminuída para uno de los 18 antibióticos analizados. Los aislados de E. coli exhibieron una mayor resistencia a la ampicilina (47 %), sin embargo, frente al resto de los β-lactámicos evaluados no presentaron valores de resistencia mayores del 9 %. Ninguno de los aislados mostró resistencia a imipenem, meropenem y piperacilina-tazobactam que también pertenecen a esta familia; aunque se encontraron dos aislados que presentaron susceptibilidad disminuída para piperacilina-tazobactam (Fig. 2).
Los aislados de E. coli evaluados mostraron elevada resistencia al antibiótico tetraciclina (35 %) seguida por la ciprofloxacina (25 %). Sin embargo, frente a los antibióticos cloranfenicol, amikacina y cefepime presentaron una baja resistencia con porcentajes por debajo del 4 %. Aunque los macrólidos no mostraron una elevada resistencia microbiana cabe destacar que se encontraron ocho aislados resistentes a la azitromicina, antibiótico de esta familia que se evaluó, provenientes de los tres ecosistemas evaluados (Fig. 2).

Fig. 2 Resistencia frente a los 18 antibióticos evaluados de los aislamientos de E. coli aislados de los ríos de La Habana.
El 37 % de los aislados de E. coli estudiados mostraron valores de índices MAR menores que 0,22. Sin embargo, el 16 % de los aislados tuvo un valor de MAR de 0,22 el cual se considera un valor intermedio, pero que a largo plazo puede transformarse en un valor de alto riesgo. El 9,3 % de los aislados mostraron resistencia a 10 de los 18 antibióticos probados, lo cual da lugar a un índice mayor que 0,5 que es considerado de alto riesgo (multirresistencia); y el 5 % mayor que 0,7 lo cual representa una resistencia a 13 de los 18 antibióticos usados lo que se considera un riesgo extremo (Fig. 3).
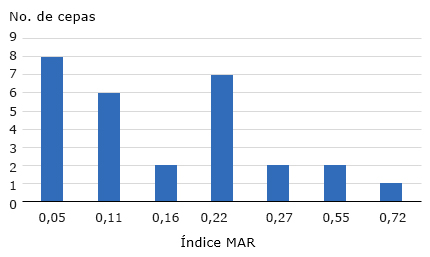
Fig. 3 Valores del índice de multirresistencia a los antibióticos (MAR) de los aislados de E. coli evaluados.
De acuerdo con la interpretación del antibiograma realizado, el fenotipo BLEE fue detectado en el 16,3 % de los 43 aislamientos ambientales de E. coli que fueron analizados teniendo en cuenta el patrón de resistencia frente a los antibióticos β-lactámicos evaluados, los cuales se describen en la tabla 2. Tres de los aislados provenían del río Almendares, uno del río Luyanó y tres del río Quibú. El número de antibióticos a los cuales los aislados mostraron resistencia varió de 8 a 13 para un total de cinco patrones de resistencia diferentes. De los patrones de resistencia, tres (60 %) se detectaron una única vez en la colección, mientras dos (40 %) se presentaron en al menos dos aislados (Tabla 2).
Todos los aislados de E.coli-BLEE resultaron resistentes a cefotaxima, aztreonam y ceftazidima 7 (100 %), mientras 5 (71,4 %), 7 (100 %) y 6 (85,7 %) fueron resistentes a ceftriaxona, ampicilina y amoxicilina/ácido clavulánico, respectivamente. Se observó co-resistencia para tetraciclina 5 (71,4 %), gentamicina 5 (71,4 %), trimetoprim/sulfametoxasol 6 (85,7 %), ciprofloxacina y levofloxacina 5 (71,4 %) y azitromicina 5 (71,4 %). Todos los aislados fueron completamente susceptibles al imipenem y meropenem (Tabla 2).
Tabla 2 Patrones de resistencia de los aislamientos ambientales de E. coli-BLEE aislados

AMP: ampicilina, AUG: amoxicilina, AZM: azitromicina, MRO: meropenem, CIP: ciprofloxacina, CRO: ceftriazona, IMI: imipenem, LEV: levofloxacina, CAZ: ceftazidima, FEP: cefepime, GN: gentamicina, TE tetraciclina, CTX: cefotaxima, TZP: piperacilina-tazobactam, AK: amikacina, ATM: aztreonam, STX: trimetropim-sulfametoxasol, C: cloranfenicol.
Entre los aislados de E. coli-BLEE se encontró que el gen bla CTX-M-1 es el más prevalente (4; 57,1 %) seguido de bla TEM (3; 42,8 %), no se obtuvieron amplificados de los genes bla CTX-M-2, bla CTX-M-9, bla OXA y bla SHV. El análisis de los productos de PCR no mostró coexistencia de enzimas BLEE entre los aislados de E. coli estudiados.
Discusión
En Cuba, se han realizado diversos estudios en los cuales se ha analizado el problema de la resistencia antibiótica en ambientes clínicos. 18) Sin embargo, el presente trabajo se realizó en ambientes dulceacuícolas y sus resultados ponen de manifiesto la presencia de bacterias resistentes a los antibióticos también en estos ambientes.
Aunque en Cuba los antibióticos requieren de prescripción médica por lo que la automedicación es menor con respecto a otros países del mundo, en la atención primaria los tratamientos son generalmente empíricos por parte del personal médico, lo que trae como consecuencia que hasta después de varios intentos fallidos de tratamiento es que se realiza un correcto antibiograma para un tratamiento específico.19) Por lo tanto, los antibióticos más usados en primera línea, los β-lactámicos, han sido los que mayor resistencia muestran en los últimos años,19 lo que también se ve reflejado en los resultados obtenidos en la presente investigación.
Más del 60 % de los aislados ambientales analizados presentaron resistencia a los antibióticos, lo que tiene una repercusión clínica y ambiental. La llegada de estas bacterias resistentes a los ecosistemas acuáticos está directamente relacionada con las aguas residuales que se han desechado directamente a los ríos de la ciudad sin ser previamente tratadas, las cuales constituyen fuentes de bacterias y de genes de resistencia a antibióticos que se liberan en el medio ambiente.7,13 Dichas aguas residuales provienen no solo de los hogares e industrias sino también de algunos hospitales, lo que contribuiría con la presencia de bacterias resistentes en los ríos. Se ha demostrado que una fuerte presión selectiva debido a la contaminación antropogénica, conlleva a la transferencia de genes que pueden propagar la resistencia de forma eficiente entre las poblaciones bacterianas.20
Del mismo modo, los genes de resistencia a antibióticos que estaban presentes de forma natural en los cromosomas de las bacterias del medio ambiente, ahora están presentes en plásmidos que se pueden transferir a los patógenos humanos.20 Se ha puesto de manifiesto que el contacto de las bacterias de la microbiota humana asociada con microorganismos ambientales en plantas de aguas residuales o en los ecosistemas naturales es una característica importante para entender la aparición de nuevos mecanismos de resistencia en patógenos humanos. Estudios en ecosistemas marinos indican que más del 90 % de las cepas bacterianas marinas son resistentes a más de un antibiótico, y 20 % son resistentes al menos a cinco.21
Una de las mayores preocupaciones clínicas de la resistencia antibiótica es la búsqueda de terapias alternativas a las infecciones causadas por microorganismos resistentes a los antibióticos. La creciente multirresistencia es un problema no solo en Cuba sino también en el mundo. En un estudio en Etiopía con muestras de agua hospitalarias así como del río en donde era liberada esta agua se encontraron cepas de E. coli resistentes a 11 de los 12 antibióticos probados, lo que demostró que la llegada de este tipo agua a los ríos promueve el aumento de la resistencia a los antibióticos22 e índices MAR mayores que 0,75. En México, en un estudio similar se encontró un índice MAR mayor que 0,5 en el 15 % de las cepas aisladas y se atribuyó a la acción antropogénica ejercida sobre el río.23
Por otra parte, el presente estudio muestra la presencia de aislados de E. coli-BLEE en ecosistemas dulceacuícolas de La Habana con altas tasas de resistencia a antibióticos utilizados como primera opción terapéutica en infecciones causadas por este microorganismo. De las β-lactamasas detectadas en las cepas ambientales evaluadas, las BLEE se consideran las más frecuentes en cepas aisladas del ambiente.24 A nivel mundial se han realizado estudios para evaluar la presencia de bacterias productoras de estas enzimas en aguas de ríos, lagos y aguas residuales.25
En la actualidad, las BLEE constituyen un problema terapéutico y epidemiológico, en el caso de las infecciones causadas por enterobacterias, pues las bacterias productoras de este tipo de β-lactamasas son resistentes a las penicilinas, las cefalosporinas y el aztreonam, y del 30 % al 60 % de ellas también a los β-lactámicos asociados a inhibidores de β-lactamasas; además, un porcentaje alto, por corresistencia, son también resistentes a las quinolonas, los aminoglucósidos, las tetraciclinas y el cotrimoxazol,26 comportamiento que presentaron los aislados BLEE positivos evaluados en la presente investigación.
Aunque los macrólidos no mostraron una elevada resistencia microbiana, cabe destacar la resistencia encontrada frente a la azitromicina. La concentración mínima inhibitoria (CMI) de este antibiótico no está correctamente definida para E. coli, es por ello que en los últimos años ha sido el centro de atención de muchos estudios.27 La resistencia a este antibiótico se debe generalmente a un gen presente en plásmidos móviles de las enterobacterias que crea una bomba de eflujo para la expulsión de este antibiótico. La azitromicina se usa para el tratamiento de E. coli desde hace pocos años, sin embargo, se ha estudiado su creciente resistencia en un corto período de tiempo y se ha detectado tanto en cepas patógenas como no patógenas.27
Los resultados de la presencia de diferentes tipos de genes bla TEM, bla CTX-M y bla SHV en aislados E. coli-BLEE es muy variable entre diferentes ciudades, países y regiones. En el presente estudio se encontró que la mayoría de los aislados presentaron el gen bla CTX-M apoyando el reconocimiento de que la enzima CTX-M es el tipo de BLEE más prevalente en el mundo. En estudios previos se había informado la enzima TEM como la más prevalente en los aislados clínicos de E. coli causantes de infecciones intrahospitalarias. Sin embargo, nuestro país al igual que en otras regiones del mundo como Europa del Este, América del Sur, Japón y la India, el gen bla CTX-M ha desplazado a otros genes que codifican para otras enzimas BLEE. 18,28)
La presencia de cepas productoras de estas enzimas en los ríos evaluados podría estar dado por el tipo de fuentes contaminantes que impactan estos ecosistemas. En un estudio realizado previamente en el 201129) en el río Almendares sobre la presencia de genes de resistencia (ARG), se encontraron correlaciones significativas (p< 0,05) en los sedimentos con respecto a los ARG, especialmente para las tetraciclinas y β-lactámicos, así como para ampicilina en la columna de agua en el río.
En los ambientes naturales, la detección oportuna de una cepa productora de BLEE es importante, pues si una comunidad de bacterias ambientales resistentes a los antibióticos incrementa su frecuencia es probable que permanezcan allí por mucho tiempo. Estos resultados subrayan la necesidad de estrategias para reducir el desarrollo de resistencia y así prolongar la vida útil de nuestros antibióticos disponibles.
Conclusiones
La presencia de aislados multirresistentes de E. coli productores de BLEE, con una prevalencia del gen bla CTX-M en ecosistemas dulceacuícolas de La Habana pone de manifiesto el riesgo potencial para la salud que representa el uso de las aguas de estos ríos e indican la necesidad de implementar estrategias de control para prevenir la diseminación de estos aislados.

