










 Servicios personalizados
Servicios personalizados Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Los sistemas silvopastoriles (SSP), con énfasis en los intensivos, constituyen una solución innovadora importante para incrementar la producción ganadera, principalmente la bovina y la ovino-caprina (tanto de leche como de carne, y de otros bienes asociados), y enfrentar el cambio climático en los países de América Latina y el Caribe, y de otras áreas tropicales y subtropicales del mundo (Murgueitio et al., 2015).
Lo anteriormente señalado se basa en las ventajas de esos sistemas (una mayor eficiencia de los procesos biológicos que ocurren en ellos, debido a que se maximiza la transformación de la energía solar en biomasa, se favorece la fertilidad del suelo con la fijación biológica del nitrógeno atmosférico, hay una mayor solubilización del fósforo y acumulación de materia orgánica por las deyecciones -orina y bostas- de los animales) y en los principios agroecológicos que se aplican para manejarlos, tales como: el empleo del pastoreo racional-rotacional, con una alta carga instantánea de animales resistentes y adaptados a determinadas condiciones climáticas, en un ambiente donde exista una adecuada conservación de la biodiversidad, los pastos y forrajes posean el tiempo de reposo necesario para que alcancen la calidad nutricional requerida, y se maneje correctamente el recurso agua, con vista a garantizar el bienestar de los animales. Ello, en su conjunto, hace que se produzca una mayor captura de CO2 en el suelo y menores emisiones de CH4; y que se incremente la resiliencia al cambio climático, con un uso mínimo de agroquímicos, hormonas y antibióticos respecto a los sistemas convencionales.
Otro aspecto a destacar en los SSP con arreglos agroforestales, según los criterios de Murgueitio-Restrepo et al. (2016) y Sisa-Benavides (2017), es la inclusión de plantas arvenses dentro del componente herbáceo (además de las especies pratenses de gramíneas y leguminosas volubles, fundamentalmente), las que interactúan con el estrato arbóreo (árboles y arbustos forrajeros, frutales, maderables, entre otros) y con los animales. Por tanto, en el ámbito ganadero debe reanalizarse el criterio de que estas sean consideradas como indicadores de un manejo inadecuado del pastizal, según señalan Milera et al. (2014).
Además, las arvenses desempeñan un papel importante en la red trófica de los agroecosistemas complejos, ya que interaccionan directa o indirectamente con otros de sus componentes y ofrecen un amplio espectro de funciones ecológicas y agronómicas, como la polinización y la regulación de plagas, al servir de refugio a los agentes naturales y biológicos de control (Caballero-López et al., 2012). Ello potencia el manejo de la agrobiodiversidad, y su función se explica mediante la regulación biótica a través de los mecanismos bottom-up y top-down (Altieri y Nicholls, 2010; Ratnadass et al., 2012), específicamente este último, ya que al disponer de un reservorio de depredadores y parasitoides se lograría el control de la plaga por sus enemigos naturales.
En tal sentido, el objetivo del estudio fue determinar las especies de plantas arvenses y de la entomofauna beneficiosa presentes en los sistemas silvopastoriles (SSP) evaluados en el occidente de Cuba, así como su interrelación ecológica en esos agroecosistemas ganaderos.
Materiales y Métodos
Las áreas experimentales se correspondieron con SSP (cuartones) compuestos por asociaciones de las cuatro variedades comerciales de la leguminosa arbórea Leucaena leucocephala (Lam.) de Wit -aprobadas en Cuba, según MINAG (2017) y diferentes gramíneas pratenses, las cuales tenían las particularidades que se relacionan en la tabla 1.
Tabla 1 Caracterización de los sistemas de producción ganadera.
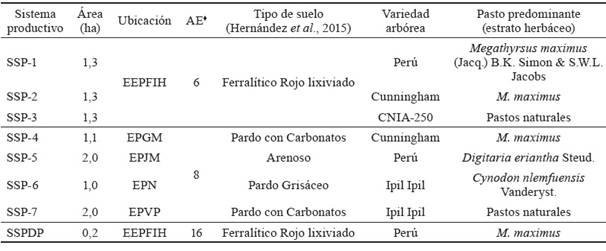
SSP: sistema silvopastoril, SSPDP: SSP doble propósito (ceba bovina-producción de semilla), EEPFIH: Estación Experimental de Pastos y Forrajes Indio Hatuey, EPGM: Empresa Pecuaria Genética de Matanzas, EPJM: Empresa Pecuaria José Martí (provincia
Matanzas), EPN: Empresa Pecuaria Nazareno (provincia Mayabeque), EPVP: Empresa Pecuaria Valle del Perú (provincia Mayabeque), AE: años de explotación; ♦: indica a partir del segundo año de establecimiento hasta el momento de la evaluación.
El período experimental fue de un año, teniendo en cuenta los meses de mayor presencia de los insectos en estos sistemas (marzo, mayo, octubre y noviembre), determinados con anterioridad por Alonso (2009).
El muestreo de las especies arvenses se realizó al inicio y al final del período de evaluación, en cinco puntos de cada cuartón (método del sobre), utilizando un marco de 1 m2. La recolección de los insectos beneficiosos se hizo cada 15 días, antes de que entraran los animales a pastorear en todas las áreas, con el empleo de dos métodos -basado en la propuesta de Nielsen (2003)- para garantizar la mayor captura posible de individuos presentes: la red entomológica (tanto en el estrato arbóreo como en el herbáceo), con la que se efectuaron 100 pases en cinco puntos de los campos evaluados, lo que equivale a 25 m2 según Faz (1990); y el nailon transparente en el caso de las plantas arbóreas, con el cual se tomó una muestra de los órganos presentes en la zona de ramoneo (2 m de altura, aproximadamente) en los cuatro puntos cardinales, según la fenología del cultivo de la leucaena en el momento de muestreo.
Los subgrupos funcionales de los insectos capturados se conformaron según el criterio de Ruiz y Castro (2005) y las observaciones de su hábito principal de alimentación, realizadas en campo.
Entre los principales indicadores del manejo de los sistemas productivos que se tuvieron en consideración en el comportamiento de las arvenses y los insectos beneficiosos se encuentran: la labor de defoliación (poda o corte) en el período de escasez de alimento (forraje); el tiempo de reposo de los cuartones después del pastoreo (28-45 días en el período lluvioso y 49-66 en el poco lluvioso); y la no aplicación de fertilización inorgánica u orgánica, riego, plaguicidas químicos o biológicos. Adicionalmente, se determinó el porcentaje de área cubierta por los pastos herbáceos predominantes como parte de la composición florística, mediante el método de los pasos EEPFIH (1980).
La identificación de las arvenses y de los insectos se realizó en los laboratorios de protección de plantas de la EEPFIH y en el de taxonomía de insectos del Centro de Investigaciones Agropecuarias (CIAP) de la Universidad Central Marta Abreu de Las Villas (UCLV) -Villa Clara, Cuba-, con el empleo de claves taxonómicas (Borror y White, 1970; Sánchez y Uranga, 1993; Peck, 2005), la revisión y comparación con las colecciones de insectos de la UCLV, y la contribución de otros especialistas de diferentes instituciones del país.
Resultados y Discusión
Las arvenses, después de los muestreos en los ocho sistemas ganaderos en estudio, no aparecieron en dos (SSP-1 y SSP-2); mientras en los seis restantes se encontraron e identificaron 34 especies, las cuales se listan en la tabla 2.

De estas arvenses sobresalieron M. pudica y S. rhombifolia, presentes en cinco de las áreas; D. cinerea, en cuatro; y S. geniculata, A. aspera y P. guajava, en tres, las cuales son consideradas especies típicas de los pastizales en Cuba según los informes del sector ganadero.
Respecto a los insectos beneficiosos capturados en las áreas muestreadas, se identificaron 79 especies (44 depredadores, 30 parasitoides y 5 polinizadores), pertenecientes a los ordenes Coleoptera, Hymenoptera, Diptera, Hemiptera, Orthoptera, Mantodea, Neuroptera, Dermaptera, Odonata y Thysanoptera. De estas, 27 pueden encontrar refugio o alimentarse de cinco de las arvenses encontradas y regular nueve de los fitófagos plagas también hallados en estos sistemas silvopastoriles, de acuerdo con el criterio de varios autores acerca de su interacción en el contexto agropecuario cubano (tabla 3).
Tabla 3 Relación ecológica entre arvenses, insectos beneficiosos y fitófagos plagas asociados a los SSP evaluados.

■depredadores, ( parasitoides; °polinizadores, Δ plagas reguladas por C. cacti, C. maculata y C. sanguinea limbifer.
Fuente: Bruner et al. (1975), Fernández T. et al. (2001), Valenciaga (2003), Veitía (2004), Martínez et al. (2007), Milán et al.(2008), Vázquez et al. (2008), Alonso (2009).
No obstante, en otros cultivos en diferentes países se informa acerca de la relación entre los insectos beneficiosos y las plantas arvenses que se encontraron en este estudio, lo cual se describe a continuación:
En el caso de I. trifida, en zonas aledañas a los cultivos de arroz, maíz y algodón en Colombia, se albergaron insectos de las familias Chalcididae (Hymenoptera), por ejemplo: Conura sp. y Brachymeria sp.; Anthocoridae (Hemiptera) (Orius sp.), Dolichopodidae (Diptera) (Condylostylus sp.), Syrphidae (Diptera), Coccinellidae (Coleoptera) (C. maculata y Scymnus sp.) y Reduviidae (Hemiptera) (Zelus sp.); además, de conjunto con S. acuta, proporcionaron polen y néctar a insectos de la familia Vespidae (Hymenoptera); mientras C. diffusa fue hospedante de insectos de la familia Syrphidae (Bedoya et al., 2018). Por otra parte B. pilosa, considerada en Perú como maleza en el cultivo del lúcumo [Pouteria lucuma (Ruiz & Pav.) Kuntze], sirvió de alojamiento a insectos de la familia Coccinellidae (Castillo et al., 2015).
La relación plantas arvenses-insectos beneficiosos en este estudio pudo estar influenciada por el manejo dado a los sistemas productivos, lo cual es una respuesta esencialmente al cubrimiento del área por las especies pratenses y forrajeras (tabla 4).
Tabla 4 Cantidad de taxones de arvenses e insectos beneficiosos y porcentaje de área cubierta por los pastos herbáceos predominantes en los SSP.

% AC PHP: porcentaje de área cubierta por el pasto herbáceo predominante, F + B: fitófagos + beneficiosos, De + Pa: depredadores + parasitoides, ♦ Indica al final del experimento. SSP: sistema silvopastoril, SSPDP: SSP doble propósito (ceba bovina-producción de semilla).
En los cuartones 6 y 7, dos de los que tuvieron menor porcentaje de AC PHP, se encontró el mayor número de especies arvenses; lo cual tuvo relación, además, con que en el primer cuartón no se realizó correctamente la labor de poda según lo establecido, y en el segundo no se podó y los animales no rotaron adecuadamente por problemas con la cerca perimetral. Se destaca, a su vez, un incremento de la presencia de beneficiosos, ya que las arvenses les sirven de refugio o para la alimentación (polen o néctar) de los adultos, como plantean Altieri y Nicholls (2007).
En el cuartón 3 el porcentaje de AC PHP fue aún menor, debido a las inundaciones prolongadas antes y durante el período experimental, lo que ocasionó la aparición de otras plantas, pero en este caso pratenses (Indigofera oxycarpa Desv. (= I. mucronata Spreng. ex DC.), de la familia Fabaceae y no arvenses.
La presencia del mayor número de especies de beneficiosos en SSP-1, SSPDP y SSP-4, donde hubo un adecuado porcentaje de AC PHP, estuvo relacionada con la condición de macollosa de la guinea, lo cual genera un mayor número de hábitats donde se refugian los insectos depredadores, parasitoides y polinizadores, entre otros beneficiosos (Alonso, 2009); y, por otro lado, coincide con el criterio de Nicholls-Estrada (2008) referente a que la mayor superficie cubierta por el pastizal favorece las dinámicas poblacionales de los enemigos naturales, y que exista un mayor índice del potencial de regulación biótica.
Los resultados descritos indican que es necesario dilucidar, en la práctica, la repercusión del complejo arvenses-cultivos-insectos (fitófagos y beneficiosos) en los diferentes agroecosistemas, con énfasis en los ganaderos; ya que, como plantearon Blanco y Leyva (2009) cuando estudiaron las arvenses y su entomofauna asociada en el cultivo del maíz posteriormente al período de competencia, estas son de gran importancia para la agroecología. Ello permite deducir que, con esa respuesta, se pudiera conocer mejor por qué un agente nocivo considerado como dañino no alcanza esa condición desde el punto de vista económico.
Así, es posible considerar los grupos de arvenses dentro de los agroecosistemas como corredores biológicos, es decir, constituyen una biodiversidad natural donde los insectos beneficiosos encuentran refugio y alimento alternativo, y ejercen su dominio atacando a los insectos plagas (Tapia-Mayer, 2013), lo cual indica una interacción trófica directa (Norris y Kogan, 2000).
Además, se requiere tener en cuenta lo informado por Vázquez et al. (2004), quienes señalan que las arvenses constituyen plantas hospedantes o «seductoras», que influyen en el comportamiento de los organismos beneficiosos y, de manera particular, en el tipo, la abundancia y el tiempo de colonización de los parasitoides (Waage y Greathead, 1986). Al mismo tiempo, ofrecen muchos recursos a los enemigos naturales, tales como: presas u hospedantes alternativos, polen o néctar, y microhábitats que no están disponibles en el monocultivo (Altieri y Nicholls, 2007) y sí en los sistemas complejos (silvopastoriles), en los que confluyen varias especies vegetales y animales.
Conclusiones
Cinco arvenses y otra Asterácea, las cuales representaron el 15 % del total, se consideran como posibles hospedantes de 27 de los controles biológicos capturados (11 depredadores, 12 parasitoides y 4 beneficiosos, también polinizadores). Entre los depredadores sobresalieron los coccinélidos C. cacti, C. maculata, C. sanguinea limbifer, D. ochroderus y D. roseicollis como biorreguladores de H. cubana y el antocórido O. pumilio, controlador de F. tritici y Podothrips sp. De los parasitoides, se destacaron el ineumónido E. purgatus y los cálcidos Conura spp., que regulan a S. frugiperda; asimismo, se distinguió la abeja A. melifera como polinizador.
Se demostró que es necesario determinar cuáles son las especies de arvenses que conviene que estén presentes en los SSP (quizás como corredores biológicos naturales) con el fin de potenciar los procesos ecológicos que en estos suceden, sobre la base de la agrodiversidad, el área que cubren los pastos predominantes y el manejo de estos sistemas productivos; ello contribuiría a propiciar la regulación de las principales plagas asociadas y a mantener el equilibrio biológico, con vista a alcanzar la sostenibilidad económica y ambiental de dichos agroecosistemas en el tiempo.

