










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Curriculum ScienTI
Curriculum ScienTI Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión impresa ISSN 1010-2752
Rev. Protección Veg. vol.27 no.2 La Habana mayo-ago. 2012
ARTÍCULO ORIGINAL
Pochonia chlamydosporia (Goddard) Zare y Gams como potencial agente de control biológico de Meloidogyne enterolobii (Yang y Eisenback) en cultivos hortícolas
Pochonia chlamydosporia (Goddard) Zare & Gams as potential biological control agent of Meloidogyne enterolobii (Yang & Eisenback) in vegetable crops
Jersys ArévaloI, S.D. SilvaII, Marina D.G. CarneiroIII, R.B. LopesIII, Regina M.D.G. CarneiroIII, Myrian S. TiganoIII, L. Hidalgo-DíazI
ICentro Nacional de Sanidad Agropecuaria, Apartado 10, San José de las Lajas, Mayabeque, Cuba. Correo electrónico: jersys@censa.edu.cu.
IIUniversidad de Brasilia UnB, Brasilia-DF, Brasil.
IIIEmbrapa- Recursos Genéticos e Biotecnologia, Brasilia-DF, Brasil.
RESUMEN
Recientemente se informaron aislamientos de Pochonia chlamydosporia (Goddard) Zare y Gams parasitando huevos de Meloidogyne enterolobii Yang y Eisenback (syn. jun. Meloidogyne mayaguensis Rammah y Hirschmann) en plantaciones de guayaba (Psidium guajava L.), al nordeste de Brasil. Teniendo en cuenta el peligro potencial de esta plaga para numerosos cultivos incluyendo las hortalizas, se realizó un experimento en casa de vegetación con dos repeticiones en el tiempo. El objetivo fue evaluar las actividades saprofítica y patogénica de tres cepas brasileñas (CG1003, CG1006 y CG1041) y una cubana (IMI SD 187) de P. chlamydosporia frente a M. enterolobii, en una sucesión de cultivos tomate (Solanum lycopersicum L.)- lechuga (Lactuca sativa L.). Al finalizar el ciclo de los cultivos, 6 meses después de la aplicación, se comprobó que la colonización de P. chlamydosporia se mantuvo en el orden de 104 UFC.g-1 de suelo y mayor de 103 UFC.g-1 en las raíces, excepto para la cepa CG1041 (102 UFC.g-1 de raíz). El porcentaje de colonización de estos hongos sobre las masas de huevos fue superior al 60% en todos los tratamientos, mientras que los porcentajes de parasitismo de huevos estuvieron entre 45 y 55%. Estos resultados validaron la cepa cubana frente a una diana y en condiciones diferentes a las que es originaria y se demostró que las cepas brasileñas son potenciales biocontroladores de M. enterolobii, abriéndose así futuras perspectivas de investigación como agentes de control biológico integrados para el manejo de este nematodo.
Palabras clave: hongos nematófagos, nematodos formadores de agallas, hortalizas.
ABSTRACT
Recently, Pochonia chlamydosporia (Goddard) Zare y Gams was isolated from parasitized eggs of Meloidogyne enterolobii Yang y Eisenback (syn. jun. Meloidogyne mayaguensis Rammah y Hirschmann) in guava (Psidium guajava L.) plantations, at the northeast of Brazil. Due to the potential danger of this pest for a large group of crops, including vegetable crops, a greenhouse experiment was conducted with two repetitions in time. The aim was to evaluate the saprophytic and parasitic activities of three brazilian strains (CG1003, CG1006 and CG1041) and the cuban strain (IMI SD 187) of P. chlamydosporia against M. enterolobii in a tomato (Solanum lycopersicum L.)- lettuce (Lactuca sativa L.) crop sequence. At the end of the cultivation, 6 month after the application, colonization of P. chlamydosporia was in the order of 104 CFU.g-1 of soil and 103 CFU.g-1 of root, with exception of the strain CG1041 (102 CFU.g-1 of root). Egg mass percentage was higher than 60% in all treatments, while parasitized egg percentage was between 45-55%. These results validated the cuban strain against a nematode target under different conditions from those it originally came from; the brazilian strain potential as biocontroler of M. enterolobii was also demonstred. Future investigation perspectives with those fungi are opened for their use as biological control agents for nematode pest management strategies.
Key words: nematophagous fungi, root-knot nematodes, vegetable.
INTRODUCCIÓN
El nematodo formador de agallas Meloidogyne enterolobii Yang y Eisenback (syn. jun. Meloidogyne mayaguensis Rammah y Hirschmann) constituye una importante plaga en países de las zonas tropical y subtropical, donde parasita de manera natural más de 30 especies de plantas como guayaba (Psidium guajava L.), cafeto (Coffea spp.), hortalizas, ornamentales y arvenses. Se considera una de las especies más peligrosas del género por su habilidad de reproducirse en plantas que poseen genes de resistencia a Meloidogyne spp. Su distribución ocurrió relativamente rápido debido a factores antropogénicos y puede hallarse en poblaciones concomitantes junto a Meloidogyne incognita (Kofoid y White) Chitwood, Meloidogyne arenaria (Neal) Chitwood y Meloidogyne javanica (Treub) Chitwood (1), lo que hace complejo su diagnóstico y manejo.
En Brasil, este nematodo se ha encontrado en áreas agrícolas (2) y forestales (3). Su diseminación a través de diversos estados, en cultivos de importancia económica, ocasionó pérdidas millonarias en la fruticultura de este país (4). En Cuba se confirmó su presencia en plantaciones de café de la región oriental del país, posteriormente se informó afectando tomate (Solanum lycopersicum L.) en la región occidental y en sistemas de producción protegida de hortalizas en el centro del país (1). Para su manejo se trabaja en la evaluación de medidas efectivas y compatibles con el ambiente, entre las que el control biológico resulta importante (5,6).
El hongo nematófago Pochonia chlamydosporia (Goddard) Zare y Gams (ex Verticillium chlamydosporium Goddard), es un parásito facultativo de huevos de nematodos de quistes y formadores de agallas presente en suelos supresores de nematodos (7). Su potencial como agente de control biológico para el manejo de Meloidogyne spp. se ha estudiado ampliamente (8, 9).
En Cuba, Hidalgo et al. (10) informaron el aislamiento de cepas autóctonas a partir de huevos de M. incognita y seleccionaron entre ellas, la cepa IMI SD 187 de P. chlamydosporia var. catenulata (Kamyscho ex Barron y Onions) Zare y Gams. Esta cepa constituye el ingrediente activo del bionematicida KlamiC®, registrado en el país (11) y cuya efectividad en el manejo de nematodos formadores de agallas se ha comprobado en condiciones de producción de cultivos protegidos y campo (12, 13).
En Brasil, recientemente se obtuvieron aislamientos nativos de P. chlamydosporia parasitando huevos de M. enterolobii en una plantación de guayaba severamente afectada, en condiciones de clima semiárido y altas temperaturas (5).
El objetivo del presente trabajo fue evaluar las actividades saprofítica y parasítica de cuatro cepas de P. chlamydosporia frente a M. enterolobii en una sucesión de cultivos tomate-lechuga (Lactuca sativa L.).
MATERIALES Y MÉTODOS
Se evaluaron las actividades saprofítica y patogénica de cuatro cepas de P. chlamydosporia (Tabla 1), frente a M. enterolobii en una sucesión de cultivos tomate (Solanum lycopersicum L.)- lechuga (Lactuca sativa L.). El experimento se realizó en casa de vegetación durante seis meses, con dos repeticiones en el tiempo. Se establecieron cinco tratamientos, correspondientes a cada cepa probada y uno control (sin P. chlamydosporia), con 8 réplicas por tratamiento, colocadas completamente al azar.
El inóculo fúngico de cada cepa se obtuvo mediante Fermentación en Estado Sólido (FES) (14). El experimento se realizó en bolsas de polietileno con 4kg de sustrato esterilizado (suelo y abono orgánico en proporción 1: 1 v/v). En cada bolsa se trasplantó una plántula de tomate (Solanum lycopersicum L.) cv. Santa Clara de 30 días de germinada. Al momento del trasplante, se inoculó una suspensión de 5000 clamidosporas viables por cada gramo de sustrato, suspendidas en 100mL de agua potable esterilizada, en el hueco de siembra de cada plántula. Transcurridos 10 días posteriores al trasplante se inoculó una suspensión de 5000 huevos de M. enterolobii alrededor de la raíz de cada planta. Después de cuatro meses, se removieron las plantas de tomate de cada bolsa y se sembró en el mismo sitio una plántula de lechuga (Lactuca sativa L.) cv. Crespa de 20 días de germinada, la cual se mantuvo durante los dos meses siguientes.
Se utilizaron cuatro bolsas de cada tratamiento para evaluar la colonización de P. chlamydosporia en el suelo y las raíces, mediante el método de dilución y siembra en medio semiselectivo (15). Durante el ciclo de tomate, se extrajo una muestra de 1g de suelo de cada bolsa a intervalos de 30 días y al final de ambos cultivos (120 días tomate y 60 días lechuga) y se determinó la dinámica de colonización del hongo en el suelo de la rizosfera durante los 180 días del experimento. En las raíces se analizó la colonización final en ambos cultivos.
Al finalizar el cultivo de tomate, se extrajeron 50 masas de huevos de cada planta con ayuda de un estéreo microscopio. Las masas se colocaron en un eppendorf con un mL de agua destilada y se lavaron tres veces con agua destilada estéril. Se sembraron 20 masas de huevos en dos placas Petri que contenían Agar Agua con Antibiótico (AAA) (Agar Técnico: 15g.L-1, Cloranfenicol: 50mg.L-1, Tetraciclina: 50mg.L-1 y Sulfato de Estreptomicina: 50mg.L-1) a razón de 10 masas de huevos por placa. Las placas se incubaron durante 5 días a 25ºC. Luego se evaluó el porcentaje de colonización de cada cepa de P. chlamydosporia. Con el resto de las masas de huevos se preparó una suspensión de huevos en 1mL de agua esterilizada -mediante ruptura mecánica de las masas. De esta se tomó una alícuota de 0,2mL y se colocó en placas con AAA y se incubó durante 24-48h a 25ºC. Posteriormente se contaron 30 huevos (parasitados y no parasitados) en cada placa y se determinó el porcentaje de huevos parasitados.
Al final de cada cultivo, las raíces de las cuatro réplicas restantes, se lavaron cuidadosamente con agua corriente para retirar los restos de suelo y se colocaron sobre papel absorbente para eliminar el exceso de agua. Inmediatamente se determinó su masa fresca y el índice de agallamiento, mediante la escala de 0 a 5 grados propuesta por Taylor y Sasser (16). Los huevos de M. enterolobii de cada raíz se extrajeron con hipoclorito al 1 % (17) y se colectaron en un tamiz de 500 Mesh en un volumen de 100mL de agua. Posteriormente se realizó el conteo de huevos en cámara de Peter por planta en el microscopio óptico (4X) y se determinó el número de huevos por gramo de raíz. Finalmente se estimó el número de huevos sanos en las raíces, según Sorribas et al. (18).
Los resultados de la concentración del hongo en suelo y raíces (UFC.g-1) se transformaron en LOG(x+1). La fórmula arcosenoÖ(x/100) se utilizó para transformar los valores del porcentaje de colonización de P. chlamydosporia sobre las masas de huevos y el parasitismo de huevos. Se realizó un ANOVA Simple y para conocer si existían diferencias significativas se utilizó la prueba de rangos múltiples de Duncan (nivel de significación p£0,05). Los análisis se realizaron mediante el paquete estadístico InfoStat versión 1.1 (19).
RESULTADOS Y DISCUSIÓN
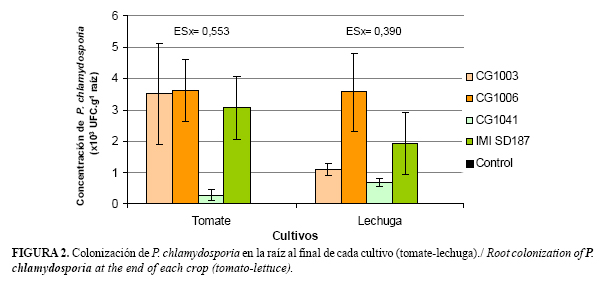
En la Figura 1 se muestran los resultados de la dinámica de la concentración de P. chlamydosporia en suelo en la sucesión de cultivo tomate-lechuga. Todas las cepas evaluadas demostraron su habilidad para desarrollarse saprofíticamente en el suelo. Los mayores valores de colonización en el tomate se alcanzaron a los 60 días después de la inoculación. Se comprobó la persistencia de estos hongos en el orden de 104 UFC.g-1 de suelo, después de 6 meses de su aplicación. La colonización de la cepa CG1041 al final del ciclo del tomate (120 días) fue significativamente inferior al resto; sin embargo no hubo diferencias significativas en la concentración de las cuatro cepas al final del cultivo de la lechuga (180 días).
En cuanto a la colonización de la raíz de ambos cultivos, los resultados se relacionan con los anteriores, ya que la cepa CG1041 alcanzó valores significativamente bajos (2,50x102 UFC.g-1 de raíz) en el cultivo del tomate, mostrando un incremento a 6,69x102 UFC.g-1 de raíz en la lechuga. El resto de las cepas mantuvo la colonización en el orden de 103 UFC.g-1 de raíces (Figura 2).
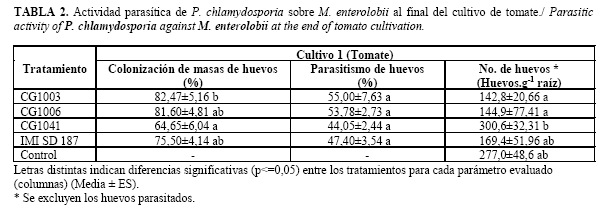
Respecto al porcentaje de colonización de los hongos sobre las masas de huevos de M. enterolobii, éste fue superior al 60% en todas las cepas, sobresaliendo CG1006 y CG1003 (Brasil) que alcanzaron más de 80% de colonización. Mientras que los porcentajes de parasitismo de huevos de cada cepa no mostraron diferencias significativas y oscilaron entre 44 y 55%. La cepa nativa CG1041 presentó los menores porcentajes de colonización de masas de huevos y parasitismo sobre los huevos del nematodo (Tabla 2).
Al estimar el número de huevos sanos en las raíces de tomate, se comprobó que aunque las cepas CG1003, CG1006 e IMI SD 187 lograron una reducción respecto al control sin P. chlamydosporia, las diferencias no fueron significativas. Por su parte, la cepa CG1041 no redujo la cantidad de huevos sanos, valor que fue superior al de otras cepas y el control (Tabla 2).
Las cepas de P. chlamydosporia no redujeron significativamente el índice de agallamiento sobre los cultivos durante la sucesión tomate-lechuga. La infestación de M. enterolobii alcanzó grados de 4-5 en el tomate y 4-4,8 en la lechuga, sin diferencias significativas respecto al control sin hongo.
Los resultados obtenidos confirman el potencial como agentes de control biológico, de cepas originarias de Cuba y Brasil de Pochonia chlamydosporia frente a M. enterolobii en una sucesión de cultivos tomate-lechuga con una sola aplicación inicial de 5000 clamidosporas.g-1 de suelo. La colonización de estas cepas, en el orden de 104UFC.g-1 en suelo y 103 UFC.g-1 en las raíces, coinciden con resultados informados para la cepa IMI SD 187, cuyos valores de colonización de masas de huevos y parasitismo de huevos sobre M. incognita fueron superiores al 80 % (20). Peteira et al. (21) obtuvieron similares resultados de parasitismo (72%) con igual dosis de aplicación, después de dos ciclos de tomate.
Es de resaltar que la cepa CG1006 mantuvo elevada colonización sobre las raíces en ambos cultivos. Sin embargo la colonización de la cepa CG1003 fue menor en las raíces de lechuga que en tomate; contrario a lo observado con CG1041. La habilidad de los aislamientos de colonizar la rizosfera de las plantas constituye un elemento clave en la selección de cepas promisorias y el establecimiento de una adecuada secuencia de cultivos (22,23).
Las actividades saprofítica y patogénica de las cepas nativas CG1003 y CG1006 frente a M. enterolobii no mostraron diferencias significativas al final del ciclo de cada cultivo en comparación con la cepa foránea IMI SD 187, de una región distinta y aislada de un hospedante diferente; esta cepa presentó un 75 % de colonización sobre las masas de huevos y un 47,4% de parasitismo sobre huevos. El hecho de contar con cepas seleccionadas que tengan alta patogenicidad sobre varias especies de Meloidogyne y que estas pudieran co-inocularse como sugiere Atkins et al. (24), podría resultar muy útil en el manejo de poblaciones concomitantes de Meloidogyne spp.
A pesar de que las cepas tuvieron buena capacidad para establecerse en la rizosfera de los cultivos hortícolas evaluados y mostraron índices de parasitismo satisfactorios (50%) no se redujo la población de M. enterolobii. Esto pudiera deberse al alto porcentaje de juveniles que escapan a la infección y que pueden completar alrededor de 4 generaciones en cultivos susceptibles (15) (4 meses tomate y 2 meses lechuga). Por otra parte, un agente de control biológico por sí solo no alcanza resultados inmediatos en la reducción de las poblaciones de nematodos, sino como parte de una estrategia integrada de manejo (9). Para Sorribas et al. (18), el recurrir a múltiples aplicaciones del hongo en un mismo cultivo pueden reducir el índice de agallas.
Las plantas de lechuga tuvieron un pobre desarrollo radicular durante el experimento, como resultado de la alta susceptibilidad del cultivo al nematodo (15). Al finalizar el cultivo, el peso de las raíces estuvo entre 4-5 gramos, siendo el material escaso para evaluar la colonización de las cepas de P. chlamydosporia sobre las masas de huevos y el parasitismo de huevos de M. enterolobii.
Los resultados mostraron que la cepa cubana es efectiva frente a una diana y en condiciones diferentes de las que se aisló. Se demostraron además las potencialidades de las cepas brasileñas frente a M. enterolobii en cultivos hortícolas. Finalmente y tomando en cuenta que tanto el contenido de abono orgánico -fuente adicional de nutrientes- como la microbiota natural de un suelo pueden hacer a éste más receptivo para aislamientos nativos de Pochonia e influir en su actividad nematófaga (25, 26), es necesario evaluar el efecto de tales factores sobre el desarrollo saprofítico y parasítico de las cepas de P. chlamydosporia frente a poblaciones de nematodos en suelo natural. Dicha evaluación es importante para la selección, uso e integración de agentes de control biológico dentro de estrategias de manejo de Meloidogyne spp.
REFERENCIAS
1. Rodríguez MG, Gómez L, Peteira B. Meloidogyne mayaguensis Rammah y Hirschmann, plaga emergente para la agricultura tropical y subtropical. Rev Protección Veg. 2007;22(3):183-198.
2. Carneiro RMDG, Moreira WA, Almeida MRA, Gomes ACMM. Primeiro registro de Meloidogyne mayaguensis em goiabeira no Brasil. Nematologia Brasileira. 2001;25:223-228.
3. Lima IM, Souza RM, Silva CP, Carneiro RMDG. Meloidogyne spp. from preserved areas of Atlantic Forest in the State of Rio de Janeiro, Brazil. Nematologia Brasileira. 2005;29:31-38.
4. Pereira FOM, Souza RM, Souza PM, Dolinski C, Santos GK. Estimativa do impacto económico e social direto de Meloidogyne mayaguensis na cultura da goiaba no Brasil. Nematologia Brasileira. 2009;33:176-181.
5. Arévalo J, Hidalgo-Díaz L, Martins I, Souza JF, Castro JMC, Carneiro RMDG, et al. Cultural and morphological characterization of Pochonia chlamydosporia and Lecanicillium psalliotae isolated from Meloidogyne mayaguensis eggs in Brazil. Tropical Plant Pathology. 2009;34(3):158-163.
6. Carneiro RMDG, Hidalgo-Díaz L, Martins I, Silva KFA, De Sousa MG, Tigano MS. Effect of nematophagous fungi on reproduction of Meloidogyne enterolobii on guava (Psidium guajava) plants. Nematology. 2011;13(6):721-728.
7. Kerry BR. Exploitation of the Nematophagous Fungus Verticillium chlamydosporium Goddard for the Biological Control of Root-knot Nematodes (Meloidogyne spp.). In: Butt TM, Jackson C, Magan N, editors. Fungi as Biocontrol Agents: Progress, Problems and Potential. CAB International, Wallingford; 2001. p. 155-168.
8. Atkins SD, Hidalgo-Díaz L, Clark IM, Morton CO, Montes de Oca N, Gray PA, et al. Approaches for monitoring the release of P. chlamydosporia var. catenulata, a biological control agent of root-knot nematodes. Mycol Res. 2003;107(2):206-212.
9. Hidalgo-Díaz L, Kerry BR. Integration of biological control with other methods of nematode management. In: Ciancio A, Mukerji KG, editors. Integrated management and biocontrol of vegetable and grain crops nematodes. 2008; p.29-49.
10.Hidalgo L, Sánchez L, Gómez L. Verticillium chlamydosporium Goddard, parásito de huevos de Meloidogyne incognita. Rev Protección Veg. 1998;13(1):29-30.
11.Lista Oficial de Plaguicidas Autorizados en la República de Cuba. 2010. p. 325.
12.Delgado Y. Validación de la Efectividad Técnica y Económica de dos Bionematicidas en Condiciones de Cultivos Protegidos. Tesis. MINAG-IIFT; 2010.
13.Hernández MA, Hidalgo L. KlamiC®: Bionematicida agrícola producido a partir del hongo Pochonia chlamydosporia var. catenulata. Rev Protección Veg. 2008;23(2):131-134.
14.Montes de Oca N. Buenas prácticas de fabricación para la obtención de un bionematicida a partir de la cepa IMI 187 de Pochonia chlamydosporia var. catenulata. Tesis. La Habana: Universidad Agraria de La Habana; 2004.
15.Kerry BR, Bourne JM. A manual for research on Verticillium chlamydosporium, a potential biological control agent for root-knot nematodes. Ghent, Belgium, IOBC/WPRS. 2002. p. 84.
16.Taylor AL, Sasser JN. Biology, identification and control of rootknot nematodes (Meloidogyne species). North Carolina State Univ. & U.S. Agency of International Development, Raleigh. 1978; 111p.
17.Hussey RS, Barker KB. A comparison of methods of colleting inocula of Meloidogyne spp. including a new technique. Plant Dis Reptr. 1973;57:1025-1028.
18.Sorribas FJ, Ornat C, Galeano M, Verdejo-Lucas S. Evaluation of a native and introduced isolate of Pochonia chlamydosporia against Meloidogyne javanica. Biocontrol Science and Technology. 2003;13(8):707-714.
19.Di Rienzo JA, Balzarini MG, González L, Tablada M, Guzmán W, Casanoves F, et al. InfoStat/Profesional Versión 1.1. 2002.
20.Puertas A, Arévalo J, Montes de Oca N, Miranda I, Hidalgo-Díaz L. Efecto de diferentes concentraciones de inóculo de la cepa IMI SD 187 de Pochonia chlamydosporia var. catenulata en el control de Meloidogyne incognita. Rev Protección Veg. 2006;21(2):74-79.
21.Peteira B, Puertas A, Hidalgo-Díaz L, Hirsch PR, Kerry BR, Atkins SD. Real-time PCR to monitor and assess the efficacy of the nematophagous fungus Pochonia chlamydosporia var. catenulata against root-knot nematode populations in the field. Biotecnología Aplicada. 2005;22(4):261-266.
22.Puertas A, Hidalgo-Díaz L. Influencia de la planta hospedante y su interacción con Meloidogyne incognita sobre la efectividad de Pochonia chlamydosporia var. catenulata. Rev Protección Veg. 2007;22(2):104-109.
23.Leij de FAAM, Kerry BR. The nematophagous fungus Verticillum chlamydosporium Goddard, as a potencial biological control agent for Meloidogyne arenaria (Neal) Chitwood. Revue Nématol. 1991;14:157-164.
24.Atkins SD, Peteira B, Clark IM, Kerry BR, Hirsch PR. Use of real-time quantitative PCR to investigate root and gall colonisation by co-inoculated isolates of the nematophagous fungus Pochonia chlamydosporia. Ann Appl Biol. 2009;155:143-152.
25.Monfort E, López-llorca LV, Janson HB, Salinas J. In vitro soil receptivity assays to egg-parasitic nematophagous fungi. Mycological Progress. 2006;5:18-23.
26.Podestá GS, Dallemole-Giaretta R, Freitas LG, Lopes EA, Ferraz S, Zooca RJF. Atividade de Pochonia chlamydosporia em solo natural ou autoclavado sobre Meloidogyne javanica. Nematología Brasileira. 2009;33(2):191-193.
Recibido: 20-11-2011.
Aceptado: 21-3-2012.

{kind=link}
{kind=link}
{kind=link}
{kind=link}