










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl vol.35 no.1 La Habana ene.-mar. 2018
RESEARCH
Identification and characterization of a Beauveria bassiana (Bals.) Vuill. isolate having a high potential for the control of the Diatraea sp. sugarcane stem borer
Identificación y caracterización de un aislamiento de Beauveria bassiana (Bals.) Vuill. con alto potencial para el control del barrenador del tallo de la caña de azúcar Diatraea spp.
Jennifer L García1, Paula Sotelo1, Diana M Monroy1, Gloria Barrera1, Juliana Gómez-Valderrama1, Carlos Espinel1, Emiliano Barreto2, Laura F Villamizar3
1 Centro de Investigación Tibaitatá, Corporación Colombiana de Investigación Agropecuaria (AGROSAVIA). Km 14 vía Mosquera-Bogotá, Mosquera, Colombia.
2 Instituto de Biotecnología IBUN, Universidad Nacional de Colombia, Sede Bogotá, Colombia.
3 Forage Science, AgResearch Ltd. Lincoln Research Centre. Christchurch 8140, New Zealand.
ABSTRACT
A native isolate of the Beauveria sp. fungus obtained from the Diatraea sp. larva was studied and identified as B. bassiana by analyses based on ribosomal internal transcribed spacer (ITS) sequences. The code given to the isolate was Bv062 and its insecticidal activity was evaluated on the second instar larvae of D. saccharalis, D. indigenella, D. tabernella and D. busckella, under laboratory conditions, producing mortality rates of 73.3, 83.3, 67.6 and 24.1 %, respectively. Subsequently, the activity of the fungus was evaluated on D. saccharalis eggs. Egg mortality was of 82.2 % when spraying a conidial suspension adjusted to aconcentration of 1 × 108 conidia/mL on the eggs. The mean lethal concentration (LC50) was found to be 9.04 × 105 conidia/mL and the lethal concentration ninety (LC90) was of 2.07 × 107 conidia/mL on the second instar larvae of D. saccharalis. In-plant bioassays under semi-controlled conditions showed that fungus efficacy was above 50 % with the sporulation of dead larvae of 80 %, using concentrations between 1.0 × 107 and 5.0 × 107 conidia/mL. Results demonstrated the high potential of B. bassiana Bv062 asa biocontrol agent for the development of a bio-insecticide to be used in the management of different species of the sugarcane stem borer complex.
Keywords: entomopathogenic fungi, biological control, Diatraea saccharalis.
RESUMEN
Se estudió un aislamiento native del hongo Beauveria sp. obtenido de una larva de Diatraea sp., el cual fue identificado molecularmente como B. bassiana mediante el análisis basado en las secuencias de la región espaciadora interna transcrita del ADN ribosomal (ITS). El aislamiento de B. bassiana fue codificado como Bv062 y se evaluó su actividad insecticida sobre larvas de segundo ínstar de D. saccharalis, D. indigenella, D. tabernella y D. busckella en condiciones de laboratorio, donde causó mortalidadesdel 73.3, 83.3, 67.6 y 24.1 % respectivamente. Posteriormente se evaluó la actividad del hongo sobre huevos de D. saccharalis y se encontró una mortalidad de huevos del 82.2 % al asperjarlos con una suspensión de conidios ajustada a una concentración de 1× 108 conidios/mL. Se determinó una concentración letal media (CL50) de 9.04 × 105 conidios/mL y una concentración letal noventa (CL90) de 2.07 × 107 conidios/mL sobre larvas de segundo ínstar de D. saccharalis. Ensayos de actividad in planta en condiciones semicontroladas evidenciaron una eficacia del hongo superior al 50% con una esporulación de las larvas muertas superior al 80 %, al utilizar concentraciones entre 1.0 × 107 y 5.0 × 107 conidios/mL. Los resultados demostraron el alto potencial de B. bassiana Bv062 como agente de biocontrol para el futuro desarrollo de un bioinsecticida para el manejo de diferentes especies del complejo del barrenador del tallo de la caña de azúcar en este y otros cultivos hospederos.
Palabras clave: hongos entomopatógenos, control biológico, Diatraea saccharalis.
INTRODUCTION
Diatraea saccharalis (Fabricius) is known to be one of the most economically important pests in sugarcane (Saccharum officinarum) production, where significant losses in yield are found in many parts of the United States and South America [1, 2]. This insect causes direct damage to the sugarcane by penetrating and opening galleries in the stem. The damaged tissue affects the weight of sugar production and produces the deterioration of the stems used as seed, as well as indirect damages occurring by allowing the entrance to opportunistic fungi that digest sugar and contaminate the juice, thereby decreasing its quality for sugar production [1, 3]. In Colombia there is a complex of borer species that affects sugarcane plantations in different areas of the country, with the presence of up to six species in the same region [1]. The losses produced by the insect are estimated as a reduction of 145 kg of unit, where the estimated decrease in tons of harvested sugarcane is around 0.826 % [4]. Agrochemicals have been used to control this pest, most of them have been inefficient due to the insect’s cryptic habits, since its larval cycle develops within sugarcane stems [5]. Furthermore, the high toxicity and adverse effects of agrochemicals on the environment and human health have led to the search of other environmentally friendly alternatives. Biological control through natural enemies such as parasitoids and predators has been mostly used for pest control [6]; nonetheless, different factors have affected the efficacy of this strategy for the control of the borer complex. One of them is the presence of different species of Diatraea spp., for which there is a differential response to the parasitoids, indicating the need for several approaches in the pest management of these new species [7].
At the same time, some controller species cannot be produced massively, or their extensive use is limited, and they must be preserved in the agroecological system, which is, however, hampered by the indiscriminate use of agrochemicals and the disappearance of wild plants [1]. For this reason the use of entomopathogenic fungi as a complement to biological control with parasitoids is an interesting tool that should be included in integrated pest management programs; this is due to their high pathogenicity against different species, the fact that they may be produced and formulated at an industrial scale and that they can generate natural epizootic diseases in the field, as well as their natural dissemination in the environment [8, 9]. Several fungi (Hypocreales: Clavicipitaceae) including Beauveria bassiana, B. brongniartii, M. anisopliae, I. fumosorosea, Hirsutella sp., Cylindrocarpon sp., and M. rileyi have been isolated from Diatraea spp. And have become important agents for their microbial control in America [6]. In Colombia, in spite of its high potential, this strategy has been poorly used, and currently there is no biological pesticide registered for the control of any of the species of the sugarcane borer [10]. This underlines the importance of exploring the use of entomopathogenic fungi that may be developed into biopesticides for the control of Diatraea spp.
Among the previously mentioned fungi, Beauveria bassiana (Balsamo) Vuillemin (Hyphomycetes: Moniliales) is one of the best known, widely distributed and used entomopathogenic fungi species for the control of many crop pests worldwide. It is a ubiquitous filamentous fungus with a high potential for the control of insects of different orders, such as Hepteroptera, Lepidoptera, Coleoptera, Hymenoptera and Diptera [8]. Its action mechanism is through the contact with the cuticle of the host, where it penetrates the exoskeleton reaching the hemocoel, through enzymatic mechanisms; later it invades the hemolymph where it consumes the necessary nutrients for its growth, thus colonizing all tissues and producing the death of the insect [11]. In this paper, an isolate of Beauveria sp. that had been recovered from a Diatraea spp. larva that was naturally infected in the field and obtained in a sugarcane plantation for panela (a piece of unrefined whole cane sugar) production in the Department of Boyacá (Colombia), was identified and characterized, determining its potential as a biocontrol agent of the stem borer complex.
MATERIALS AND METHODS
Microorganism and culture medium
The microorganism used in this research was the isolate of Beauveria bassiana obtained from one larva of Diatraea sp. coming from a sugarcane plantation and collected in the Department of Boyacá; it was supplied by the Germplasm Bank for Microorganisms of Interest in Biological Control of the Colombian Corporation for Agricultural Research (AGROSAVIA), and coded as Bv062. The B. bassiana strain was grown in potato-dextrose agar (PDA), supplemented with 0.1 % chloramphenicol and incubated for 7 days at 25 °C.
Insects
The experiments were carried out with D. saccharalis larvae from the colony located at the Insect Rearing Unit of AGROSAVIA at the Research Center “Tibaitatá” (Mosquera-Colombia), which was established with larvae donated by Cenicaña (Colombian sugarcane Research Center). The larvae of D. tabernella, D. indigenella and D. busckella were also donated by Cenicaña from its rearing unit located at the Experimental Center in the Cauca Valley (Cali-Florida). The larvae were kept at 25 ± 5 ºC with a relative humidity of 60 ± 10 % and a photoperiod of 16 h of light: 8 h of darkness, using an artificial diet described by Lastra and Gómez [12].
Ribosomal DNA amplification and sequencing
Fungal spores and mycelia were recovered with a loop from massive sporulated cultures grown on Petri dishes with PDA. The conidia (20 mg) were suspended in 200 μL of sterile water. The genomic DNA extraction was carried out with the ZR Fungal/Bacterial DNA MicroPrepTM Kit (Zymo Research) and the DNA was stored at –20 ºC until it was used. For molecular identification, amplification by PCR was made at a ribosomal DNA (rDNA) region with universal primers, a DNA ribosomal region was amplified with primers that flanked ITS. This region is widely used for genera and specie classification and to calculate genetic distance among entomopathogenic fungi [13]. PCR amplification was performed in a total volume of 25 μL, which include 50 ng genomic DNA.
The PCR product was separated in 1 % agarose gel and visualized using SYBR® Safe (Thermo Fisher Scientific) in an UV transluminator. The PCR product was purified with the Wizard® SV Gel Kit and the PCR Clean-Up System (Promega, USA) and cloned in the pGEM-T (Promega, USA) plasmid. The fragment was finally sequenced in both directions with universal T7 and M13 primers. The sequences obtained were edited and aligned to obtain a consensus sequence that was compared to the representative sequences of different fungi species from the database of the National Center for Biotechnology Information (NCBI). Phylogenetic reconstruction was made with the Neighbor-Joining method, using the nucleotide substitution method of Kimura 2 parameters, with the MEGA program version 6 [13] including 25 sequences of the genera Beauveria, Isaria, Metarhizium and Nomuraea (Table 1).
Evaluation of the insecticidal activity of B. bassiana Bv062 against larvae of different Diatraea species
A seven days-old culture of the isolate Bv062 was used to prepare a suspension that was adjusted to 1 × 107 conidia/mL, which was topically inoculated by placing 2 μL on the dorsum of second instar larvae of D. saccharalis, D. indigenella, D. tabernella and D. busckella. The inoculated larvae were individually placed in plastic half-ounce containers with a grain of tender corn as the feeding substrate. The control treatment consisted of larvae that did not undergo any treatment. The completely randomized experimental design had three replicates per treatment with 10 larvae per replicate. Larval mortality was assessed every two days until all individuals died or formed the pupa. Mortality results obtained with treatments were corrected with the mortality in the control to calculate the efficacy using the formula of Schneider-Orelli [14].
Evaluation of the insecticidal activity of B. bassiana Bv062 against D. saccharalis eggs
A bioassay was carried out using the nebulization technique described by Carrera et al. [15] consisting of the use of the PulmoMate Sunrise® nebulizer connected to a PVC tube that was the pulverization chamber of the fungal suspension. Five suspensions of conidia adjusted to five concentrations of: 5 × 106, 1 × 107, 5 × 107, 1 × 108, 5 × 108 conidia/mL in a 0.1 % Tween 80 solution, were prepared. A volume of 5 mL was taken for each treatment and sprayed on three egg-masses of D. saccharalis collected 24 hours after laying, with a known number of eggs estimated by counting through a stereoscope. The spraying lasted two minutes and the inoculated eggs were left to dry in the room. The control treatment consisted of D. saccharalis eggs sprayed with a 0.1 % solution of Tween 80. The eggs were placed individually in half-ounce plastic containers with a grain of tender corn and a piece of dampened cotton. After 7 days, the infected eggs and the neonate larvae were counted using a stereoscope. The larvae were placed individually in half-ounce plastic containers with a grain of corn. Larval mortality was assessed every two days until all individuals died or formed the pupa. To confirm fungal infection, the dead larvae were collected and placed in a humid chamber at 25 °C for approximately 7 days.
Determination of the lethal concentrations of Bv062 on D. saccharalis larvae
A bioassay was carried out following the above described methodology, for which suspensions of conidia were prepared; these were adjusted to six concentrations of 5 × 105, 1 × 106, 5 × 106, 1 × 107, 5 × 107, 1 × 108 conidia/mL with which the D. saccharalis larvae were inoculated. Each larva was placed in a plastic container containing a grain of corn; they were allotted in groups of 10 units by triplicate, in Selloplus® boxes of 64 ounces, and incubated at 26 ± 2 °C, with 70 % RH, and mortality and sporulation were evaluated every two days. The control treatment consisted in non-inoculated larvae. The results of mortality were subjected to a Probit analysis with the POLO Plus program to determine lethal concentrations. To confirm fungal infection, the dead larvae were collected and placed in humid chambers at 25 °C for approximately 7 days.
Determination of insecticidal activity of B. bassiana Bv062 against D. saccharalis larvae on sugarcane plants
The bioassay was developed in the municipality of Barbosa, Santander (Colombia), having an average temperature of 22 ± 5 °C and relative humidity (HR) of 63.5 ± 10 %. Sugarcane plants of the RD7511 variety of three months of age were planted individually in pots and sprayed with 2 mL per plant with the treatments using a manual nebulizer. The treatments consisted in conidial suspensions of B. bassiana Bv062 adjusted to three concentrations (5 × 106, 1 × 107, 5 × 107 conidia/mL) and a control treatment where plants were sprayed with water. After applications, each plant was infested with five D. saccharalis larvae of five eight-day-old (second instar), and plants were placed outdoor and protected with a poly-shade net. Thirty plants per treatment were distributed in three experimental units of 10 plants each. The experimental design was completely randomized. After eight days, a destructive sampling was carried out, in which the plant was cut longitudinally, and all larvae found were removed (alive and dead). The live larvae were placed individually in plastic half-ounce containers with a grain of tender corn and taken to the laboratory to evaluate mortality, according to the above described methodology. The efficacy of the treatments was estimated with the formula of Abbott [16].
Statistical analysis
The homogeneity of variances and the normality of results were determined through the Bartlett and Shapiro-Wilk tests with a 95 % confidence, respectively. After demonstrating normality, differences between treatments were determined by variance analysis (ANOVA) and Tukey test (95 %) through the statistical program Statistix (8.0).
RESULTS AND DISCUSSION
Molecular identification of the selected isolate For the molecular identification of the Bv062 isolate based on an ITS fragment, an amplimer of 595 bp was obtained (Figure 1).
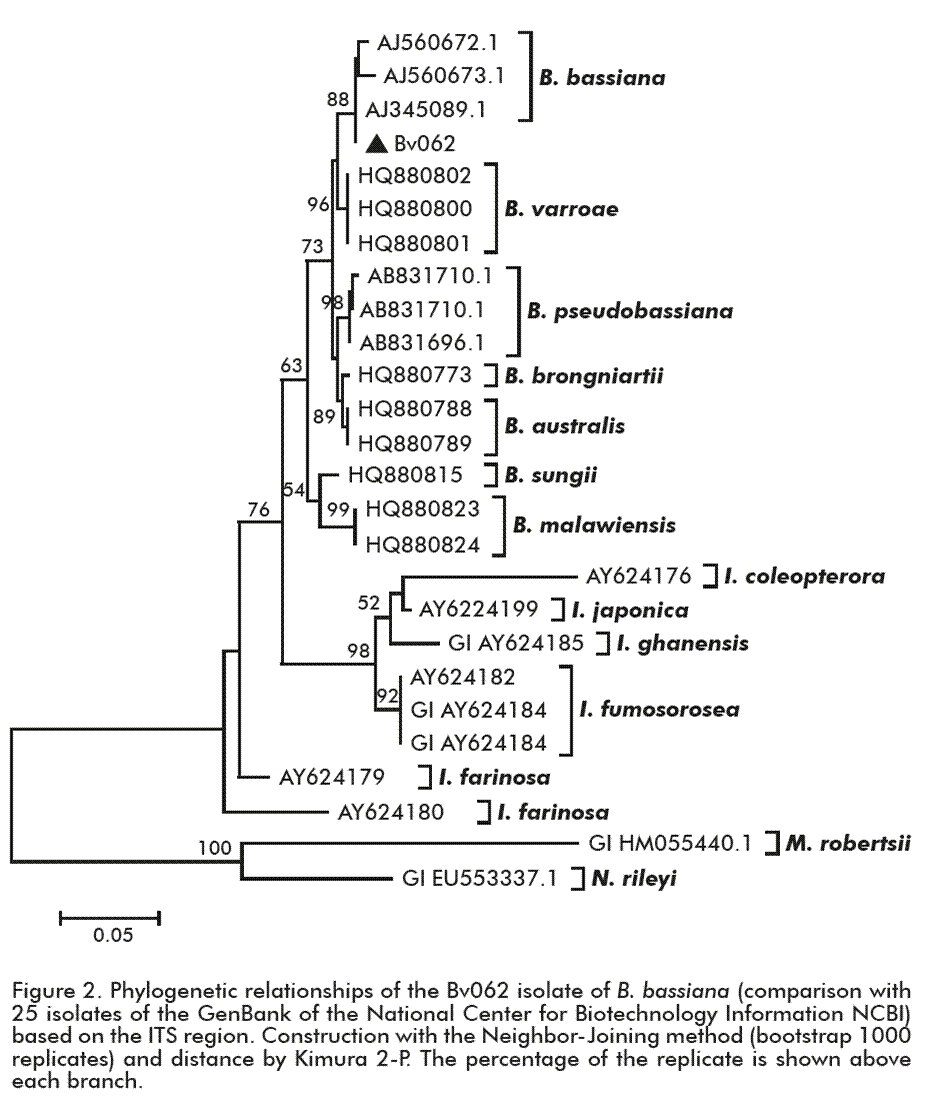
The inferred phylogenetic reconstruction with the Neighbor-Joining method using 26 isolates from the GenBank, showed that the Bv062 isolate is grouped in the same clade as other B. bassiana isolates, with a high bootstrap value (1000 replicates) (Figure 2). In the tree there is a clear separation per group of isolates of the Beauveria genus, in relation to other genera such as Isaria or Metarhizium. The genetic distance calculated with the Kimura 2-parameter method showed that the distance between the Bv062 isolate compared to the three B. bassiana isolates of different geographical origin (United Kingdom, United States and Hungary) is less than 0.007, while the distance from other Beauveria species ranges between 0.01 (B. varroae) and 0.05 (B. malawiensis) (data not shown).
Previous studies have used the regions between ITS ribosomal genes for the identification of different genera of entomopathogenic fungi, demonstrating the usefulness of this region to discriminate isolates of the Beauveria sp, Metarhizium, and Isaria [17, 18] genera. Considering its morphological characteristics and its molecular identification, the Bv062 isolate was classified as B. bassiana.
Evaluation of the insecticidal activity of B. bassiana against larvae of different Diatraea species
The isolate of B. bassiana Bv062 was pathogenic for the second instar larvae of the four borer species evaluated in this study under laboratory conditions. This was observed through the mortality of the individuals, followed by mycelial growth, and finally abundant white sporulation, which produced the mummification of the bodies. These symptoms correspond to typical signs of infection by the B. bassiana isolate [19]. The mortality of the larvae ranged from 24 to 83 % (Figure 3); these values were not significantly different between the first three species, but they were different from D. busckella (F = 18.8, df = 11, p = 0.0006), indicating a lower degree of pathogenicity of the fungus in this species compared to the others.
The B. bassiana isolate evaluated in this study was obtained from a naturally infected larva from the field, which was collected in the sugarcane plantation of the department of Boyacá (Colombia). Here the D. saccharalis, D. rosa and D. busckella species have been identified within the borer complex that affects the crop, where the latter has a higher incidence compared to the other species (unpublished data). Possibly, the coexistence of the fungus with this species in the same geographical region has generated the establishment of the B. bassiana in this host population without producing a drastic mortality because of the adaptation processes or their coevolution, resulting in a lower pathogenicity of the fungus or a greater resistance of the host [20]. Another hypothesis suggests that when there are different host populations within the same ecosystem, variations can be produced in the characteristics of the pathogen, such as its transmission, infection rate and fitness [21].
It should be noted that most of the work related to the entomological pathogenicity of the sugarcane borer reported in the literature has been carried out with the D. saccharalis species, and information is scarce on the activity of the B. bassiana on other species of the complex.
Evaluation of the insecticidal activity of B. bassiana Bv062 on eggs of D. saccharalis
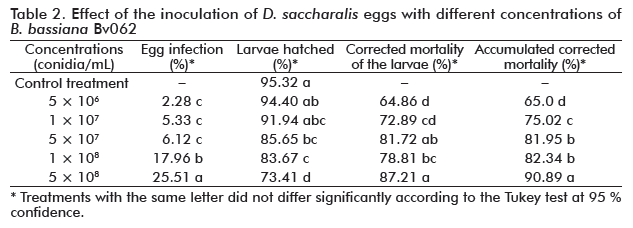
The infection of the D. saccharalis eggs produced by the B. bassiana Bv062 isolate, ranged from 2 to 25 % (Table 2), where the highest concentration, was significantly higher compared to the others (F = 49.5; df = 14; p = 0.00). Mycelial growth was observed on the surface of the eggs, and there was an evident reddish color, which is a characteristic symptom of entomopathogenic fungus infection on insect eggs, as reported by other authors [22].
In relation to the recovery of the larvae hatching from the eggs, significantly lower values were found when the eggs were treated with the fungus at the three highest concentrations, compared to the control treatment, in which hatching was of 95.32 % (F = 19.8; df = 17; p = 0.00) (Table 2). Finally, the mortality of the hatched larvae was evaluated, ranging between 65 and 91% according with the different evaluated concentrations. Larvae mortality was significantly higher when eggs were inoculated with the highest concentrations of 5 × 107, 1 × 108 and 5 × 108 conidia/mL (F = 30.6; df = 14; p = 0.00). The mortality of hatched larvae could be the result of fungal infection, due to the penetration of the fungus inside the egg, or the indirect acquisition of the inoculum from the contaminated chorion, during hatching [22].
These results demonstrate the ability of B. bassiana to produce the insecticidal effect, not only on the larvae of the insect, but also on the eggs, thereby extending the action spectrum of the fungus, since it can affect the pest insect during two stages of its development, i.e. eggs and larvae.
Determination of lethal concentrations of Bv062 on D. saccharalis larvae
The mortality of D. saccharalis larvae was directly dependent on fungal dose, which increased as the concentration of the inoculated suspension increased. During the initial days of infection, a pinkish coloring and the hardening of the body of the larva, were observed. Subsequently, a superficial mycelial growth was found on the dermis of the dead larva, until it was completely covered with the white mycelia and powdery white sporulation (Figure 4), all of which are typical symptoms confirming that the cause of death was the infection by B. bassiana.
Lethal concentrations LC50 and LC90, the confidence limits and the slope, were calculated from the average of three replicates, and the results are presented in Table 3. Lethal concentrations were lower than those found for other isolates of B. bassiana evaluated on D. saccharalis larvae. For example, Zappelini [23] determined the LC90 of 1 × 108 conidia/mL, corresponding to five times that estimated in this study for Bv062. Wenzel et al. [24] also estimated the LC50 of 1.58 × 107 conidia/mL for the Brazilian isolate IBCB 66. In another study, Alves et al. [25] obtained the LC50 of 4.8 × 106 conidia/mL for the Brazilian isolate B. bassiana 447 on the third stage larvae of D. saccharalis. However, it is not possible to compare the results obtained because of the methodological differences.
In relation to the mean lethal time (LT50), this was of six days, and lethal time ninety (LT90) was of approximately 12 days (Table 4). These values contrast with other studies such as those of Zappelini [11] who reported LT50 of 5.4 days, Wenzel et al. [24] with LT of 6 days at concentration of 1.58 × 107 conidia/mL and Díaz and Lecuona [26] with LT ranging from 2.9 to 7.78 days.
Determination of insecticidal activity of B. bassiana Bv062 against D. saccharalis larvae on sugarcane plants
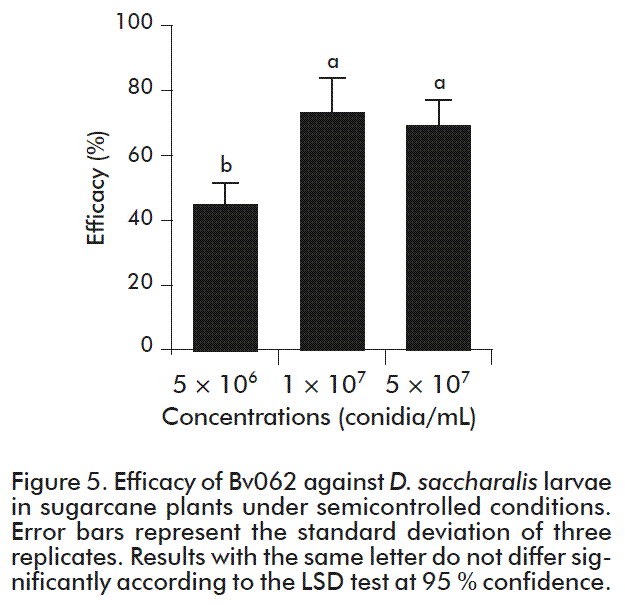
After eight days of the evaluation, a high number of dead larvae were found inside plants and on their surface. Many of them were mummified, and showed evident sporulation. These results demonstrate that the foliar application of the entomopathogenic fungus enables the infection of larvae that crawl on the foliar surface, which is a normal behavior in borer larvae during the initial instars. The corrected mortality, using the mortality of the control treatment (7.9 %) was expressed as efficacy, and it is shown in Figure 5. Efficacy showed values of 46, 75 and 69 % with concentrations of 5 × 106, 1 × 107 and 5 × 107 conidia/mL respectively.
The efficacy obtained with the highest concentration (T1) was not significantly different from that obtained with the medium concentration (T2), but it was significantly higher than that of the lowest concentration (T3) (F = 12.1; n = 2; p = 0.0078). Sporulation of dead larvae was over 80 % in the three treatments, thus confirming the cause of death as a fungal infection. Considering that there are no significant differences in efficacy with T1 or T2, but there is a numerical difference between them, the concentration of 1 × 107 conidia/mL was selected as the minimum effective dose for further evaluations of this entomopathogenic agent in the field.
Considering that the volume of insecticides applied under field conditions ranges from 200 to 400 L/ha, the dose based on the selected concentration would range from 2 × 1012 to 4 × 1012 conidia/ha. This concentration is similar to that used by Estrada et al. [27], who applied 1 × 1012 conidia/ha in a homogeneous population of this pest in a sugarcane plantation in Cuba, where results showed a significant reduction in the population levels of D. saccharalis and the damage produced by it, with an important increase in yield. In another study, a commercial myco-insecticide (Mycotrol®) was evaluated under field conditions in a sugarcane plantation in Texas (USA) using a dose of 5 × 1013 conidia/ha. Although results showed a reduction in the incidence and the damage done by D. saccharalis, this reduction was not related to greater yields or the quality of the sugarcane juice; the authors considered that there was an insufficient coverage of the biological agent on the foliar surface [28]. Furthermore, the fungus used as the active ingredient in this biological agent was not isolated from Diatraea spp., for which reason its activity on this insect and its behavior, may be inferior to other strains.
Another important factor in the possible future use of this fungus in sugarcane is the effect on non-target organisms, considering that especially for Diatraea spp. natural enemies are important in the regulation of this pest. Although this fungus has a broad spectrum of action, current evidence suggests that the impact on other, non-target organisms, is minimum because of its specificity [8]. In previous studies, the use of entomopathogenic agents such as B. bassiana and M. anisopliae showed no effect against Cotesia flavipes (Hymenoptera: Braconidae) and on Trichogramma sp. (Hymenoptera: Trichogrammatidae) [29, 30], which are the main parasitoids of D. saccharalis. This suggests that these two control tools can be used simultaneously within an integrated pest management program for sugarcane plantations, although specific studies should be carried out with the B. bassiana Bv062 isolate.
CONCLUSIONS
The B. bassiana Bv062 isolate is a potential agent for the control of eggs and larvae of D. saccharalis and some species of the borer complex affecting sugarcane in Colombia. Under controlled conditions, B. bassiana Bv062 is able to reduce the population of insect pests, which makes it a viable alternative for the development of a biopesticide. Studies should be continued on the formulation of the fungus, its evaluation under field conditions by spraying the crop, the determination of persistency, studies on non-targeted species, among others, to define the viability of its use in sugarcane plantations.
ACKNOWLEDGEMENTS
The authors would like to thank the Ministry of Agriculture and Rural Development, and the Colombian Corporation for Agricultural Research (AGROSAVIA) for funding this research. We are indebted to Dr. Germán Vargas of Cenicaña for his advisory and for providing the larvae of different species of Diatraea sp. for the bioassays. We also thank engineers Julio Ramírez and Liliana Murcia from AGROSAVIA, for their logistic approach in setting up the bioassays in sugarcane shoots and their collaboration in the initial work for the search of entomopathogenic agents. Special thanks are given to the research group of AGROSAVIA, the Research Center of Tibaitatá (Nancy Barreto, Pablo Osorio, Yuly Zandoval, and Jesús Gómez), the Cimpa office (Orlando Insuasty), who supported the initial sampling work for the search of entomopathogenic agents, and particularly to Claudia Mesa, Juan Carlos Bernal and Consuelo Rojas for their constant support in all activities of the project.
CONFLICTS OF INTEREST STATEMENT
The authors declare that there are no conflicts of interest.
REFERENCES
1. Vargas G, Gómez LA, Michaud JP. Sugarcane stem borers of the Colombian Cauca River Valley: Current pest status, biology, and control. Florida Entomol. 2015;98:728-35.
2. Guevara LA, Wiendl FM. Aspectos da biologia, en condições de campo, da broca da Cana-de-Açucar (Diatraea saccharalis (Frab,1794)). Agric Técnica. 1980;1:32-9.
3. Vargas G, Gómez L. Evaluación del daño causado por Diatraea spp. en caña de azúcar y su manejo en el valle del río Cauca. Ser Técnica Cenicaña;2005;(9):1-7.
4. Bustillo A. Insectos plaga y organismos benéficos del cultivo de la caña de azúcar en Colombia. Cali: Ceñicaña; 2013.
5. Svedese V, Tiago P, Bezerra J, Paiva L, Lima E, Porto AL. Pathogenicity of Beauveria bassiana and production of cuticle-degrading enzymes in the presence of Diatraea saccharalis cuticle. African J Biotechnol. 2013;12:6491-7.
6. Hernández-Velázquez VM, Lina-García LP, Obregón-Barboza V, Trejo-Loyo AG, Peña-Chora G. Pathogens associated with sugarcane borers, Diatraea spp. (Lepidoptera: Crambidae): A Review. Int J Zool. 2012;2012:1-12.
7. Vargas G, Lastra LA, Solís MA. First record of Diatraea tabernella (Lepidoptera: Crambidae) in the Cauca river valley of Colombia. Florida Entomol. 2013;96:1198-201.
8. Zimmermann G. Review on safety of the entomopathogenic fungus Beauveria bassiana and Beauveria brongniartii. Biocontrol Sci Technol. 2007;17:879-920.
9. Carruthers R, Soper R. Fungal diseases. In: Fux A, Tanada Y, editors. Epizootiology of insect diseases. New York: John Wiley; 1987. p. 416.
10. Instituto Colombiano Agropecuario. Productos registrados Bioinsumos-Junio 2017. Bogotá: ICA; 2017.
11. Valero-Jiménez CA, Wiegers H, Zwaan BJ, Koenraadt CJM, van Kan JAL. Genes involved in virulence of the entomopathogenic fungus Beauveria bassiana. J Invertebr Pathol. 2016;133:41-9.
12. Lastra B, Gómez L. La cría de Diatraea saccharalis (F.) para la producción masiva de sus enemigos naturales. Ser Técnica Cenicaña. 2006;36:30.
13. Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol. 2011;28:2731-9.
14. Zar J. Biostatistical Analysis. New Jersey: Prentice Hall; 1999.
15. Carrera M V, Zeddam JL, Pollet A, Léry X, López-Ferber M. Evaluation of the per os insecticidal activity of baculoviruses by a nebulization method. IOBC/wprs. 2008;31:40-3.
16. Abbott WS. A Method of computing the effectiveness of an insecticide. J Econ Entomol. 1925;18:265-7.
17. White T, Bruns T, Lee S, Taylor J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In: Innis M, Gelfand D, Sninsky J, White TJ, editors. PCR-Protocols and Applications---A Laboratory Manual. New York, NY: Academic Press; 1990. p. 315-22.
18. Sevim A, Demir I, Demirbag Z. Molecular characterization and virulence of Beauveria spp. from the pine processionary moth, Thaumetopoea pityocampa (Lepidoptera: Thaumetopoeidae). Mycopathologia. 2010;170(4):269-77.
19. Khosravi R, Sendi JJ, Zibaee A, Shokrgozar MA. Virulence of four Beauveria bassiana (Balsamo) (Asc., Hypocreales) isolates on rose sawfly, Arge rosae under laboratory condition. J King Saud Univ Sci. 2015;27:49-53.
20. Keller S, Schweizer C, Shah P. Differential susceptibility of two Melolontha populations to infections by the fungus Beauveria brongniartii. Biocontrol Sci Technol. 1999;9:441-6.
21. Arrieta F, Dukic V, Andreasen V, Dwyer G. Effects of host heterogeneity on pathogen diversity and evolution. Ecol Lett. 2015;8:1252-61.
22. Ekesi S, Adamu RS, Maniania NK. Ovicidal activity of entomopathogenic hyphomycetes to the legume pod borer, Maruca vitrata and the pod sucking bug, Clavigralla tomentosicollis. Crop Prot. 2002;21:589-95.
23. Zappelini L. Seleção deisolados dos fungos entomopatogênicos Beauveria bassiana e Metarhizium anisopliae visando o controle da broca da cana-de-açúcar, Diatraea saccharalis (Lepidoptera:Crambidae). São Paulo: Instituto Biológico, da Agência Paulista de Tecnologia dos Agronegócios; 2009.
24. Wenzel IM, Giometti FH., Almeida J. E. Patogenicidade doisolado IBCB 66 de Beauveria bassiana á broca da cana-de-açúcar Diatraea saccharalis em condições de laboratório. Arq Inst Biol. 2006;73:259-61.
25. Alves SB, Rossi LS, Lopes RB, Tamai MA, Pereira RM. Beauveria bassiana yeast phase on agar medium and its pathogenicity against Diatraea saccharalis (Lepidoptera:Crambidae) and Tetranychusurticae (Acari:Tetranychidae). J Invertebr Pathol. 2002;81:70-7.
26. Díaz B., Lecuona RE. Evaluación decepas nativas del hongo entomopatógeno Beauveria bassiana Bals. (Vuill.) (Deuteromicotina) como base para la selección de bioinsecticidas contra el barrenador Diatraea saccharalis (F.). Agriscientia.1995;XII:33-8.
27. Estrada ME, Romero M, Snowball M. Aplicación de Beauveria bassiana en la lucha biológica contra Diatraea saccharalis. Rev Caña Azúcar. 1997;15:39-43.
28. Legaspi JC, Poprawski TJ, Legaspi BC. Laboratory and field evaluation of Beauveria bassiana against sugarcane stalkborers (Lepidoptera: Pyralidae) in the Lower Rio Grande Valley of Texas. J Econ Entomol. 2000;93:54-9.
29. Potrich M, Alves LFA, Lozano E, Roman JC, Pietrowski V, Neves P. Interactions between Beauveria bassiana and Trichogramma pretiosum under laboratory conditions. Entomol Exp Appl. 2015;154:213-21.
30. Rossoni C, Kassab SO, Loureiro ED, Pereira FF, Costa DP, Barbosa RH, et al. Metarhizium anisopliae and Beauveria bassiana (Hypocreales: Clavicipitaceae) are compatible with Cotesia flavipes (Hymenoptera: Braconidae). Florida Entomol. 2014;97:1794-804.
Received in October, 2017.
Accepted in March, 2018.
Jennifer L García. Centro de Investigación Tibaitatá, Corporación Colombiana de Investigación Agropecuaria (AGROSAVIA) Km 14 vía Mosquera-Bogotá, Mosquera, Colombia. E-mail: jgarciar@agrosavia.co.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}