










 Servicios personalizados
Servicios personalizados texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkQuiroz et al. (2009) defined invasive species as "naturalized species that reproduce in large amounts and that have potential to spread in a considerable area, occupying natural habitats." These characteristics are present in Leucaena leucocephala, included in the list of the hundred invasive exotic species most harmful in the world, developed by the Unión Internacional para la Conservación de la Naturaleza (Lowe et al. 2004). However, it should be noted that not all exotic species become invasive. Their effect vary according to species and habitat. But, the invasive ones cause serious effects, with different consequences, especially if they are not controlled (Anon 2005). Nevertheless, internationally, there are many information sources that recommend this legume as an important tree in cattle rearing systems in different areas of the world. In Cuba, studies conducted with this plant have also confirmed this trait of leucaena. Studies of Ruiz et al. (2003) and Ruiz et al. (2015) demonstrated, integrally and in a multidisciplinary way, the potential of this plant for cattle rearing systems, besides proposing a technology. Therefore, it would be very interesting to study this possible contradiction and offer scientific results in this respect. The objective of this research was to evaluate the effect of management to prevent leucaena to become an invasive plant.
Materials and Methods
Experimental design and treatments. The evolution of two systems was evaluated: 1) tree legume Leucaena leucocephala cv. Peru in silvopastoral system in 100 % of the grazing area, associated with Brachiaria brizantha cv. Marandú for cattle fattening; 2) tree legume Leucaena leucocephala cv. Peru, where it occupied 100 % of the area, associated with Brachiaria brizantha vc. Marandú without being grazed. A random block design was applied, with five replications.
Experimental procedure. Leucaena was sown in double furrows, separated at 0.80 m from each other and spaced at 3.20 m. Brachiaria was planted on the same day of sowing the legume in three separate furrows at 1.0 m and a dose of 5 kg/ha. In the necessary cases, control of weeds was taken care of during the first 90 d.
The study was carried out on a physical surface of 50 ha on a wavy ferialitic soil (Hernández et al.2015). The land was divided into 40 paddocks of 1.25 ha, under dry conditions. Fattening cattle was used, with a stocking rate of 2 animals/ha, with an initial weight of 150 kg of LW and good physical condition and health. The occupation time was one and two days in rainy and dry season, respectively. Meanwhile, the rotation interval reached 40 and 80 d for rainy and dry period, respectively. The area that did not receive animal grazing occupied 6 ha and was protected with barbed wire fence. The studied areas were evaluated over five years (2011-2015). Samplings were carried out twice per season, in fixed areas of each paddock. The information offered is the mean per season. All samplings were considered and the first (beginning) and the last (end) were analyzed independently. The studied systems had eight years of establishment.
Samplings were systematically developed during dry and rainy season. The work was carried out on plots with a net area of 8.0 x 10.0 m. Performance of treatments was taken in 10 fixed frames of 1.0 m2. Height (grasses and weeds in cm and tree in m), yield of grass, weeds and leucaena (t/ha, DM), population (individuals/m2) and botanical composition of grass and weeds were measured, as well as the depopulation. These last two measures were transformed by √x and arc. sin √x, respectively. To determine the biomass production of the grass and weeds, it was cut at 15 cm height.
Statistical processing. The theoretical assumptions of the analysis of variance were verified for the counting variables, expressed in percent, from Shapiro and Wilk (1965) test for normality of errors and Levene test (1960) for homogeneity of variance. The analyzed variables fulfilled the theoretical assumptions of ANAVA. Analysis of variance was carried out according to design and Duncan (1955) test was applied when necessary. The statistical package INFOSTAT version 2012, prepared by Di Rienzo et al. (2012) was used.
Result and Discussion
All the indicators measured in the leucaena-brachiaria system were superior at the beginning and at the end of experiment (table 1). It should be noted that at that time the decrease in depopulation (P = 0.0012), the high grass population (P = 0.0014) and the low presence of weeds (P = 0.0039) were better with respect to the non-grazed system. This was characterized by low presence of grasses (P = 0.0041) and high levels of weeds (P = 0.0027). The same performance was presented at the beginning of the experiment, when comparing the grazed and non-grazed system.
Table 1 Indicators of grassland performance in different moments of experimental stage
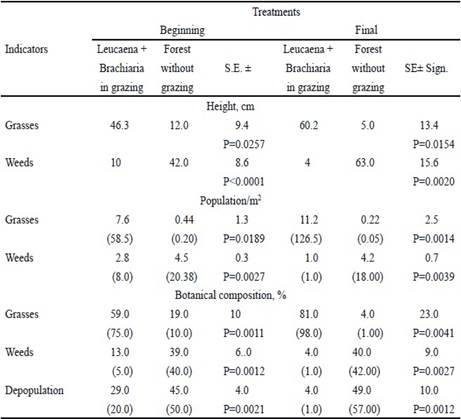
( ) Original values
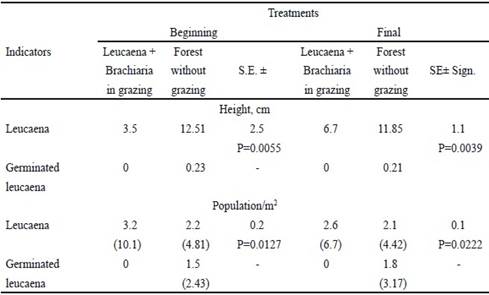
Leucaena component (table 2) evolved differently according to the studied system. In leucaena- brachiaria, planted area had a higher population at the beginning (P=0.0127) and at the end (P=0.0222) of the research, when compared with the system without grazing, which is normal in these systems. The increase in the plant height indicates that it is necessary to prune. In the area without grazing, these values were always higher than in the grazed system, when measured at the beginning (P = 0.0055) and at the end (P = 0.0039) of the study. There was only presence of new leucaena plants between rows of the non-grazed system. These plants were germinated from the seeds produced in the furrows planted with this legume. This clearly indicated that the non-management of leucaena areas caused the presence of new leucaena plants, which did not happen in the grazing system.
As it was expected, biomass production (table 3) had a similar performance to that indicated in table 1. In the system under grazing, brachiaria base grass yield was always superior to the unmanaged system, at the beginning (P=0.0028) and at the end of experimental stage. The same happened with the presence of natural grass (P=0.0011), which disappeared at the end. Weeds had a higher presence in the non-managed area (P=0.0101) at the beginning of the study. Leucaena maintained its production, according to Alonso (2004) and Ruiz et al. (2008). Total biomass production was always superior, at the beginning (P < 0.0001) and at the end (P < 0.0001) of the experiment for the grazed system. Disappearance of brachiaria and natural grass at the end of the experiment in the non-managed system could be caused by the excess of shade, which limited growth and stability of the herbaceous population. With respect to this topic, Ruiz et al. (2010) demonstrated the importance of management in order to achieve only the shade levels that contribute to the productive stability of silvopastoral systems. Biomass production for leucaena is not offered in the system without grazing due to the height reached by this plant (table 2).

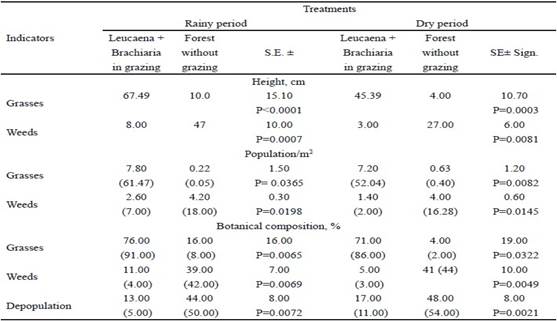
When analyzing the performance of these systems, according to the weather station, it is evident that in the leucaena-brachiaria grazing system, the evaluated indicators were always better than those found in the system that was not managed with animals (table 4). In general, the values of rainy season were better than in dry season. It should be noted that the presence of weeds was lower in leucaena-brachiaria, in rain as dry, with population (P = 0.0198 and P = 0.0145) and percent of weeds (P=0.0069 and P=0.0049), respectively. The same situation was presented for depopulation (P=0.0072 and P=0.0021). Pasture population was superior for this system (P=0.0365 and P=0.0082), as well as its presence in the botanical composition (P=0.0065 and P=0.0322), respecti- vely.
Leucaena component, very important in this study due to its characteristics of invader plant, showed higher values of population in leucaena-brachiaria system under grazing, after being compared with the one without grazing, during rainy (P=0.0041) and dry (P=0.0034) periods. This was not the same for plant germinated in the space between double furrows for the system without animals, and it shows the invasion of this tree when the system does not use animals properly (table 5).
Table 6 shows that, in leucaena-brachiaria system under grazing, components of biomass production were better than in the system without grazing, with brachiaria biomass production (P=0.0072) in rainy season and its disappearance in dry period in the area without management, and total production in rainy and dry periods (P=0.0622 and P=0.0062), respectively. Weeds showed lower values (P=0.0014 and P=0.0005) in rainy and dry seasons. This fact demonstrated that an improper management of areas provoked disappearance of each of the evaluated components, caused by the excessive growth of leucaena plants and their shade over the herbaceous stratum. This caused the decrease of sown grass growth, and the appearance and progressive development of weeds and new leucaena plants in non-sown areas. This last is a signal of invasion of this species when it does not receive an adequate management.


Rivera et al. (2011) and Paciullo et al. (2014) also reported this result with the silvopastoral system. These author referred a high biomass production during rainy periods and lower production in dry season. They attributed this performance to tree-grass interaction, which also favored animal production. They also pointed out that these systems reduce the decrease of production under unfavorable climate conditions.
A study of Mack et al. (2000) demonstrated that invasive plants may change ecological characteristics, like identity of dominant species in a community, physical properties of the ecosystem, nutrient cycle and plant productivity. In addition, it was confirmed that impact of biological invasions threatens the efforts for preserving biodiversity, maintaining productivity of agricultural systems, sustaining the functioning of natural ecosystems and protecting human health. All this could be appreciated in aspects mainly related to plant productivity and functioning of the system with and without grazing.
Studies developed in Cuba (Villate et al. 2016) with these species stated that if efficient strategies are not implemented for diminishing their impact, there will be the risk of becoming poor and homogenizing these ecosystems from which agriculture, cattle rearing, forestry, and fishing depend on, as well as other resources that provide irreplaceable natural services.
Despite the evident negative consequences of invasive species to native biological diversity, especially in islands, management of exotic species tend to have more into account the cost/benefit relation that implies its use, mainly in those that have important ecological and economical values. Under determined circumstances, the removal of an invasive species may cause bigger problems (Maron and Connors 1996 and Yelenik et al. 2004).
Sometimes, manageable invasive species may be used in activities of ecological and economical value. Under Cuban conditions, for example, it is in the reforestation of areas affected by mining, degraded grasslands and others, where exotic species, invasive in some cases, like L. leucocephala, favor recovery in front of the failure of repopulating with native species. This study also demonstrated that with a proper management of this species, some of the positive qualities of this plant can be used, which was also reported by Ruiz et al. (2005). Likewise, with the study of leucaena performance in different regions of Colombia, Calle et al. (2011) stated that this plant has been cultivated for the last 20 years, with the implementation of intensive silvopastoral systems with populations that surpass 20.000 plants per hectare, without the invasion of this plant in studied areas. In Australia, Shelton (2017) got to the same conclusion in a review of 70 years.
Therefore, it can be concluded that the technological proposal for managing silvopastoral systems with Leucaena leucocephala is a functional and feasible option to apply, so that this species does not become invasive.

