










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Desde tiempos inmemorables el ser humano ha modificado su entorno natural; entre otras cosas, para satisfacer las necesidades alimentarias de la población. Para lograrlo ha incurrido en la domesticación de un reducido número de especies vegetales, a partir de la reproducción selectiva de poblaciones con características deseables para el consumo. Con ello se han producido plantas diferentes a sus ancestros silvestres. Sin embargo, la domesticación de unas pocas especies ha puesto en evidencia la simplificación de los sistemas productivos (dada la existencia pocos cultivares) y el deterioro de los recursos fitogenéticos para la agricultura y la alimentación (RFAA).
Los RFAA se pueden definir como las plantas que han sido seleccionadas y conservadas por los agricultores de forma tradicional con valor potencial para la alimentación y la agricultura (AA), tanto en el presente como en el futuro (Eguiarte, et al., 2018). La diversidad de los recursos genéticos es la base de la AA, su conservación es esencial para su continuidad y al mismo tiempo provee de productos a la humanidad y proporciona seguridad alimentaria y nutricional. Las especies aprovechables están integradas por variedades tradicionales, modernas y parientes silvestres. Además, constituyen la materia prima a partir de la cual es posible obtener nuevas variedades necesarias para la alimentación de la población mundial (Gutiérrez, et al., 2019).
Es por ello que se hace necesario que los sistemas agrícolas incorporen lo aprendido sobre el funcionamiento de los ecosistemas naturales. La diversidad de especies, diversidad genética, redundancia y resiliencia son algunos ejemplos para enfatizar en la necesidad de una agricultura que garantice la seguridad alimentaria. A pesar de ello Gutiérrez, et al. (2019), señalaron que, en los últimos cincuenta años se ha modificado radicalmente la distribución y diversidad de especies destinadas a la alimentación. Como consecuencia se ha perdido más del 80 % debido a los procesos de desplazamiento de variedades tradicionales o criollas por mejoradas y a la eliminación de la población silvestre progenitora. Por otro lado, se ha observado que en los avances de la producción y tecnificación agrícola no se ha considerado la erosión de la diversidad biológica.
En el caso de Cuba, con el desarrollo del Programa Nacional de Agricultura Urbana (AU), Suburbana (ASU) y Familiar (AF), esta realidad se ha transformado. Según Companioni, et al. (2017), en el período 2017-2018 existían 23 subprogramas de los cuales 19 estaban presentes en la AU y la totalidad en la ASU. De estos subprogramas seis son agrícolas, cinco son de la parte pecuaria y 12 sirven de apoyo. Dentro de los subprogramas agrícolas se han popularizado tres de ellos ((i) hortalizas y condimentos frescos, (ii) granos, (iii) plátanos, raíces y tubérculos) todos relacionados con el grupo de plantas destinadas para la alimentación o que sirven de complemento a estas. El Ministerio de la Agricultura de Cuba (2018), señaló que con este programa se ha trabajado y se continúa trabajando en la consolidación de los enfoques agroecológicos y de sostenibilidad local que permiten incrementar la producción, la diversificación y la resiliencia general en la agricultura cubana. La diversificación de la producción, introduciendo y generalizando nuevos resultados, experiencias y cultivos ha sido otro de los aportes significativos.
La Organización de las Naciones Unidas para la Alimentación y la Agricultura (2018), señaló que para alimentar a los 10 mil millones de personas que representan la población estimada del planeta en 2050, se debe encontrar un equilibrio entre calidad y diversidad, vincular la productividad con la sostenibilidad y atender las necesidades de las personas. Lo imperativo de salvaguardar la biodiversidad y gestionar los recursos naturales (RN) de manera sostenible está cada vez más reconocido en los planes nacionales como un instrumento clave para ofrecer alimentos nutritivos a las generaciones presentes y futuras. Las técnicas que integran con éxito las dimensiones de la sostenibilidad tienen el potencial de conservar mejor los RN al tiempo que permiten cultivar más alimentos saludables con menos recursos y responden a una mejor nutrición y un consumo responsable.
De ahí la importancia de desarrollar estudios en los sistemas productivos de manera tal que permitan conocer cómo se comporta este componente y que los resultados obtenidos sirvan de base en el proceso de toma de decisiones en función de mantener la agrobiodiversidad dentro de los parámetros establecidos. En el caso específico de fincas suburbanas los estudios no resultan concluyentes sobre todo en Santiago de Cuba donde las investigaciones encaminadas en este sentido solo toman en cuenta algunos grupos funcionales dentro de la biodiversidad agrícola. Vargas, et al. (2016), realizan un análisis tomando en cuenta especies arvenses y arbustivas, por su parte Vargas, et al. (2017), centran su atención en grupos como las especies medicinales y ornamentales, en tanto, Vargas, et al. (2019a), consideraron las especies frutales. Lo planteado refuerza la teoría de que el componente vegetal (diferente a los frutales) que se destina para la alimentación dentro de la biodiversidad no ha sido suficientemente estudiado, a pesar de que se conoce que constituye la base para el logro de la seguridad alimentaria.
Teniendo en cuenta lo planteado anteriormente, el trabajo tuvo como objetivo determinar la composición, diversidad y distribución de las especies destinadas para la alimentación en fincas de la agricultura suburbana en Santiago de Cuba.
Materiales y métodos
La investigación se realizó en el Departamento de Agronomía de la Universidad de Oriente y en fincas de la Agricultura Suburbana del municipio Santiago de Cuba perteneciente a la provincia homónima. Se tuvieron en cuenta las dos etapas en las que se desarrolla la agricultura en Cuba (período lluvioso y poco lluvioso). El procedimiento empleado en cada fase de trabajo se describe a continuación.
El trabajo se llevó a cabo en las 10 fincas suburbanas que fueron seleccionadas por Guerrero (2017). En cada uno de los predios se procedió al conteo de las especies de plantas destinadas para la alimentación (PDPD). Para ello fueron establecidas parcelas como unidades de muestreo con dimensiones de 100 m2 (10 m x 10 m). Las parcelas fueron delimitadas por estacas de madera en sus cuatro ángulos y fueron trazadas en el sentido de las manecillas del reloj. Es válido destacar que el número de parcelas fue variable en dependencia del área de cada predio productivo y que en los dos períodos evaluados fueron utilizadas las mismas parcelas. Siempre se cuidó de que el área muestreada fuera al menos el 70 % del total. La información obtenida de los muestreos referente a cada una de las especies (nombre vulgar y número de individuos) en las parcelas levantadas fue recogida en hojas de campo datos que fueron utilizados en la identificación de las especies de plantas.
La identificación se realizó en el Departamento de Agronomía con la utilización del Diccionario Botánico de nombres Vulgares Cubanos según Roig (1988). Aquellas especies cuya identificación resultó difícil por las vías antes descritas fueron llevadas al Herbario del Centro Oriental de Ecosistemas y Biodiversidad (BIOECO) para ser identificadas por parte de los especialistas. En todos los casos los nombres científicos fueron cotejados con la obra de Acevedo & Strong (2012), y con el inventario preliminar de las Plantas Vasculares de Cuba de Greuter & Rankin (2017).
La determinación de la composición botánica se realizó teniendo en cuenta tres niveles fundamentales (Composición botánica general, Composición botánica por período y Composición botánica por finca en cada uno de los períodos). Se elaboraron listados de especies correspondientes a cada uno de los niveles de composición botánica establecidos. Los listados fueron organizados en orden alfabético partiendo de los nombres vulgares de las especies. Posteriormente se procedió a la cuantificación del total de individuos, número de familias, número de géneros y número de especies.
También se tuvo en cuenta las familias que más influyeron en el comportamiento de la composición botánica según los niveles establecidos. Se consideraron aquellas familias que aportaron como mínimo tres especies diferentes en cada una de las fincas aunque no necesariamente tenían que ser las mismas especies cuando se compara entre finca. Además, dicha familia debía tener una tendencia a la repetitividad considerando aquellas que se encontraron, de acuerdo con del Toro (2015), al menos en seis de las 10 fincas objeto de estudio a lo sumo en uno de los períodos evaluados.
Para la evaluación espacial y temporal del comportamiento de las PDPA se evaluaron indicadores de diversidad alfa (α) y diversidad beta (β). En el caso de la diversidad α se tuvo en cuenta el comportamiento de la Riqueza de especies (S), Dominancia de Berger-Parker (d) y Diversidad general (Shannon H´). Además de lo anterior se clasificaron las especies de acuerdo a la frecuencia de aparición. Para ello se procedió a calcular el porcentaje de abundancia (% Abd) y luego se cotejó con la escala utilizada por Vargas, et al. (2017). Se reportaron también las especies de mayor presencia en cada uno de los ecosistemas para ambos períodos de muestreo teniendo en cuenta las que presentaron mayor % Abd. Cada uno de los indicadores de diversidad α fueron determinados a partir del tabulador electrónico Biodiversity Calculator de Danoff-Burg & Chen (2005).
En el caso de la diversidad β se tuvo en cuenta el índice de similitud de Sorenson (Iss). El mismo fue determinado por medio del software estadístico SIMIL.exe. Una vez obtenidos los valores, para su interpretación se consideraron los rangos clasificatorios según la escala de RATLIFF referida por Rodríguez, et al. (2016). A partir de dicha escala fueron elaborados planos cartesianos con los diferentes rangos clasificatorios para la representación gráficas de los resultados. Es válido destacar que para el análisis de la diversidad β fue necesario el establecimiento de un total de 45 comparaciones entre las fincas suburbanas estudiadas.
Seguidamente se estableció para cada una de las PDPA la distribución de las mismas a partir de un análisis bibliográfico de la literatura especializada en el tema. Fueron utilizados como referencia documentos de estudios realizados por diferentes autores e instituciones de referencia nacional e internacional que han sido publicados en diversos sitios y bases de datos referenciadas preferentemente de los últimos 10 años según el procedimiento empleado por Guerrero (2017).
Resultados y discusión
En la investigación, se contabilizaron un total de 181 789 individuos de especies de PDPD o que sirven de complemento para esta, pertenecientes a 18 familias, 32 géneros y 43 especies. El comportamiento de cada categoría según período evaluado se muestra en la tabla 1.
Tabla 1 - Comportamiento de la composición botánica de las especies de plantas objeto de estudio.
| Especies | Categorías taxonómicas | |||||||
|---|---|---|---|---|---|---|---|---|
| Total de individuos | Familias | Géneros | Especies | |||||
| P. ll. | Ll. | P. ll. | Ll. | P. ll. | Ll. | P. ll. | Ll. | |
| PDPA | 120 596 | 61 193 | 18 | 18 | 33 | 32 | 42 | 40 |
Leyenda: PDPA: Plantas destinadas para la alimentación, P. ll.: Período poco lluvioso, Ll.: Período lluvioso
La composición botánica tuvo un comportamiento estable de un período al otro, con una ligera tendencia a la disminución para el total de individuos, géneros y especies, siendo el número de individuos el indicador que mostró la variación más marcada con más de un 49 %. Relacionado con el número de familias se puede decir que permanece constante con el cambio de época. De manera general este grupo de plantas mostró una buena composición botánica. La disminución del total de individuos puede que esté influenciada por el comportamiento de aquellos cultivos que encuentran su época óptima en el período poco lluvioso. Al llegar la lluvia determinados grupos de cultivos no permanecen en el predio productivo y en caso de permanecer lo hacen con un número de individuos muy reducido.
Por fincas (Tabla 2) la composición botánica tuvo un comportamiento entre la estabilidad y el aumento considerando los resultados de la cantidad de familia, géneros y especies. Sin embargo, el total de individuos tuvo una tendencia generalizada a la disminución con aumento solo en las fincas La Esperanza y El Sol. El número de especies mostró una disminución entre 1 y 9 en las fincas Tres Palmas, La Caballería, La Juliana y La Sorpresa. De forma general se evidencia que las fincas de un período al otro pueden sufrir un mayor grado de simplificación.
Tabla 2 - Composición botánica de las plantas destinadas para la alimentación en las fincas por períodos evaluados.
| Fincas | Categorías taxonómicas | |||||||
|---|---|---|---|---|---|---|---|---|
| Total de individuos | Familias | Géneros | Especies | |||||
| P. ll. | Ll. | P. ll. | Ll. | P. ll. | Ll. | P. ll. | Ll. | |
| Erick Vega | 2 858 | 2 855 | 8 | 8 | 10 | 12 | 11 | 14 |
| La Esperanza | 4 639 | 10 037 | 5 | 6 | 7 | 8 | 9 | 11 |
| Tres Palmas | 15 748 | 2 116 | 15 | 12 | 19 | 13 | 23 | 15 |
| La República | 10 399 | 6 222 | 13 | 13 | 16 | 18 | 19 | 20 |
| La Caballería | 3 270 | 1 386 | 12 | 11 | 16 | 14 | 19 | 16 |
| Los Cascabeles | 1 731 | 514 | 8 | 9 | 12 | 13 | 17 | 17 |
| La Juliana | 59 925 | 19 477 | 14 | 13 | 18 | 18 | 22 | 20 |
| El Sol | 6 203 | 12 647 | 10 | 12 | 17 | 18 | 20 | 22 |
| La Sorpresa | 1 942 | 1 337 | 13 | 11 | 15 | 13 | 17 | 14 |
| La Cecilia | 13 881 | 4 602 | 11 | 12 | 17 | 17 | 20 | 20 |
Leyenda: P. ll.: Período poco lluvioso, Ll: Período lluvioso
Las causas de la reducción del número de individuos coinciden con las descritas en los resultados de la composición botánica por período. Otras de las causas pueden ser el comportamiento de variabilidad de las especies. Puede darse el hecho de que en la época poco lluviosa aparezcan especies que luego no se encuentren en la lluviosa y que en esta temporada se introduzcan otras especies al sistema sin que ello implique necesariamente la incorporación de un número de individuos igual o mayor que los que resultan eliminados.
En la finca Tres Palmas de las 25 especies que se encontraron 13 son comunes a ambos períodos. Ajo puerro (Allium ampeloprasum L.), berenjena (Solanum melongena L.), cebolla (Allium cepa L.), col (Brassica oleracea L.), culantro (Eryngium foetidum L.), malanga (Xanthosoma sagittifolium (L.) Schott.), perejil (Petroselinum crispum (Mill.) Nymam ex. A. w. Hill.), pimiento (Capsicum annuum L.), sorgo (Sorghum bicolor (L.) Moench.) y tomate (Solanum lycopersicum L.) solo se reportaron en el período poco lluvioso. El frijol gandul (Cajanus cajan (L.) Huth.) y el quimbombó (Abelmoschus esculentus (L.) Moench.) se encontraron únicamente en la época lluviosa. De las 19 especies que se contabilizaron en La Caballería, 15 se reportaron en ambas temporadas. Ñame (Dioscorea alata L.), oreganito (Lippia sp.), habichuela (Vigna unguiculata (L.) Walp.) y S. lycopersicum solo aparecieron en el período poco lluvioso.
En La Juliana se reportaron 25 especies vegetales de las cuales 17 son compartidas por ambos períodos. Ají cachucha (Capsicum baccatum L.), café (Coffea arabica L.), cebolla multiplicadora (Allium cepa var. Aggregatum Dom.), B. oleracea y C. cajan se listaron solo en la temporada poco lluviosa. Espinaca (Spinacia oleracea L.), girasol (Helianthus annuus L.) y maíz (Zea mays L.) le correspondieron únicamente al período lluvioso. La Sorpresa fue una finca donde se contabilizaron 23 especies de las cuales 8 son iguales para los dos momentos evaluados. De las que no son comunes, C. baccatum, S. melongena, E. foetidum, king-grass (Cenchrus purpureus (Schumanch.) Morrore), lechuga (Lactuca sativa L.), Lippia sp., plátano macho (Musa paradisiaca L.), S. lycopersicum y zanahoria (Daucus carota L.) se presentaron únicamente en la época poco lluviosa. En la temporada lluviosa aparecieron caña de azúcar (Saccharum officinarum L.), S. oleracea, plátano burro (Musa sp.), A. esculentus, rábano blanco (Raphanus sativus L.) y remolacha (Beta vulgaris L.).
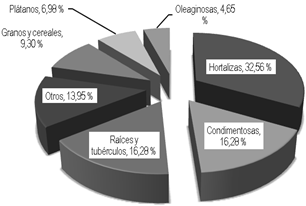
De forma general se identificaron siete grupos de cultivos dentro de las PDPA (Fig. 1). El más representado fue el de las hortalizas con un 32,56 % y le siguieron con igual porcentaje (16,28 %) las condimentosas, así como, las raíces y los tubérculos. Con menor representatividad estuvieron los granos y cereales, los plátanos y las oleaginosas. Un hecho interesante es que el 13,95 % está representado por otros cultivos como el C. arabica, S. officinarum, C. purpureus, morera (Morus alba L.), S. bicolor y tabaco (Nicotiana tabacum L.) que, si bien no forman parte de la dieta de los seres humanos, estos de manera indirecta si tributan al logro de una alimentación adecuada. En las condiciones estudiadas S. officinarum, C. purpureus y M. alba contribuyen a la alimentación animal, al igual que S. bicolor que demás es utilizado en el control de plagas como barrera proteccionista. Finalmente N. tabacum en condiciones de cultivo pudiera contribuir a generar ingresos que una vez obtenidos se pondrían en función de la producción de alimentos, pero teniendo en cuenta el reducido número de individuos que se reportó para esta especie (3), es posible que su uso esté más relacionado con la obtención de productos naturales para el control de plagas, con lo cual contribuiría a crear condiciones óptimas para una mayor producción de aquellos cultivos que si contribuyen directamente a la alimentación.
Todos los resultados expuestos hasta aquí explican, por un lado, la reducción que se experimentó en el número de especies y con ello en el número de individuos. En el grupo de plantas objeto de estudio, las especies que son propias o que aparecen en cada uno de los períodos evaluados, existe una amplia representación de aquellas que se consideran cultivos hortícolas. De ello se infiere que este tipo de cultivos si bien constituyen importantes elementos de diversificación, también es cierto que cuando concluye su período óptimo tributan a la simplificación de los ecosistemas agrícolas. Por otro lado, se evidencia que las necesidades alimenticias que más se satisfacen desde los predios productivos estudiados son las relacionadas con la disponibilidad de hortalizas, condimentos frescos y las viandas (fundamentalmente raíces y tubérculos).
Del total de familias reportadas solo el 50 % (Tabla 3) son las que mayor influencia tienen en el comportamiento de la composición botánica. Leguminosae (Fabaceae) y Musaceae tienen buena representatividad con al menos una especie muestreada en ambos períodos en el 100 % de los predios productivos objeto de estudio. Por su parte las familias Amaranthaceae y Cucurbitaceae ganan presencia en +1 predio productivo de una temporada a la otra, aumentando del 50 al 60 % para el caso de la primera y en el caso de la segunda del 80 al 90 %. En el resto de las familias se experimenta una reducción de su presencia en -1 finca de un período al otro, excepto en la familia Solanaceae, donde la reducción en los sistemas productivos donde se encuentra es de -3. Lo planteado demuestra que, con la llegada de la lluvia, existe una pérdida de estas familias en el 10 y 30 % de los predios productivos objeto de estudio. A pesar de todo lo planteado anteriormente se considera que las nueve familias listadas como las de mayor contribución tienen buena representatividad en todas las fincas suburbanas estudiadas.
Tabla 3 - Familias botánicas de mayor contribución al comportamiento de la composición botánica en cada uno de los grupos.
| Familias | Fincas donde aparecen | |
|---|---|---|
| Período poco lluviosos | Período lluvioso | |
| 3,4,6,9,10 | 3,4,6,7,9,10 | |
| 3,4,5,6,7,8,9,10 | 4,5,6,7,8,9,10 | |
| 3,4,5,6,7,9,10 | 3,4,5,6,7,10 | |
| 3,4,5,7,8,9,10 | 3,4,5,7,8,10 | |
| 1,3,4,5,6,7,8,9 | 1,3,4,5,6,7,8,9,10 | |
|
|
1,2,3,4,5,6,7,8,9,10 | 1,2,3,4,5,6,7,8,9,10 |
| 1,2,3,4,5,6,7,8,9,10 | 1,2,3,4,5,6,7,8,9,10 | |
| 1,2,3,4,5,7,8,9,10 | 1,2,3,4,5,7,8,9 | |
| 1,2,3,4,5,6,7,8,9,10 | 2,4,5,6,7,8,10 | |
Leyenda: 1: Erick Vega, 2: La Esperanza, 3: Tres Palmas, 4: La República, 5: La Caballería, 6: Los Cascabeles, 7: La Juliana, 8: El Sol, 9: La Sorpresa, 10: La Cecilia
El que estas familias tengan buena representatividad se debe a que en ellas se encuentran gran número de especies cultivadas que comúnmente se encuentran en los sistemas productivos al estar ampliamente difundidos y ser demandados por la población. El que Leguminosae sea una familia con contribución importante puede estar relacionado con que desde la nueva clasificación taxonómica esta familia quedó integrada por las antiguas familias Mimosaceae, Caesalpinaceae y Fabaceae dentro de las cuales se incluyen importantes cultivos agrícolas. Algo similar sucede con las familias Amaranthaceae, Amaryllidaceae y Apiaceae que incluyen un grupo de familias que fueron renombradas y reagrupadas.
Los resultados obtenidos en este trabajo son superiores al 66,67 % obtenido por Siret (2018), en sistemas productivos del tipo familiar. Este autor reportó a Musaceae dentro de las familias de mayor contribución a la composición botánica y a Asteraceae dentro de las de mayor distribución. Rodríguez (2018), al analizar comportamiento de especies vegetales y criterios sociales que sustentan su presencia en la agricultura familiar en Santiago de Cuba señaló dentro de las familias más contribuyentes, además de las anteriores, a Leguminosae, Euphorbiaceae, Malvaceae y Araceae. En tanto Fernández (2019), al determinar las potencialidades de la flora existente en patios familiares para generar bienes y servicios en la provincia antes mencionada, al listado de las familias anteriores, agregó la familia Solanaceae.
La riqueza de especies de una temporada a la otra dentro de una misma finca fue estable, aunque es válido destacar que dentro de la estabilidad hubo una tendencia al aumento en algunos predios productivos. Se manifestó un aumento en el 40 % de los sistemas productivos y en igual porcentaje de estos se produjo una disminución, en el 20 % restante la riqueza de especies se mantuvo constante. En los predios donde se manifestó una disminución no fue muy marcada, solo en la finca Tres Palmas hubo una pérdida de ocho especies siendo este predio productivo donde se reportó el mayor valor de riqueza para el período poco lluvioso. En la época lluviosa el mayor valor se alcanzó en la finca El Sol. De forma general en todos los predios productivos se asegura la presencia de plantas cuya finalidad es la producción de alimentos o como complemento de esta.
El comportamiento observado para este grupo de plantas está fuertemente influenciado por los fines productivos que se desarrollan en las fincas. Otro de los factores puede estar relacionado con la disponibilidad de algunos recursos sobre todo semillas de las cuales depende la presencia o ausencia de una determinada especie en un ecosistema agrícola. El cambio de temporada también tuvo influencia pues los cultivos se desarrollan en espacios de tiempos que se consideran períodos óptimos los cuales cambian conforme las condiciones y con ello varía tanto la presencia como la cantidad de una determinada especie.
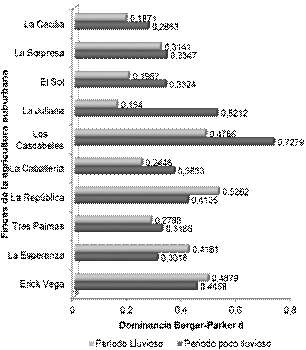
Fig. 2 - Dominancia de las plantas destinadas para la alimentación en las fincas suburbanas objeto de estudio.
En cuanto a la dominancia (Fig. 2) se puede observar que existe una disminución del valor de este indicador en el 70 % de las fincas suburbanas estudiadas. La pérdida de importancia de las especies más abundantes tiene fuerte implicación en el comportamiento de estos resultados, lo que también influyó en el comportamiento del resto de las especies dentro de la muestra. Aunque todos los valores de dominancia obtenidos se encuentran dentro del rango establecido para este indicador, el mayor valor para el período poco lluvioso se reportó en la finca Los Cascabeles (0,7279) y en La República (0,5262) para la temporada lluviosa.
De acuerdo con lo referido por Vargas, et al. (2019a), la dominancia de Berger-Parker (d) es un indicador que debe asumir valores de 0 a 1. De lo planteado se entiende que en la misma medida que los resultados obtenidos se acerquen al límite superior es indicativo de mayor dominancia y por tanto, menor diversidad y equidad. Agregaron además que este indicador considera la dominancia del taxón o la especie más abundante. Teniendo en cuenta lo antes señalado se puede decir que en las fincas donde se experimenten los mayores valores en uno u otro período, son las áreas donde menor diversidad para este grupo de plantas se debe manifestar y por tanto la distribución de los individuos dentro de la muestra, supone debe ser menos equitativa. En el caso de aquellos predios donde se manifiesten los valores más bajos, será indicativo de que existe una mayor diversidad por lo cual la distribución de los individuos dentro de la muestra es más equilibrada y existe una menor influencia del taxón más dominante.
Las especies más dominantes (Tabla 4) tienden a disminuir su porcentaje de abundancia los que se relaciona con el comportamiento mostrado por el indicador de dominancia evaluado. Dentro del grupo de plantas objeto de estudio las especies más abundantes se mantienen de un período al otro solo en el 40 % de los predios productivos (Erick Vega, Tres Palmas, La República, Los Cascabeles). En el resto de los sistemas agrícolas estudiados (60 %) las especies más dominantes cambian con el cambio de época. Por otra parte, el 45,45 % de las especies más dominante (Yuca (Manihot esculenta Crantz.), S. officinarum, L. sativa, A. ampeloprasum, Musa sp.) son comunes a las dos épocas estudiadas.
Del total de especies identificadas, el 25,58 % (11 especies), se encontraron dentro de las más dominantes. De esas 11 especies, el 81,82 % (nueve especies) se reportaron en la temporada poco lluviosa y sietes especies que representan el 63,64 % en el período lluvioso. D. carota y P. crispum solo aparecen dentro de las especies más dominante en el período poco lluvioso, lo que puede estar relacionado con el hecho de que son cultivos hortícolas y su ciclo de cultivo pudiera estar relacionado con la escases de lluvia. Por otra parte, V. unguiculata y H. annuus son especies dominantes solo en la época lluviosa. En el período poco lluvioso las especies más dominantes fueron clasificadas como ocasionales en el 60 % de los sistemas productivos, poco frecuente en el 30 % y frecuente en el 10 %. En la temporada lluviosa las taxas de mayor contribución fueron escasas en el 30 % de las fincas y en igual porcentaje de predios productivos al referido anteriormente se clasificaron como ocasionales, en tanto en el 40 % restante fueron consideradas poco frecuentes.
Tabla 4 - Comportamiento de las especies más abundantes en el grupo de plantas objeto de estudio.
| Fincas | Período poco lluvioso | Período lluvioso | ||||
|---|---|---|---|---|---|---|
| Especies | % Abd | Clasif. | Especies | % Abd | Clasif. | |
| 1 | 48,79 | Poco frecuente | 48,79 | Poco frecuente | ||
| 2 | 30,18 | Ocasional | 41,61 | Poco frecuente | ||
| 3 | 31,86 | Ocasional | 27,88 | Ocasional | ||
| 4 | 41,35 | Poco frecuente | 52,62 | Poco frecuente | ||
| 5 | 36,33 | Ocasional | 24,46 | Ocasional | ||
| 6 |
|
72,79 | Frecuente |
|
47,86 | Poco frecuente |
| 7 |
|
52,12 | Poco frecuente | 15,40 | Escasa | |
| 8 | 33,24 | Ocasional | 19,67 | Escasa | ||
| 9 | 33,47 | Ocasional | 31,41 | Ocasional | ||
| 10 | 26,83 | Ocasional |
|
18,71 | Escasa | |
Leyenda: 1: Erick Vega, 2: La Esperanza, 3: Tres Palmas, 4: La República, 5: La Caballería, 6: Los Cascabeles, 7: La Juliana, 8: El Sol, 9: La Sorpresa, 10: La Cecilia, % Abd: Porcentaje de abundancia, Clasif.: Clasificación
También se puede apreciar que en la temporada de poca lluvia, del total de fincas, las especies identificadas dentro de las más dominantes, en el 60 % de ellas, se incluyen en los taxas con menor implicación dentro de la muestra al estar en límite inferior de la escala de clasificación (escasa-ocasional). El resto de las especies se encuentran en el lado centro superior de la escala (poco frecuente-frecuente-abundante). A. ampeloprasum es una especie que estuvo presente en dos (20 %) de las 10 fincas objeto de estudio. Con la llegada de la lluvia se mantuvo constante el porcentaje de fincas (60 %) con especies cuya clasificación se ubica en el límite inferior de la escala, aunque hay que destacar que en el 30 % de ellas, para esta época se incluyen especies escasas. En este período A. ampeloprasum se reportó en tres (30 %) de las fincas estudiadas y Musa sp. en dos (20 %) de los sistemas productivos.
Excepto S. officinarum y H. annuus (ornamental), el resto de las especies dentro de las más dominantes son cultivos que se consumen como hortalizas o viandas lo que ratifica que las necesidades alimenticias relacionadas como estos grupos de cultivos son las que buscan satisfacer desde el contexto más cercano. También es posible que el objeto de productivo de estas fincas tenga influencia en la predominancia de estos cultivos pues, excepto en Los Cascabeles que de acuerdo con Vargas, et al. (2016), el mango (Mangifera indica L.) es su cultivo principal y La Caballería que según Vargas, et al. (2019b), se dedica fundamentalmente al cultivo de plantas ornamentales, en el resto de las fincas se dedican a los cultivos varios.

Fig. 3 - Diversidad de las plantas destinadas para la alimentación en las fincas suburbanas objeto de estudio.
La diversidad general (Fig. 3) tuvo un comportamiento variable aumentando su valor de un período al otro en el 70 % de los predios productivos estudiados. Los mayores valores para este indicador en ambos períodos se reportaron en La Cecilia con 2,2221 y 2,2565 respectivamente. Los valores más bajos se obtuvieron en La Juliana (1,0701) y La Esperanza (1,5885) para el período poco lluvioso y lluvioso en cada caso. A pesar de lo planteado todos los valores obtenidos se encuentran en el rango establecido para una correcta diversidad y abundancia (de 1 a 5).
Vargas, et al. (2017), cuando estudiaron la diversidad de algunas de las fincas incluidas en el presente estudio considerando las especies alimenticias, ornamentales y medicinales como grupo de plantas, obtuvieron valores de diversidad más variables que los reportados en la presente investigación, inclusive, hubo valores inferiores. El que existan estos valores de diversidad en ambos períodos es importante. En primer lugar es muestra de que estas fincas garantizan una presencia variada de elementos nutritivos para el contexto social más cercano. En segundo lugar cumplen su papel en logro de la seguridad alimentaria en condiciones de un desarrollo local sostenible, que es uno de los principios del Programa Nacional de AU, ASU y AF desde su comienzo hasta la actualidad.
La similitud entre estas fincas, considerando las PDPA o que sirven de complemento a esta, manifiesta que el valor del Iss, considerando todas las comparaciones, se encontró entre 0,000 y 0,500 en ambos períodos (Fig. 4). Específicamente en la temporada poco lluviosa (Fig. 4A) de las 45 comparaciones realizadas el 8,89 % (las comparaciones La Esperanza-Los Cascabeles, Tres Palmas-La República, Tres Palmas-La Cecilia, La República-La Caballería) presenta una similitud moderada. Las 41 comparaciones restantes (el 91,11 %) se concentran en la baja similitud. De las 41 comparaciones que se encuentran en la baja similitud, el 53,66 % se ubica en el cuadrante de la disimilitud.
En la época lluviosa (Fig. 4B) se mantiene el que no existen comparaciones que alcanzan valores altos de similitud (por encima de 0,5000), aunque, se manifestó un incremento (17,78 %) de aquellas que alcanzan una similitud moderada (Erick Vega-La Esperanza, Tres Palmas-La Cecilia, La República-La Juliana, La República-La Cecilia, La Caballería-Los Cascabeles, La Caballería-La Sorpresa, Los Cascabeles-La Sorpresa, La Juliana-El Sol). De ellas solo la comparación 24 (Tres Palmas-La Cecilia) se mantiene con el mismo comportamiento de un período al otro, el resto fueron comparaciones que en la temporada anterior alcanzaron baja similitud. En el caso de las comparaciones con baja similitud se experimentó una disminución pues en este período solo 37 de ellas, que representan el 82,22 %, mantienen esta condición, de las cuales 16 (43,24 %) se ubican en el cuadrante de la disimilitud, resultado que también es inferior al de la temporada anterior.
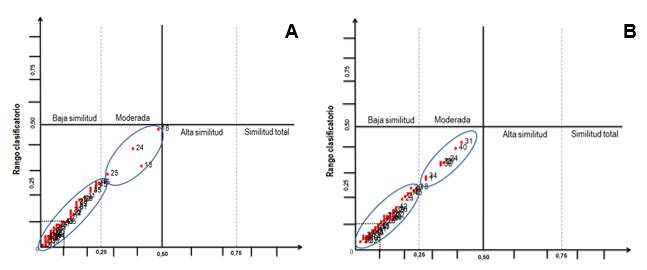
Fig. 4 - Comportamiento de la similitud según el índice de Sorenson (Iss) en el período poco lluvioso (A) y período lluvioso (B).
A pesar del comportamiento descrito anteriormente todos los valores obtenidos están dentro del rango establecido para este indicador (0-1). De forma general los bajos valores de similitud que se manifiestan entre estas fincas teniendo en cuenta el grupo de PDPA (anexo 1), están asociados en primer lugar a la existencia de pocas especies comunes entre los sistemas que se comparan. En segundo lugar, la existencia de especies comunes con una diferencia muy elevada entre ellas en cuanto al número de individuos, también influyó en los bajos valores obtenidos.
Estos resultados para nada se consideran negativos y si hablan, quizás, de la presencia de un número de especies en estas fincas que siempre que se cultiven se adaptan muy bien a las condiciones edafoclimáticas de cada una de ellas. Por otra parte, la influencia del contexto social pudo tener repercusión, pues como sistemas productivos a pequeña escala que son manejados por la familia fundamentalmente, siempre aspectos subjetivos como los gustos a las preferencias van a influir en lo que se cultive, siendo este el primer paso para satisfacer necesidades. Aparejado a lo señalado anteriormente, el aspecto económico tiene una influencia importante, pues una vez que las necesidades de quienes viven y trabajan en las fincas quedan satisfechas, se trata producir lo que mejor se venda.
Estudios como los desarrollados por Guerrero (2017); Rodríguez (2018); Siret (2018); Fernández (2019), han señalado la existencia de similitud baja o moderada al comparar sistemas productivos a pequeña escala (fincas suburbanas, patios familiares y ecosistemas cafetaleros) en las condiciones de Santiago de Cuba. En todos los casos han señalado entre las principales causas la existencia de pocas especies comunes entre las áreas comparadas, así como, las tradiciones de las personas que viven y trabajan en los predios productivos. Los resultados del presente estudio considerando las comparaciones ubicadas en la similitud alta o moderada, son inferiores a los reportados por Vargas, et al. (2019a), para estas mismas fincas, pero considerando las especies frutales. Sin embargo, fueron superiores cuando se tienen en cuenta las comparaciones ubicadas en la baja similitud. Estos autores señalaron que el indicador de similitud utilizado tiene en cuenta el número de especies comunes entre las entidades que se comparan y el total de individuos dentro de dichas especies. Según aumente la cantidad de especies comunes y el número de individuos dentro de ellas no difiera en exceso, así aumentará la similitud de las muestras de menor riqueza de especies, frente a aquellas que tienen mayor riqueza.
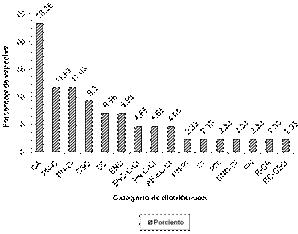 Leyenda: CA: Cultivada ampliamente, PN-C: Posiblemente naturalizada, cultivada en Cuba, PN-CI: Posiblemente naturalizada, considerada invasora, CSO: Cultivado solo ocasionalmente, CC: Cultivada en Cuba, ENC: Exótica o naturalizada en Cuba, ENC-C-CI: Exótica o naturalizada en Cuba, cultivada, considerada invasora, PN-C-CI: Posiblemente naturalizada, cultivada, considerada invasora, AF-EC-CI: Adventicia frecuente, exótica en Cuba, considerada invasora, PN-PI: Posiblemente naturalizada, potencialmente invasora, CI: Considerada invasora, PDI: Presente pero dudosamente indígena, ENC-CI: Exótica o naturalizada en Cuba, considerada invasora, PN: Posiblemente naturalizada, E-CA: Exótica o cultivada ampliamente, EC-CSO: Exótica en Cuba, cultivado solo ocasionalmente.
Leyenda: CA: Cultivada ampliamente, PN-C: Posiblemente naturalizada, cultivada en Cuba, PN-CI: Posiblemente naturalizada, considerada invasora, CSO: Cultivado solo ocasionalmente, CC: Cultivada en Cuba, ENC: Exótica o naturalizada en Cuba, ENC-C-CI: Exótica o naturalizada en Cuba, cultivada, considerada invasora, PN-C-CI: Posiblemente naturalizada, cultivada, considerada invasora, AF-EC-CI: Adventicia frecuente, exótica en Cuba, considerada invasora, PN-PI: Posiblemente naturalizada, potencialmente invasora, CI: Considerada invasora, PDI: Presente pero dudosamente indígena, ENC-CI: Exótica o naturalizada en Cuba, considerada invasora, PN: Posiblemente naturalizada, E-CA: Exótica o cultivada ampliamente, EC-CSO: Exótica en Cuba, cultivado solo ocasionalmente.Fig. 5 - Representatividad de los rangos de distribución de las especies destinadas para la alimentación.
Se identificaron 16 rangos de distribución considerando el total de PDPA o que sirven como complemento de esta en las fincas estudiadas (Figura 5). Las cultivadas ampliamente en nuestro país fueron las que alcanzaron mayor porcentaje (23,26 %) seguidas con igual porcentaje (11,63 %) de las posiblemente naturalizadas que son cultivadas y de las posiblemente naturalizadas que se consideran invasoras. En tercer lugar, con un 9,3 % se encontraron las especies que solo se cultivan ocasionalmente.
El resto de los rangos de distribución estuvo representado entre el 2,33 y el 6,98 %. Un hecho interesante es que del total de rangos encontrados el 43,75 % está relacionado con especies que, a pesar de ser cultivadas en los predios productivos estudiados, tienen un potencial invasivo. Dentro de estas especies se encuentran: C. baccatum, ají guaguao (Capsicum frutescens L.), calabaza (Cucurbita pepo L.), frijol caballero (Phaseolus lunatus L.), C. cajan, maní (Arachis hypogaea L.), papasán (Coccinia grandis (L.) Voigt.), pepino (Cucumis sativus L.), C. annuum, A. esculentus, R. sativus, N. tabacum, S. lycopersicum, M. esculenta.
Lo descrito anteriormente explica por un lado el por qué muchas de estas especies fueron encontradas en varios de los predios productivos estudiados y, por otro lado, la razón de que otras se encuentren entre las especies más dominantes, inclusive en los dos períodos evaluados. Ello hace suponer que el hombre de forma tradicional ha utilizado este potencial de manera controlada para domesticar muchas de estas especies que hoy conocemos en su forma cultivada. Estudios como el realizado por Gutiérrez, et al. (2019), al analizar la diversidad agrícola y seguridad alimentaria y nutricional en dos localidades Mayas de Yucatán, señalaron la presencia de especies con potencial invasivo como P. lunatus, M. esculenta, S. lycopersicum y diferentes especies del género Capsicum dentro de las más cultivadas. De forma general las PDPA tienen amplia distribución en todo el mundo, aunque buena parte de ellas tienen en la región de Centroamérica y América Latina su centro de origen y distribución.
Conclusiones
La composición de las especies destinadas para la alimentación tuvo un comportamiento estable de un período al otro manifestando una ligera tendencia a la disminución en el total de género y especies, excepto en el total de individuos que la disminución fue más marcada, por fincas, se experimentó un aumento con la estabilidad en algunas de ellas.
Las hortalizas (32,56 %), las condimentosas (16,28 %), las raíces y los tubérculos (16,28 %) fueron los grupos más representados. La diversidad α mostró un aumento con la llegada de la lluvia en el 70 % de los sistemas estudiados, sin embargo, predominaron las especies clasificadas como escasas dentro de cada una de las muestras, en tanto, la diversidad β arrojó un predominio de la baja similitud entre las comparaciones realizadas con más de un 43 % de ellas en el área de la disimilitud.
Este grupo de plantas en las fincas está representado en su mayoría por especies que son cultivadas ampliamente en Cuba, sin embargo, varios de los rangos de distribución identificados incluyen especies con potencial invasivo.

