










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
En el marco de los sistemas alternativos a la avicultura intensiva, la implementación de estrategias productivas con aves mantenidas en semicautiverio en condiciones más afines con el comportamiento natural de la especie requiere de genotipos adaptados. Para este tipo de producción son adecuadas poblaciones doble-propósito habituales antes de la especialización en carne y huevo propia de la avicultura industrial actual. La utilización de este tipo de aves evita el sacrificio de todos los machos de un día de edad provenientes de las líneas especializadas en producción de huevos, práctica que colisiona tanto con la filosofía que sustenta la producción orgánica como con principios éticos en un mundo en el que millones de personas padecen de hambre crónica o directamente mueren por falta de alimentos (Leenstra et al., 2011; Krautwald-Junghanns et al., 2018; Busse et al., 2019).
Si no son aptas para la producción intensiva (Leenstra, Horne y Krimpen, 2010; Damme, 2015), sí lo son para aquellos productores capaces de utilizar un nicho particular del mercado asociado a la demanda de productos generados en sistemas más amigables tanto con el animal como con el ambiente (Hammershøj, Kristiansen y Steenfeldt; 2021). Su producción se vincula también con poblaciones vulnerables en términos de seguridad alimentaria y con una mayor productividad de la avicultura de traspatio basada, por lo general, en la cría de aves criollas de gran rusticidad, pero de limitada productividad (Mack, Hoffmann y Otte, 2005; Alders y Pym, 2009). Campero Casilda es un cruzamiento de tres vías propuesto como ave doble propósito para sistemas avícolas alternativos al industrial, que preservan el bienestar animal, que ha sido evaluado satisfactoriamente para la producción de carne (Canet et al., 2021). La utilización de las hembras como ponedoras, y eventualmente para consumo como gallinas de descarte, enfrenta el desafío de la incompatibilidad genética entre velocidad de crecimiento -asociada a la producción de carne- y los caracteres reproductivos asociados con la oviposición (Robinson et al., 1993; Barbato, 1999).
Si bien presenta una adecuada curva de postura (Romera et al., 2020), por su condición de aves pesadas su caracterización como ponedora requiere estudiar las particularidades de sus patrones de crecimiento y aumento de peso del huevo. El objetivo de este trabajo fue evaluar dichos patrones dinámicos en comparación con los de dos genotipos de reproductoras pesadas comerciales.
MATERIALES Y MÉTODOS
Aves
Se evaluaron gallinas Campero Casilda, cruzamiento de tres vías entre gallos de la población sintética AH’ (50% Hubbard 50% estirpe Anak grises) y gallinas derivadas del cruzamiento simple entre machos de la sintética ES (87,5% Cornish Colorado 12,5% Rhode Island Red) y hembras de la sintética A (75% Cornish Colorado 25% Rhode Island Red). Las poblaciones sintéticas mencionadas se generaron y mantienen en la Sección Avicultura de la Estación Experimental Agropecuaria “Ing. Agr. Walter Kugler” del Instituto Nacional de Tecnología Agropecuaria (INTA), Argentina, en cuyas instalaciones se llevaron a cabo los cruzamientos mencionados y las incubaciones. Al nacimiento, las aves se vacunaron contra la enfermedad de Marek, se identificaron con banda alar y se trasladaron a la Facultad de Ciencias Veterinarias de la Universidad Nacional de Rosario. Todas las aves recibieron el plan sanitario y se mantuvieron bajo las condiciones establecidas en el protocolo de producción de pollos camperos (Bonino, 1997).
Entre el nacimiento y los 35 días de edad, se criaron a piso, con una densidad inicial de 25 aves/m2 hasta los 15 días, la que se redujo en ese momento a 10 aves/m2, un fotoperíodo decreciente con un ciclo mínimo de oscuridad de 8 horas y libre acceso al agua y al alimento. En la 5a semana de vida 100 pollas elegidas al azar se alojaron en jaulas individuales de postura (30 cm de frente x 42 cm de altura x 50 cm de profundidad). Las aves recibieron alimento balanceado Iniciador (18 % PB; 2.875 kcal EM) entre el nacimiento y las 8 semanas; Crecimiento (15,4 % PB; 2.881 kcal EM) entre las semanas 9 y 16; Prepostura (16 % PB; 2.833 kcal EM) entre las semanas 17 y 22 y Postura (15,6 % PB; 2.824 kcal EM) a partir de las 22 semanas. Dada su condición de aves pesadas, a partir de la semana 6 se les restringió el consumo de alimento, de acuerdo al protocolo de manejo de aves reproductoras camperas, en función de su peso corporal hasta el inicio de la postura y en función de su peso corporal y su producción de huevos a partir de ese momento y hasta la finalización del ciclo.
Método
Para todas las aves se registró el peso corporal individual a intervalos semanales entre el nacimiento y las 60 semanas de edad y el peso de todos los huevos puestos entre la madurez sexual y la finalización del primer ciclo de postura.
Los datos individuales peso corporal (g) - edad cronológica (semanas) se ajustaron con el modelo sigmoideo de Gompertz (Fitzhugh, 1976): Wp(t)= Ap exp (-b exp (-k*t)), dónde: Wp(t) = peso corporal en el tiempo t, Ap = peso corporal asintótico (valor de Wt cuando t → ∞, b = constante de integración parámetro de posición sin valor biológico, k = tasa de maduración (velocidad de aproximación a A) y t = edad cronológica (semanas).
Peso promedio semanal del huevo (g) - edad de postura (semanas) de cada ave se ajustaron con el modelo exponencial asintótico de Weatherup y Foster (1980): Wh(t) = Ah - B * (r^t), donde Wh(t) = peso del huevo (g) en el tiempo t, Ah = peso asintótico del huevo, B = rango de peso del huevo entre el inicio de la postura y la asíntota, (A-B) = peso teórico del primer huevo, r = tasa de maduración para peso del huevo, velocidad de aproximación al valor de A (0≤ r ≤ 1), y t = tiempo en semanas de postura.
En ambos casos los ajustes se llevaron a cabo por regresión no lineal utilizando una técnica iterativa basada en el algoritmo de Marquardt (Marquardt, 1963) y su bondad se evaluó a partir de la convergencia de las iteraciones del programa de regresión no lineal en una solución, del valor del coeficiente de determinación no lineal ajustado (R2) y de la normalidad (prueba ómnibus de D’Agostino & Pearson) y aleatoriedad (test de ciclos o rachas) de la distribución de los residuales. Paralelamente se procedió a efectuar los mismos ajustes con los datos de peso corporal y peso del huevo correspondientes a dos genotipos de gallinas reproductoras pesadas -Ross308® y Cobb500® - extraídos de las guías de manejo publicadas por las respectivas compañías comerciales, Aviagen (2017) y Cobb-Vantress (2013), respectivamente.
Análisis estadístico
A los efectos del análisis, los estimadores de los parámetros con significado biológico, peso corporal asintótico (Ap) y tasa de maduración para peso corporal (k), peso asintótico del huevo (Ah), rango de peso del huevo entre el inicio de la postura y la asíntota (B) y de tasa de maduración para peso del huevo (r) se trataron como nuevas variables aleatorias. Los valores promedio de los estimadores antes mencionados se compararon con los valores de referencia correspondientes a las reproductoras comerciales con una prueba t de Student para una única muestra poblacional.
RESULTADOS Y DISCUSIÓN
Patrón dinámico de crecimiento dimensional
Las aves Campero Casilda crecieron hacia un peso asintótico Ap= 3.468 ± 33,7 g (media aritmética ± error estándar) el que resultó significativamente inferior al correspondiente a los dos genotipos de referencia (Ross308®: 4.168 g; t= 20,8; p< 0,0001 y Cobb500®: 4.119 g; t= 19,3; p< 0,0001). Contrariamente a lo esperado, dada la habitual asociación negativa entre el tamaño asintótico y la velocidad para alcanzarlo, Campero Casilda presentó también una menor tasa de maduración para peso corporal (k= 0,0816 ± 0,00152 g-1) que Ross308® (k= 0,0943 g-1; t = 8,322; p< 0,0001) y Cobb500® (k= 0,1009 g-1; t = 12,68; p< 0,0001). La Fig. 1 resume gráficamente el comportamiento de las curvas teóricas de crecimiento de los tres grupos genéticos.
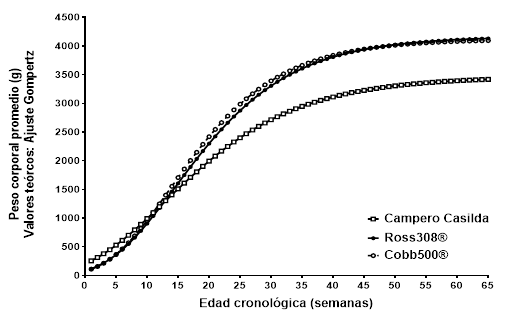
Fig. 1 Patrón de crecimiento dimensional teórico de gallinas Campero Casilda y dos genotipos comerciales de reproductoras pesadas
Patrón dinámico de aumento de peso del huevo en función de la edad de postura
Campero Casilda presentó un valor asintótico (Ah= 72,8 ± 0,83 g) que no difirió significativamente del correspondiente a las reproductoras Ross308® (Ah= 71,9 g; t= 1,117; p= 0,245), pero fue mayor al de Cobb500® (Ah= 70,8 g; t= 2,465; p= 0,015). A diferencia del parámetro k de la función sigmoidea de Gompertz que muestra una relación directa entre el valor de su estimador y la tasa de maduración para peso corporal (a mayor valor de k, mayor velocidad de aproximación al peso corporal asintótico Ap), el parámetro r de la función exponencial asintótica de Weatherup & Foster presenta una asociación inversa con la tasa de maduración para peso del huevo (a mayor valor de r, menor velocidad de aproximación al peso asintótico del huevo Ah). Campero Casilda presentó un valor promedio del estimador de la tasa de maduración para peso del huevo (r= 0,9396 ± 0,00452 g-1) que no difirió significativamente del correspondiente a las reproductoras Ross308® (r= 0,9397 g-1; t= 1,351; p= 0,180) pero fue mayor al de Cobb500® (r= 0,9056 g-1; t= 6,196; p< 0,0001) indicando similar velocidad de aproximación al peso asintótico del huevo Ah en el primer caso y menor en el segundo. El rango de aumento del peso del huevo (b) entre el primer huevo y el valor asintótico de los mismos (Ah) de Campero Casilda (21,3 ± 0,67 g) fue similar al de los dos genotipos de referencia (Ross308®: 21,4 g; t= 0,178; p= 0,859 y Cobb500®: 21,2 g; t= 0,179; p= 0,882). Por último, el peso estimado promedio del primer huevo (Ah - B) de Campero Casilda (51,6 ± 0,55 g) fue levemente mayor al de Ross308® (50,5 g; t= 1,930; p= 0,056) y significativamente mayor al de Cobb500® (49,6 g; t= 3,574; p= 0,005). La Fig. 2 resume gráficamente el comportamiento de las curvas teóricas de modificación del peso del huevo de los tres grupos genéticos.

Fig. 2 Patrón teórico de aumento de peso del huevo de gallinas Campero Casilda y dos genotipos comerciales de reproductoras pesadas
La propuesta de una producción avícola doble-propósito se enfrenta con el desafío de compatibilizar crecimiento y producción, dos aspectos que guardan entre sí una asociación genética desfavorable. La comparación del comportamiento productivo de Campero Casilda con reproductoras comerciales se justifica por su condición de aves pesadas. Si bien la comparación de las curvas de postura (Romera et al., 2020) mostró que en el pico de producción presentan un porcentaje de postura 4,7 % menor que Ross308® y 4,4% menor que Cobb500® y que alcanzan dicho pico tres semanas después que Ross308® y dos semanas después que Cobb500®, la misma es compatible con la propuesta de utilizarla como ponedoras. Sumada a esta evidencia, los resultados de este trabajo muestran que, en comparación con los dos genotipos de referencia, las aves Campero Casilda presentan un patrón de crecimiento caracterizado por un menor peso asintótico y una menor tasa de maduración o velocidad de aproximación a dicho peso. Ante este comportamiento común del cruzamiento experimental frente a ambas reproductoras comerciales, el peso de los huevos puestos por las aves Campero Casilda en comparación con Ross308®, aumenta hacia un valor asintótico similar y con similar velocidad, con un rango entre el peso del primer huevo y el peso asintótico del mismo orden y un peso teórico estimado del primer huevo algo mayor; mientras que en comparación con Cobb500® el peso de los huevos puestos por Campero Casilda presenta un patrón de modificación caracterizado por dirigirse hacia un mayor valor asintótico y hacerlo con menor velocidad, con un rango de aumento entre el peso del primer huevo y el peso asintótico coincidente y un mayor peso teórico del primer huevo. Estos resultados ponen en evidencia cierta independencia en la base genética de naturaleza poligénica que controla la dinámica del peso corporal y el peso del huevo, ya observada en trabajos previos (Di Masso, et al, 1998) lo que plantea la posibilidad de combinar en una misma gallina patrones dinámicos más favorables de ambas variables de indudable trascendencia productiva en la avicultura de puesta.
CONCLUSIONES
En términos productivos, y en comparación con los genotipos comerciales de referencia, el cruzamiento de tres vías Campero Casilda presenta una combinación favorable de los dos patrones dinámicos -peso corporal y peso del huevo- estudiados. Su menor peso corporal representa una ventaja porque, dada la asociación positiva entre peso y consumo, estaría asociado a un menor costo de la alimentación. Paralelamente, pese a la asociación positiva entre peso corporal y peso del huevo, el menor peso de las aves no repercute negativamente sobre el peso del huevo lo que transforma a Campero Casilda, al menos con referencia a estos dos caracteres expresados por las hembras, en una opción como genotipo doble propósito en el contexto de modelos avícolas alternativos.
AGRADECIMIENTOS
Los autores agradecen la responsable colaboración de los estudiantes de la carrera de Medicina Veterinaria de la Facultad de Ciencias Veterinarias de la Universidad Nacional de Rosario, Argentina, en especial a aquellos participantes del Programa de Becas de Promoción de las Actividades Científicas y Tecnológicas, que con su dedicación posibilitaron la concreción de este Trabajo.
El trabajo fue parcialmente financiado por la Universidad Nacional de Rosario (UNR) en el marco del proyecto acreditado VET228 - Evaluación productiva de gallinas del cruzamiento experimental de tres vías campero Casilda en su primer ciclo de postura y por el Instituto Nacional de Tecnología Agropecuaria (INTA) a través de un convenio interinstitucional INTA- UNR.
El autor principal, alumno del Doctorado en Ciencias Veterinarias de la Universidad Nacional de Rosario, agradece al Consejo Interuniversitario Nacional el otorgamiento de una beca doctoral del Programa Estratégico de Formación de Recursos Humanos en Investigación y Desarrollo (PERHID)

