










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
De acuerdo con Thornton et al. (2022) los efectos del stress climático se han convertido en un obstáculo de envergadura que representa pérdidas de alrededor del 10% del valor comercial de la leche y la carne en todo el sector de producción animal a nivel mundial, lo que explica la cantidad de esfuerzos que se dedican a esta temática. Las tendencias son las mismas para el área del Caribe y América Central (UNEP, 2008) donde los efectos colaterales pueden también afectar gran parte del hábitat y áreas geográficas de una gran cantidad de islas grandes o pequeñas. En Cuba, las predicciones del cambio climático son similares (Roura-Pérez et al., 2020) y de acuerdo a Milera (2011) este se manifestará en toda la actividad agrícola en general y en el sector ganadero afectando los rendimientos de los pastos y forrajes que son los componentes básicos de la alimentación del ganado, lo cual explica una parte de los marcados decrecimientos en los niveles productivos de carne y leche que se manifestaron en el país (Sabina Roque, 2017).
El ganado Charoláis es una de las más reconocidas razas francesas y ha estado presente en Cuba desde inicios de la década del 1900 y por más de un siglo su principal rebaño ha estado situado en Jiguaní, provincia de Granma. Desde entonces ha manifestado un adecuado comportamiento y adaptación a las condiciones ambientales del país, manteniendo su potencial carnicero el cual fue corroborado en una comparación realizada con el Charolais de Francia (Renand et al., 1997), estos resultados representan el alto mérito de esta raza en la producción de carne del país. En esta región existen evidencias de un empeoramiento de las condiciones climáticas a lo largo del tiempo (Suárez-Tronco et al., 2021) que pueden producir igualmente stress térmico (ST) sobre el comportamiento del vacuno lo cual fue demostrado por Suárez-Tronco et al. (2022a, b) en animales de la raza Criollo explotado en la misma finca, pero en potreros separados. Estos autores aplicaron un índice que combina la temperatura ambiente y la humedad relativa reconocido como ITH, estimando negativos efectos significativos sobre el peso vivo al destete (PD) y a 18 meses de edad en esta raza de reconocida adaptación al trópico, de manera que sería de utilidad examinar este mismo problema de ST sobre el ganado Charoláis.
Los resultados de ST en el ganado Criollo son coincidentes con lo publicado por Santana et al. (2016) en las condiciones de Brasil todo lo cual es coherente con el enfoque y conclusiones generales de que los programas de selección que producen animales muy especializados, manifiestan efectos colaterales negativos en otros rasgos de interés económico y manifiestan una menor capacidad adaptativa (Rauw et al., 1998).
Existen metodologías estadísticas apropiadas para la estimación de los componentes genéticos ligados a ST a partir de los resultados originales sobre este tema presentados por Ravagnolo y Misztal (2000) con datos de producción mensual de leche en ganado Holstein, pero las características intrínsecas de los sistemas de producción de carne requieren de otros puntos de vista ya que los rasgos de interés económico se cuantifican como resultado final de un proceso (por ejemplo, peso vivo a una edad fija). Las evidencias disponibles indican antagonismo entre niveles extremos de ST y PD (Santana et al., 2016; Suárez Tronco et al., 2022b) en razas de reconocida adaptación al calor de Brasil (Nellore y cruzamiento) y en Cuba con Criollo, mientras que Bradford et al. (2016) muestran el mismo patrón en animales Angus de Estados Unidos.
En el caso de PD es un carácter compuesto que depende tanto del genotipo del animal que lo manifiesta, sino también de otro componente, de naturaleza ambiental con respecto al ternero, pero genético ligado a la producción de leche de su madre. Ambos efectos genéticos fueron incluidos en el estudio de Santana et al. (2016) quienes además utilizaron los efectos acumulativos del ITH (CCATHI) recibidos por el animal que produce el registro durante los meses previos al destete, para medir la importancia de ST. Estos autores demuestran la existencia de relaciones negativas tanto fenotípicas como genéticas entre CCAITH y el PD en tres tipos de animales de reconocida adaptación a las condiciones de Brasil, la misma tendencia fue presentada por Suárez Tronco et al. (2022b) en animales Criollo, aunque este último autor no consideró los efectos genéticos maternos en su análisis.
Los estudios de componentes de (co)varianza entre efectos genéticos directos y maternos presentan muchas dificultades debidas a la estructura y distribución de los datos (Meyer, 1992), la cual puede ser aún más compleja si consideramos la estimación de esos parámetros a lo largo de una escala de niveles climáticos, no obstante, es necesario alguna alternativa que permita estimar el mérito genético directo de los animales frente a una trayectoria de niveles de ambiente.
Los objetivos de este estudio es evaluar el efecto del ST sobre el peso al destete en animales Charolais y estimar los componentes de (co)varianza genética a lo largo de la trayectoria del nivel de CCAITH recibido por el animal durante los 6 meses previos al destete.
MATERIALES Y MÉTODOS
Datos productivos: Para este estudio se dispuso de 22 960 registros de PD de animales Charoláis puros pertenecientes a las bases de datos del programa de evaluación genética de esta raza. De este total se seleccionaron 19 466 observaciones pertenecientes a la empresa Manuel Fajardo, situada en Jiguaní, provincia Granma, donde se encuentra el rebaño principal de esta raza. Estos datos fueron adicionalmente depurados eliminándose aquellos registros fuera de ±3,2 desviaciones del PD, este conjunto de datos eran progenies de 157 sementales y 6 298 vacas, paridas entre enero de 1988 y mayo del 2017 en 16 rebaños de esa empresa, el pedigrí contenía un total de 34 451 animales.
Datos climáticos: Estos datos son los mismos que los empleados por Suárez Tronco et al. (2021) y se trata de los estimados de los promedios mensual de temperatura ambiente (Ta) en oC y humedad relativa (HR) en por ciento, los cuales se combinaron en un índice denominado como ITH según el mismo procedimiento empleado por Suárez Tronco et al (2021): ITH = 0.81*Ta + (HR/100) * (Ta - 14.4) + 46.4 donde los términos ya fueron definidos. Con este indicador se estimó la carga calórica acumulada (CCAITH) recibida por el animal durante los 6 meses previos al mes del destete (md), la formula aplicada fue:
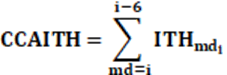
Los resultados de CCAITH tenían un total de amplitud entre 41 niveles entre CCAITH =429 hasta CCAITH =470.
Datos finales: Los registros climáticos y productivos se unieron y conformaron la base de datos que se analizaron (Tabla1). Los meses de destete se agruparon en cuatrimestre los cuales se unieron con los años y los rebaños para conformar los grupos contemporáneos (GC) cada uno de los cuales tenía un mínimo de 5 registros con más de un semental representado.
Tabla 1 Indicadores generales de la base de datos disponible.
| Indicador* | |
| Número de animales | 19 408 |
| Número de animales en el Pedigrí | 34 151 |
| Número de sementales | 157 |
| Número de Madres | 6 298 |
| Número de Grupos Contemporáneos | 858 |
| Número de años | 28 (1988 a 2017) |
| Número de niveles de CCA | 41 (429 a 470) |
| Edad al parto (años) | 11 (2 a 12) |
| Peso al destete kg | 146 (28) |
| Edad al destete días | 199 (15) |
*entre paréntesis amplitud y desviación estándar para peso y edad al destete.
Estos datos se analizaron mediante modelos lineales de efectos aleatorios mediante proc glm de SAS, para estimar las constantes mínimo-cuadráticas para PD y describir las curvas de respuesta a lo largo de los factores ambientales incluidos que estén libres de los efectos genéticos. El modelo básico fue el siguiente:
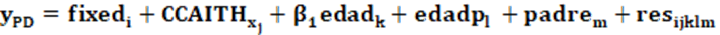 (1)
(1)
Dónde:
ypd: es la variable dependiente (peso al destete).
fixedi: representan los efectos de grupo contemporáneo (858 niveles) y el sexo de la cría (2).
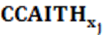 : es el efecto fijo de jth niveles de carga calórica acumulada (42 niveles).
: es el efecto fijo de jth niveles de carga calórica acumulada (42 niveles).
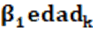 : es una covariable lineal de la edad al destete del animal.
: es una covariable lineal de la edad al destete del animal.
edadpl: es el efecto de la lth clase de edad al parto de la vaca en años (11 categorías).
padrem: es el efecto aleatorio del mth padre del ternero (157 niveles).
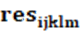 : es el efecto residual común a todas las observaciones.
: es el efecto residual común a todas las observaciones.
El análisis genético es similar al presentado previamente por Suárez Tronco et al. (2022a) y se utilizó el software ASreml 4.2 (Gilmour et al., 2015), aplicando el siguiente modelo animal de Norma de Reacción (NR),
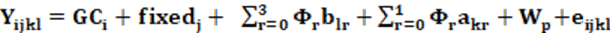 (2)
(2)
Dónde:
Yijkl representa la lth observación del PD de cada animal, GCi es la ith combinación de rebaño-año-época de destete; fixedj se incluyen los efectos fijos de edad al parto de la vaca, sexo del ternero y una covariable lineal fija de la edad al destete. La variable climática CAITH se modeló según una ecuación de regresión fija b1 expresada mediante un polinomio de Legendre 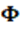 de orden r = 3 que define la curva de respuesta de toda la población del efecto de CCAITH sobre PD, Wp representa un efecto aleatorio de ambiente permanente de la madre del ternero. Por otro lado, akr es la desviación aleatoria de cada animal respecto a esa curva de respuesta mencionada, a lo largo de la trayectoria de CCAITH en este caso modelada por un polinomio de Legendre de orden r = 1 y expresada en una escala entre -1 a +1. (Ambos los brinda automáticamente el asreml). Esta akr es la solución del modelo 1 y equivale al mérito genético del animal o la Norma de Reacción para el intercepto (ai) y la pendiente (as) y se denomina función genética fg = [ai as] para este indicador de ST. Finalmente eijkl es un efecto residual aleatorio común a todas las observaciones y no correlacionado con los efectos fijos. Un modelo similar incluyendo el padre de la vaca como efecto aleatorio no resultó significativo.
de orden r = 3 que define la curva de respuesta de toda la población del efecto de CCAITH sobre PD, Wp representa un efecto aleatorio de ambiente permanente de la madre del ternero. Por otro lado, akr es la desviación aleatoria de cada animal respecto a esa curva de respuesta mencionada, a lo largo de la trayectoria de CCAITH en este caso modelada por un polinomio de Legendre de orden r = 1 y expresada en una escala entre -1 a +1. (Ambos los brinda automáticamente el asreml). Esta akr es la solución del modelo 1 y equivale al mérito genético del animal o la Norma de Reacción para el intercepto (ai) y la pendiente (as) y se denomina función genética fg = [ai as] para este indicador de ST. Finalmente eijkl es un efecto residual aleatorio común a todas las observaciones y no correlacionado con los efectos fijos. Un modelo similar incluyendo el padre de la vaca como efecto aleatorio no resultó significativo.
En este modelo 2 de NR se asume que:
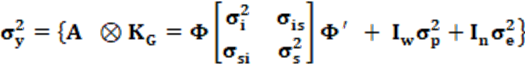
en la cual es la varianza genética, A es el denominador de la matriz de parentesco; I
w e I
n son matrices identidad de orden w (número de madres) y n (número de observaciones) y es la varianza residual. En este modelo de NR es necesario aplicar un procedimiento adicional propuesto por Jamrozik y Schaeffer (1997) mediante la matriz K
G que permite estimar los componentes de (co)varianza entre cada punto de la escala de la variable climática CCAITH expresada en términos del polinomio de Legendre de orden r = 1 indicado previamente, que contiene los elementos propios de una ecuación lineal con un significado específico:
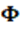 y corresponden con las varianzas genéticas para el intercepto (nivel general genético de PD a lo largo de la escala de CCAITH) y la pendiente, respectivamente que representa la Norma de reacción del animal para modificar la evolución de la varianza de PD a lo largo de la trayectoria de los diferentes niveles de CCAITH, en otras palabras, es una medida de la capacidad genética del animal a mayor o menor tolerancia al stress calórico.
y corresponden con las varianzas genéticas para el intercepto (nivel general genético de PD a lo largo de la escala de CCAITH) y la pendiente, respectivamente que representa la Norma de reacción del animal para modificar la evolución de la varianza de PD a lo largo de la trayectoria de los diferentes niveles de CCAITH, en otras palabras, es una medida de la capacidad genética del animal a mayor o menor tolerancia al stress calórico.
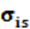 =
=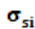 representan la covarianza genética entre el nivel genético y la tolerancia al stress calórico.
representan la covarianza genética entre el nivel genético y la tolerancia al stress calórico.
La matriz K
G es de 2x2 es compatible con la matriz de CCAITH de 41x2 de manera que manipulando las mismas se pueden estimar las (co)varianzas genéticas y los valores genéticos de cada animal en cada punto de la trayectoria de los niveles de ST que se cuantificó mediante CCAITH. Aplicando formulas clásicas (Falconer y McKay, 1996) estos estimados son:
Para la varianza genética en cada i punto de la escala de CCAITH
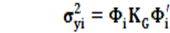
Para la covarianza genética en los puntos i y j de la escala de CCAITH
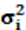
Para la heredabilidad en cada i punto de la escala de CCAITH
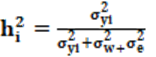
Para las correlaciones genéticas entre i y j de la escala de CCAITH
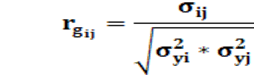
Como solución del modelo 1 previamente presentado se obtiene una función genética f
g=[a
i
a
s] para cada uno de los 34 151 animales con datos y sus antecesores sin registros, cuyos elementos son característicos de una ecuación de regresión genética lineal, es decir una matriz de 34 151x2. A partir de estos resultados y la matriz del componente genético cuyos elementos se pueden identificar como L
i y L
s para el intercepto y la pendiente, respectivamente se pueden estimar el Valor Genético (VG
i) en cada
i
punto de la trayectoria de CCAITH
VGi = [Li Ls] *[ai as]’
Con este procedimiento se pueden obtener todos los parámetros y los VG i de cada animal a lo largo de cada punto de la trayectoria de CCAITH. Estos resultados fueron sometidos a un análisis de componentes principales (ACP) utilizando MATLAB (2019) y los coeficientes de los eigenvector correspondientes a los eigenvalues de mayor importancia fueron usados como factores de ponderación de los VG i con vistas a sintetizar en un índice los resultados de VG i en toda la escala de CCAITH siguiendo la propuesta de Togashi y Lin (2006). Este nuevo índice (Ipct i) se representa de la siguiente forma general para cada ith componente principal:
donde los ev i son los coeficientes de los eigenvector de cada componente principal y los VG i ya fueron definidos previamente y se expresan en forma estandarizadas.
RESULTADOS Y DISCUSIÓN
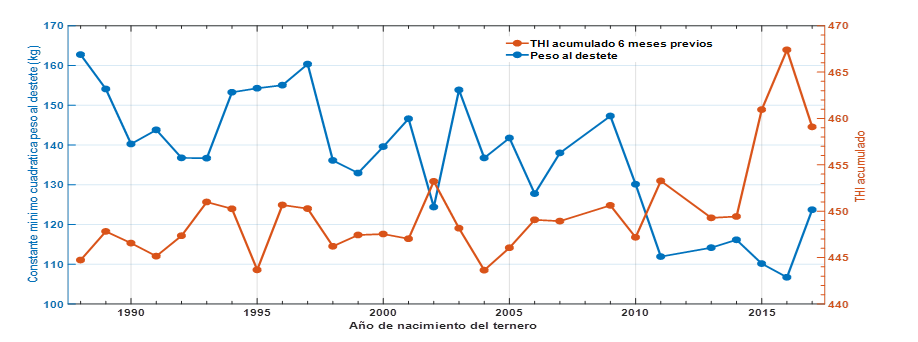
El análisis realizado según el modelo 1 de efectos fijos demuestra que la CCAITH a que estuvieron sometidos los animales durante los 6 meses previos al destete (figura 1) manifestó una respuesta más o menos estable en los primeros 11 años representados, la cual cambió a partir del 2010 cuando comenzó una tendencia de aumento en el nivel de CCAITH, en correspondencia con los cambios climáticos indicados para el país (Roura-Pérez et al., 2020), por el contrario el PD decrece significativamente durante todo el periodo siguiendo la misma respuesta que otros indicadores productivos de la ganadería vacuna del país (Sabina Roque, 2017).
Los efectos de CCAITH sobre PD fueron altamente significativos y los valores ajustados por todos los efectos incluidos en el modelo 1 se presentan en la figura 2. La curva de respuesta es francamente negativa y el análisis de regresión de las constantes mínimo cuadrada de PD ponderado por el número de registros en cada nivel CCAITH arrojó que se deprime en b=-0.807±0.04 kg por cada unidad de CCAITH (R2=90.2%). Aun con el alto nivel de ajuste se pueden diferenciar tres zonas: una fría con CCAITH<=439; una neutral con CCAITH>=440 a <=450 y una caliente CCAITH>=451, estas nuevas clases fueron incluidas en sustitución al efecto CCAITH y se aplicó el mismo modelo anterior. Los resultados fueron altamente significativos con estimados de PD= 149,3; 140,5 y 134,1 kg para las zonas fría, neutra y caliente respectivamente, la magnitud de este efecto depresivo es superior al indicado por Suárez Tronco et al. (2022a) en ganado Criollo de la misma empresa y el impacto global de esta respuesta negativa se puede inferir si se considera que en la base de datos representada, el 27,7% de los animales se mantuvieron en la zona caliente.
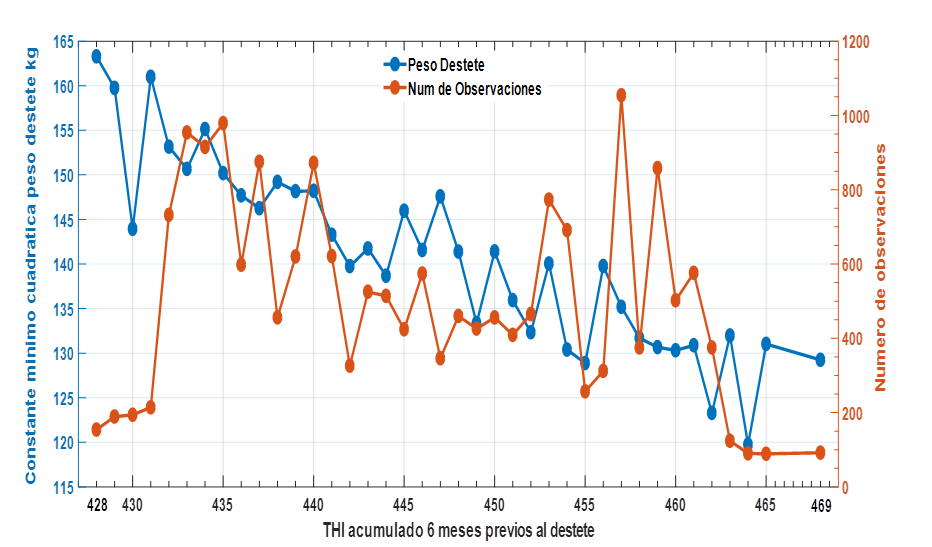
Figura 2 Constantes mínimas cuadráticas del efecto CCAITH sobre el peso al destete en animales Charoláis.
Los resultados del modelo genético arrojaron que existe una importante variabilidad genética en capacidad de tolerancia del animal a cambios en niveles de CCAITH. La heredabilidad (h2) presentó una curva decreciente en la medida que se incrementa el nivel de calor recibido por el animal durante los 6 meses previos al destete (figura 3). Esta forma de respuesta con valores máximos de h2 en ambos extremos de la escala de CCAITH, es la típica que se ha presentado en otras publicaciones con el uso de modelos de regresión aleatoria en ganado de carne y de leche (ver excelentes discusiones en Meyer, 2005 y Li et al., 2020, respectivamente) lo cual puede deberse a problemas intrínsecos de los polinomios de Legendre cuando existe una distribución desigual de los datos. En términos comparativos los estimados de la figura 3 siguen el mismo patrón presentado por Suárez Tronco et al. (2022b) en ganado Criollo en esta región, mientras que Santana et al., (2016) mostró similares tendencias, pero estimados superiores, en animales cruzados con diferentes proporciones de B. indicus en las condiciones de Brasil.
De los parámetros presentados en la figura 3 se infiere que existe mayor espacio para cualquier proceso de selección cuando el ambiente es más favorable (CCAITH zona fría), sin embargo, tales posibilidades están limitadas dado que las correlaciones genéticas disminuyen hasta alcanzar valores inferiores a 0,8 a partir de CCAITH>=450 lo cual es el límite establecido (Robertson, 1959) para indicar la existencia de Interacción genotipo ambiente (Ige).
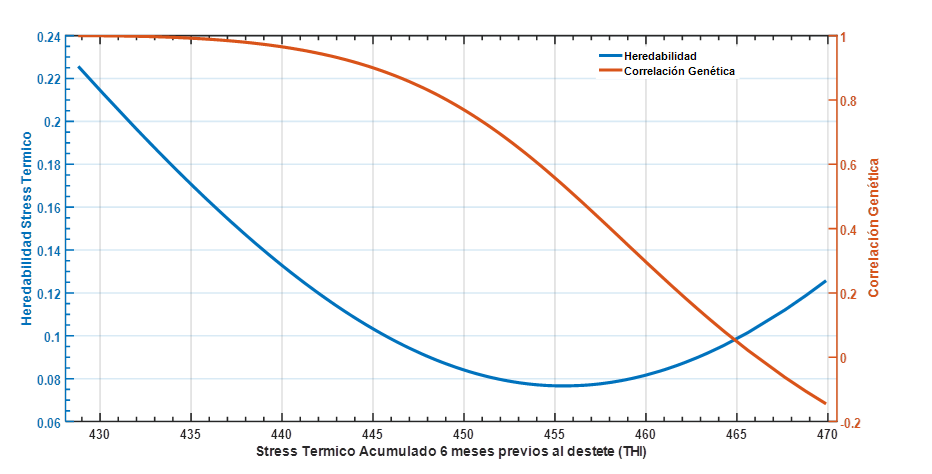
Figura 3. Estimado de heredabilidad* y correlación genética para peso al destete en animales Charoláis a lo largo de la escala de stress acumulativo durante los 6 meses previos. *Errores estándar de la h2 variaron entre 0,01 a 0,03.
La existencia de Ige es una limitante real que ha sido demostrada en diferentes escenarios de producción animal (ver excelente revisión de Fodor et al., 2023). Según estos autores la interacción ligada a efectos ambientales climáticos han sido una de las más estudiadas, en los cuales los modelos de Norma de Reacción (NR) como el aplicado en este estudio, han sido los más empleados. El uso de NR puede proporcionar resultados del VG de todos los animales en cada uno de los puntos de la trayectoria de CCITH, los cuales fueron sometidos a un ACP que arrojó que el 89,1% y 10,9% de la variabilidad de las 41 estimaciones de VG están explicados por los dos primeros eigenvalues, de manera que sus correspondientes coeficientes se pueden emplear como factor de ponderación al confeccionar el Ipc (Togashi y Lin, 2006) ya indicado en la sección previa. Este nuevo indicador facilita todo el trabajo de selección sin pérdidas importantes de la información original, existen evidencias publicadas que demuestran la utilidad de esta herramienta (Savegnago et al., 2013; Boligon et al., 2016) para enfrentar el problema de la Ige. La figura 4 presenta la variabilidad de los VG de todos los animales según los resultados en las zonas fría, neutra y caliente (figuras en la diagonal) así como las correlaciones entre tales VG (encima de la diagonal), también se presenta la variabilidad de los VG estimados según Ipc ya indicado.
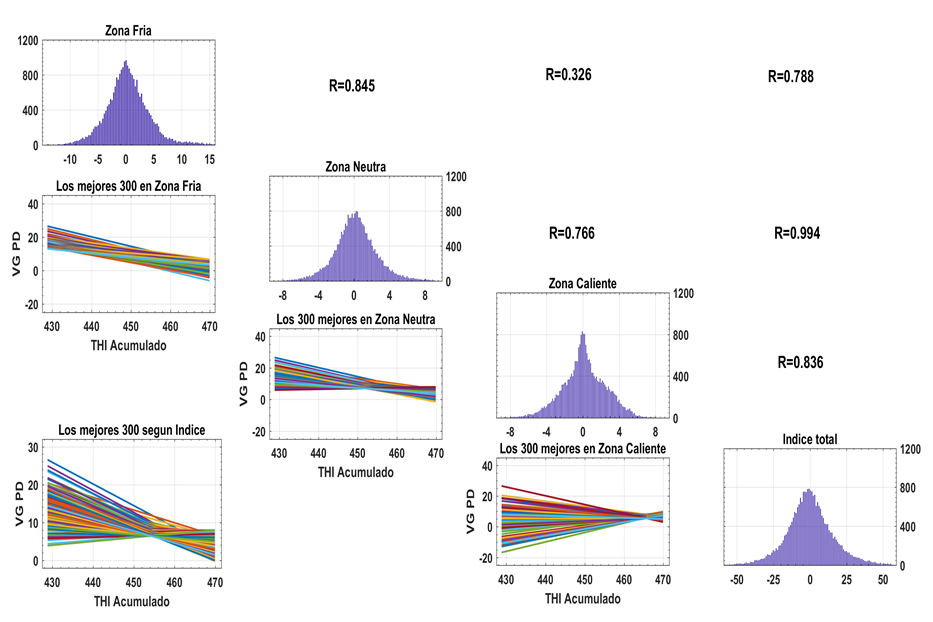
Figura 4. Variabilidad entre los Valores Genéticos* de los animales según sus resultados en las zonas fría, neutra y caliente, el Índice total, sus correlaciones y evolución de los VG a lo largo de la escala de CCAITH de los mejores 300 animales. *Los VG en la diagonal, las correlaciones encima de la diagonal y la distribución del VG a lo largo de la trayectoria de CCAITH debajo de la diagonal (detalles en el texto).
Varios elementos deben resaltarse. En primer lugar, la variación en VG es mayor en la zona fría, respecto a la neutra y caliente cuya magnitud representa el 19,3%; 12,1% y 10,4% de las medias mínimo cuadrática de estas zonas indicados previamente. Sin embargo, las correlaciones fueron muy bajas (0,33) en particular entre los niveles extremos como consecuencia de la Ige. En la parte inferior de estas distribuciones se ha representado la evolución de los VG a lo largo de la trayectoria de CCAITH para los mejores 300 animales seleccionados en cada zona, para la zona fría es evidente la uniformidad en la tendencia decreciente en la medida que se incrementa el nivel de ST, mientras que en la zona caliente manifiesta una amplitud muy marcada que alcanza animales negativos, como demostración de la Ige.
En la parte inferior derecha de la figura 4 se muestra la variabilidad genética total existente en esta base de datos en todos los niveles de CCAITH, expresada en un solo indicador Ipe el cual presenta las mayores correlaciones con el resto de las zonas, en este caso la variabilidad equivale a 0,189±0,001; 0,151±0,001 y 0,119±0,001 de PD por cada unidad de CCAITH según análisis de regresión realizado. Se seleccionaron los mejores 300 animales según Ipc y la distribución del VG de los mismos animales en cada punto de CCAITH se muestra en la parte inferior izquierda de la figura 4. La distribución simula el mismo patrón que en la zona neutral, pero su amplitud en las zonas extremas no incluye valores negativos lo cual resulta una evidencia adicional sobre las ventajas de este procedimiento de confección de un Ipc.
Los resultados mostrados pueden ser de mucha utilidad para el programa de mejora de esta raza, aunque en términos prácticos lo más recomendable es examinar las variaciones entre los sementales padres de estos animales ya que es la vía más idónea para favorecer a toda la población. Para ejemplificar esta posibilidad se seleccionaron los mejores 30 sementales según su mérito genético por Ipc y los resultados se muestran en la figura 5.
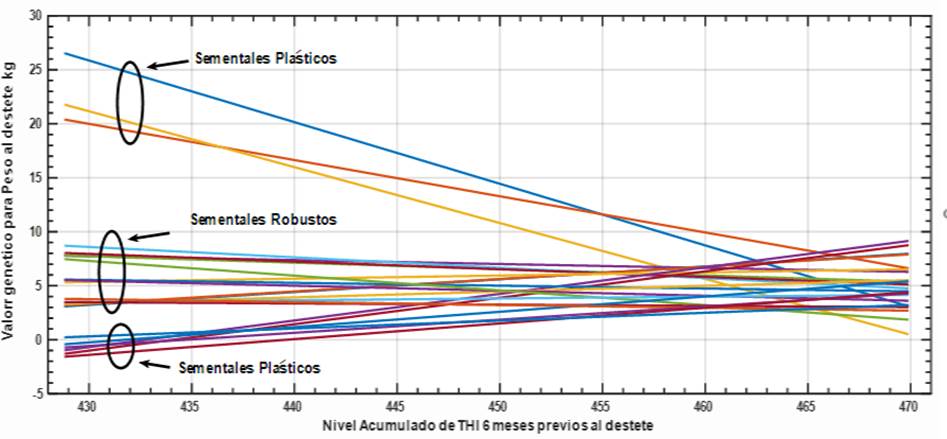
Figura 5 Valores genéticos de los 30 mejores sementales seleccionados según Ipc y su evolución a lo largo de la escala de stress climáticos 6 meses previos al destete.
De ese conjunto se identificaron 21 sementales que tenían entre 30 y 730 progenies los cuales manifiestan diferencias de forma de respuesta a la tolerancia al stress térmico recibido por sus progenies en los 6 meses previos al destete. De esta representación se pueden confeccionar dos grandes patrones de conducta, por un lado, existen un grupo principal de sementales que denominamos Robustos cuyas progenies mantienen un comportamiento uniforme a lo largo de CCAITH, los cuales deben ser la respuesta de los que atenúen los efectos de la Ige. Mientras que dentro de estos sementales élites existe otro grupo Plástico cuya respuesta puede ser contrastante. Este tipo de resultado está en correspondencias con las evidencias publicadas por Santana et al., (2016) y Suárez Tronco et al., (2022b) en animales de carne en Brasil y Cuba, respectivamente.
CONCLUSIONES
El trabajo realizado demuestra que existe un significativo efecto depresivo sobre el peso al destete en esta raza Charoláis de Cuba debido al stress acumulado por el calor recibido por el animal durante los 6 meses previos al destete. Los resultados indican la existencia de una base genética para tolerancia a este efecto climático con manifestaciones de interacción genotipo ambiente (Ige), de manera que cambia el orden de mérito en los mejores en condiciones extremas de zonas de calor a frio. La magnitud de este efecto depresivo es de -15,4 kg (10,4% respecto a la media) en los animales que se desarrollaron en la zona de frio, magnitud en modo alguno despreciable si se considera que cerca del 28 % de la muestra analizada se encontraba en esta zona climática. La utilización de un modelo de Norma de Reacción es altamente recomendable en el procedimiento de evaluación genética de esta raza, el cual ha brindado la información necesaria para identificar sementales Robustos cuyas progenies manifiestan un alto nivel de tolerancia con un comportamiento uniforme a lo largo de CCAITH, los cuales deben ser la respuesta de los que atenúen los efectos de la Ige.

