










 Servicios personalizados
Servicios personalizados texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Ecuador es uno de los 17 países megadiversos del planeta, debido a su ubicación en el centro del mundo, a la presencia de la Cordillera de los Andes, la Región Costa, selvas amazónicas y las Islas Galápagos (Aguirre 2018), posee una gran riqueza florística dada por 18 568 especies de plantas vasculares de las cuales el 25 % son endémicas. Gran parte de esta diversidad y endemismo de plantas se desarrollan en el bosque andino, por lo que representa uno de los hot spots de biodiversidad en los Andes del Norte más amenazado (Tejedor-Garavito et al., 2012).
En el caso de la Hoya de Loja, el bosque andino, se encuentra en peligro de desaparecer, cerca de 3 459 hectáreas han sido deforestadas durante el período 2014 2016, por la conversión de uso para la expansión agropecuaria, incendios forestales y ampliación de la zona urbana (MAE 2018). La desaparición de estos no solo ocasiona la disminución de especies forestales, sino que también afecta a uno de los grupos ecológicos importantes como son las epífitas vasculares (MAE 2016 y Paredes-Ulloa et al., 2021).
Estudiar las epífitas vasculares es importante no solo porque son un componente de la diversidad en los bosques tropicales y andinos, sino por la función que cumplen en la dinámica recuperativa de estos, ya que, al estratificarse verticalmente, constituyen el hábitat para la fauna y ofrecen una gran variedad de nichos y recursos (Einzmann y Zotz 2016) para otros grupos. Por ello, el bosque andino del Parque Universitario "Francisco Vivar Castro" a una altitud de 2 250 m s.n.m., con una extensión de 12,93 ha (Aguirre et al., 2016) constituye un sitio de interés para el estudio de este grupo ecológico.
Bajo estas consideraciones, el estudio tiene como objetivo evaluar la composición, estructura y preferencia de hábitat de las epífitas vasculares en una parcela permanente de una hectárea del bosque andino.
MATERIALES Y MÉTODOS
Área de estudio
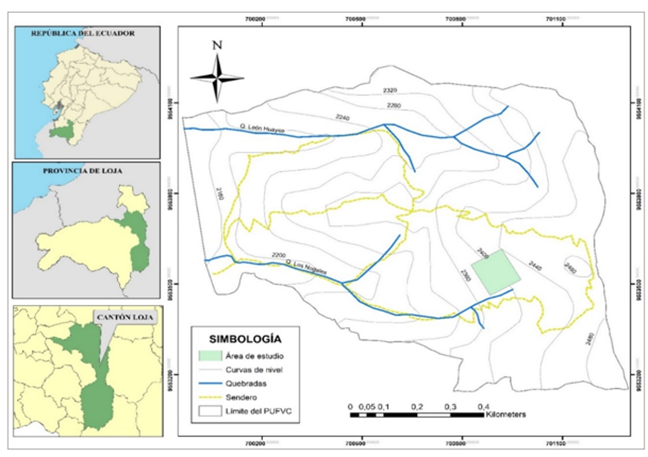
El estudio se realizó en el Parque Universitario "Francisco Vivar Castro" (PUFVC), propiedad de la Universidad Nacional de Loja, en la provincia de Loja, localizado en las coordenadas UTM: 700 5929 554 223 N, 700 9709 553 139 S, 701 3099 553 171 E, 699 9619 554 049 W, entre 2 130 a 2 520 m s.n.m de altitud y ocupa una superficie de 99,13 ha (Figura 1). Esta área es parte del Proyecto: Procesos ecológicos de la vegetación en el parque universitario "Francisco Vivar Castro", Universidad Nacional de Loja. Fase III. 2022-2023.
Selección de los forófitos
Para esta investigación se tomó como base la información del censo de los árboles de una parcela permanente de una hectárea en el bosque andino del PUFVC. En cada una de las 25 subparcelas de 20 x 20 m (400 m2), se seleccionaron tres árboles (forófitos), considerando cuatro aspectos: D1,30 m superior a 20 cm, amplitud de la copa y visibilidad (altura de copa y diámetro de copa) y ramificación (Figura 2).

Evaluación de ensambles de epífitas vasculares en cada forófito
La evaluación de ensambles de epífitas presentes en cada forófitos se efectuó mediante observación directa con binoculares y ascenso a cada árbol en estudio. Se registraron los individuos de epífitas vasculares (bromelias, orquídeas, aráceas y piperáceas) presentes en cada forófitos. Estos fueron colectados para su posterior identificación en el Herbario "Reinaldo Espinosa" de la Universidad Nacional de Loja.
Registro de datos para los parámetros estructurales
Dado a que la distribución de las epífitas sobre el forófito varía vertical y horizontalmente, cada árbol seleccionado en las subparcelas, se subdividió en tres secciones (Figura 3): sección 1 (Sa1) desde la base hasta 3 m; sección 2 (Sa2) es la parte media, por encima de la sección 1 y por debajo de la primera rama; y sección 3 (Sa3) desde la primera rama hasta la corona del árbol, esto es una modificación al método de Johansson (1974).

Análisis de datos para la composición y estructura
Con los datos colectados de las epífitas vasculares en cada uno de los árboles de las subparcelas se calcularon los parámetros estructurales: densidad relativa (DR), frecuencia relativa (FR) e IVI. Se realizó la curva de acumulación de especies usando el índice de chao 2 para comprobar la representatividad del muestreo. Para comparar la densidad entre sección vertical de los forófitos, la riqueza por familias botánicas y la densidad por especies de epífitas, se realizó una prueba no paramétrica de Kruskal-Wallis con un nivel de confianza del 5 %. Los análisis estadísticos se utilizaron con el paquete estadístico EstimateSWin910.
Se definieron las epífitas vasculares que mayor preferencia tienen hacia un hospedero(s) y, se calculó el porcentaje de preferencia a través de una regla de tres directa, debido a que las magnitudes son directamente proporcionales. Además, se identificaron los forófitos con mayor presencia de individuos epífitos en cada sección, las especies con mayor abundancia de individuos y, las secciones que prefieren estas especies.
Se realizó un análisis de conglomerados para reconocer la existencia de grupos entre forófitos de acuerdo a las especies de epífitas que lo habitan; y otro entre las tres secciones. Para esto se utilizó el método de encadenamiento completo (complete linkage) y se consideró como punto de corte o línea de referencia para la definición de los conglomerados el 50 % de similitud y el 80 % respectivamente.
RESULTADOS
Composición florística y estructura de las epífitas vasculares
En la Figura 4, se presenta la curva de acumulación de especies, que muestra el esfuerzo de muestreo, esta curva se ajustó con el estimador Chao 2. La riqueza observada es de 12 especies, el estimador no paramétrico Chao 2 estima en 18 especies para la muestra de 75 hospederos. El muestreo no permitió abarcar toda la diversidad de epífitas, registrándose el 67 % de las especies esperadas.

Se registraron 12 especies de epífitas dentro de ocho géneros y cuatro familias. La cantidad de epífitas registradas fue de 7 610 individuos en 75 forofitos inventariados. En la Tabla 4 se identifican las 21 especies hospederas de epífitas vasculares, con la densidad en el muestreo y densidad estimada por hectárea (Tabla 1).
Tabla 1. - Número de individuos de epífitas en el muestreo y la densidad estimada para una hectárea en el bosque andino del PUFVC
| Especie hospedera | No.E/H | No.E/m | No.H/ha | No.E/ha |
|
|
22 | 3 830 | 51 | 8 879 |
|
|
1 | 17 | 8 | 136 |
|
|
4 | 1 313 | 11 | 3 611 |
|
|
7 | 476 | 80 | 5 440 |
|
|
1 | 55 | 6 | 330 |
|
|
4 | 56 | 8 | 112 |
|
|
2 | 8 | 64 | 256 |
|
|
2 | 54 | 15 | 405 |
|
|
1 | 20 | 39 | 780 |
|
|
1 | 4 | 1 | 4 |
|
|
2 | 65 | 23 | 748 |
|
|
4 | 497 | 73 | 9 070 |
|
|
1 | 70 | 7 | 490 |
|
|
2 | 173 | 30 | 2 595 |
|
|
5 | 128 | 44 | 1 126 |
|
|
2 | 263 | 19 | 2 499 |
|
|
2 | 62 | 14 | 434 |
|
|
4 | 213 | 36 | 1 917 |
|
|
1 | 42 | 66 | 2 772 |
|
|
6 | 211 | 22 | 774 |
|
|
1 | 53 | 2 | 106 |
Leyenda: No.E/H:Número de individuos de epífitas por hospedero, No.E/m:Número de individuos epífitos de la muestra, No.H/ha:Número de hospederos estimados por hectárea; No.E/ha:Número de individuos epífitos estimados para 1 ha.
Estructura vertical de las epífitas vasculares por secciones
La Tabla 2 muestra las epífitas más abundantes por sección, de manera general en la sección 3 hubo mayor número de individuos de epífitas y la presencia única de orquídea Cyrtidiorchis rhomboglossa. En la sección 1 las especies más abundantes fueron de la familia Piperaceae: Peperomia alata y Peperomia galioides, seguida de Anthurium oxybelium; mientras que las menos abundantes correspondían a las familias Bromeliaceae y Orchidaceae. En la sección 2 como en la 3, las epífitas vasculares con mayor cantidad de individuos correspondieron a Tillandsia tovarensis (Bromeliacea), Cyrtochilum aureum y Pleurothallis maxima. Las especies con menos individuos fueron
Tillandsia cylindrica, Stelis emarginata y Epidendrum sp.
Tallandsia tovarensis es la única especie que muestra un incremento drástico en cuanto al número de individuos, en particular entre la sección 3 y el resto de las secciones, presentando una marcada presencia en todos los forófitos de la parcela. Las especies mejor representadas en el número de individuos se corresponden con Pleurothallis máxima y Cyrtochilum aureum que tienden a aumentar hacia la parte superior de los árboles (Tabla 2).
Tabla 2. - Parámetros estructurales de las epífitas vasculares de la sección I en la parcela permanente del PUFVC
| Sección 1 | |||||
| Familia | Nombre científico | D | Dr % | Fr % | IVI |
| Piperaceae |
|
292 | 54,07 | 36,78 | 45,43 |
| Araceae |
|
84 | 15,56 | 22,99 | 19,27 |
| Piperaceae |
|
44 | 8,15 | 14,94 | 11,55 |
| Orchidacea |
|
43 | 7,96 | 3,45 | 5,71 |
| Orchidacea |
|
34 | 6,3 | 5,75 | 6,02 |
| Bromeliaceae |
|
28 | 5,19 | 9,2 | 7,19 |
| Bromeliaceae |
|
9 | 1,67 | 2,3 | 1,98 |
| Bromeliaceae |
|
3 | 0,56 | 1,15 | 0,85 |
| Araceae |
|
2 | 0,37 | 2,3 | 1,33 |
| Orchidacea |
|
1 | 0,19 | 1,15 | 0,67 |
| Total | 540 | ||||
| Sección 2 | |||||
| Bromeliaceae |
|
350 | 41,03 | 36,27 | 38,65 |
| Orchidacea |
|
174 | 20,4 | 17,65 | 19,02 |
| Orchidacea |
|
158 | 18,52 | 10,78 | 14,65 |
| Piperaceae |
|
53 | 6,21 | 6,86 | 6,54 |
| Piperaceae |
|
51 | 5,98 | 7,84 | 6,91 |
| Bromeliaceae |
|
31 | 3,63 | 10,78 | 7,21 |
| Araceae |
|
24 | 2,81 | 5,88 | 4,35 |
| Bromeliaceae |
|
6 | 0,7 | 1,96 | 1,33 |
| Orchidacea |
|
5 | 0,59 | 0,98 | 0,78 |
| Orchidacea |
|
1 | 0,12 | 0,98 | 0,55 |
| Total | 853 | ||||
| Bromeliaceae |
|
4 846 | 78 | 39,47 | 58,73 |
| Orchidacea |
|
547 | 8,8 | 16,45 | 12,63 |
| Orchidacea |
|
333 | 5,36 | 11,84 | 8,6 |
| Bromeliaceae |
|
279 | 4,49 | 15,79 | 10,14 |
| Bromeliaceae |
|
187 | 3,01 | 13,16 | 8,08 |
| Piperaceae |
|
12 | 0,19 | 0,66 | 0,43 |
| Piperaceae |
|
8 | 0,13 | 1,32 | 0,72 |
| Araceae |
|
3 | 0,05 | 0,66 | 0,35 |
| Orchidacea |
|
2 | 0,03 | 0,66 | 0,35 |
| Total | 6 217 |
Nota: D: Densidad; Dr:Densidad Relativa; Fr:Frecuencia relativa; IVI:Índice de valor de importancia.
Comparación de las especies por familia en cada sección de los forófitos
La prueba de Kruskal-Wallis mostró que no existen diferencias significativas en cuanto al número de especies de epífitas con respecto la sección de los forófitos. En el caso la riqueza de especies por familia, la prueba sí mostró diferencias significativas (Figura 5, P = 0,0189) las letras diferentes indican que existe diferencia entre el número de especies por familia.

Preferencia de las epífitas vasculares hacia los hospederos
Se detectó que solo cuatro especies arbóreas son más preferidas como hospedero (Tabla 3), entre estas Alnus acuminata representa uno de los hospederos con mayor preferencia para las epífitas de cualquier familia, en particular para las especies de la familia Orchidaceae.
Tabla 3. - Porcentaje de preferencia de la especie epífita al hospedero en la parcela permanente del PUFVC
| Epífitas | Hospedero | % r | No.E |
|---|---|---|---|
|
|
22,52 | 25 | |
|
|
|
50 | 1 |
|
|
|
50 | 1 |
|
|
|
38,82 | 210 |
|
|
|
100 | 2 |
|
|
|
43,87 | 154 |
|
|
|
26,61 | 29 |
|
|
|
73,40 | 549 |
|
|
|
100 | 5 |
|
|
|
43,57 | 139 |
|
|
|
21,32 | 68 |
|
|
|
50,80 | 2 654 |
Nota: % Pr:porcentaje de preferencia, No.E:Número de individuos de epífitas
Similitud entre secciones y entre hospederos por carga epífita
Según la posición de la línea de corte en el dendrograma del análisis de conglomerados se distinguen dos grupos (Figura 6), uno que agrupa la sección 1 (Sa1) y sección 2 (Sa2) y otro para la sección 3 (Sa3).

El análisis de conglomerados de las especies epífitas con relación al hospedero (Figura 7 y Tabla 4), muestra que con un 50 % de similitud se identifican 8 grupos. Con igual asociación de epífitas se reconocen los forófitos Oreopanax rosei, Axinaea macrophylla y Morella interrupta y el grupo formado por Vismia baccifera, Alnus acuminata y Nectandra laurel. El forófito Siparuna muricata que más se diferencia en cuanto a especies epífitas con relación al resto.
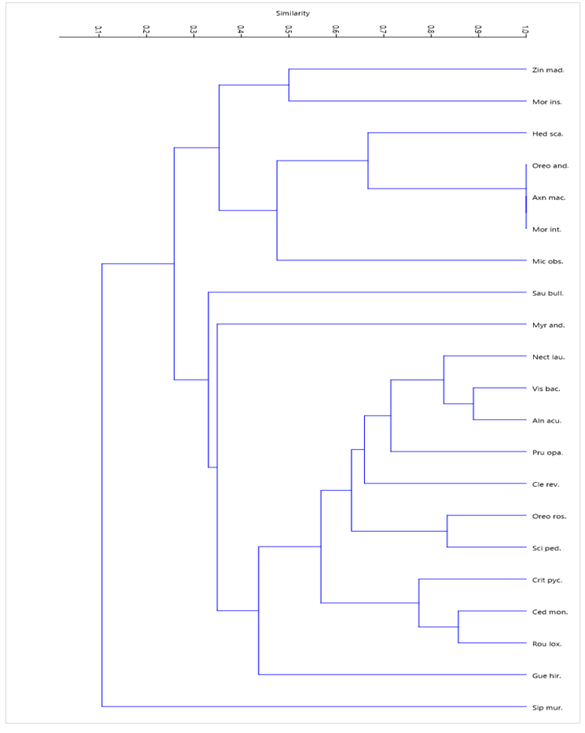 Leyenda: Sip mur.: Siparuna muricata (Ruiz y Pav.) A. DC, Gue hir.: Guettarda hirsuta (Ruiz y Pav.) Pers., Rou lox.: Roupala loxensis I.M. Johnst., Ced mon.: Cedrela montana Moritz ex Turcz, Cri pyc.: Critoniopsis pycnantha (Benth.) H. Rob., Sci ped.: Sciodaphyllum pedersenii, Oreo ros.: Oreopanax rosei Harms, Cle rev.: Clethra revoluta (Ruiz y Pav.) Spreng, Pru opa.: Prunus opaca (Benth.) Walp., Aln acu.: Alnus acuminata Kunth, Vis baca.: Vismia baccifera (L.) Triana y Planch., Nec lau.: Nectandra laurel Klotzsch ex Nees, Myr and.: Myrsine andina (Mez) Pipoly, Sau bull.: Saurauia bullosa Wawra, Mic obs.: Miconia obscura (Bonpl.) Naudin, Mor int.: Morella interrupta (Benth.) Lægaard, Axn mac.: Axinaea macrophylla (Naudin) Triana, Ore and.: Oreopanax andreanus Marchal, Hed scan.: Hedyosmum scabrum (Ruiz y Pav.) Solms, Mor ins.: Morus insignis Bureau, Zin mad.: Zinowiewia madsenii C. Ulloa y P. Jørg.
Leyenda: Sip mur.: Siparuna muricata (Ruiz y Pav.) A. DC, Gue hir.: Guettarda hirsuta (Ruiz y Pav.) Pers., Rou lox.: Roupala loxensis I.M. Johnst., Ced mon.: Cedrela montana Moritz ex Turcz, Cri pyc.: Critoniopsis pycnantha (Benth.) H. Rob., Sci ped.: Sciodaphyllum pedersenii, Oreo ros.: Oreopanax rosei Harms, Cle rev.: Clethra revoluta (Ruiz y Pav.) Spreng, Pru opa.: Prunus opaca (Benth.) Walp., Aln acu.: Alnus acuminata Kunth, Vis baca.: Vismia baccifera (L.) Triana y Planch., Nec lau.: Nectandra laurel Klotzsch ex Nees, Myr and.: Myrsine andina (Mez) Pipoly, Sau bull.: Saurauia bullosa Wawra, Mic obs.: Miconia obscura (Bonpl.) Naudin, Mor int.: Morella interrupta (Benth.) Lægaard, Axn mac.: Axinaea macrophylla (Naudin) Triana, Ore and.: Oreopanax andreanus Marchal, Hed scan.: Hedyosmum scabrum (Ruiz y Pav.) Solms, Mor ins.: Morus insignis Bureau, Zin mad.: Zinowiewia madsenii C. Ulloa y P. Jørg.Fig. 7. - Similitud entre hospederos por presencia de epífitas en la parcela permanente
El 75 % de las epifitas prefiere como hospedero a Alnus acuminata, a Cedrela montana el 33 %, y Cletra revoluta el 25 %, el resto de especies arbóreas son usadas como hospedero indistintamente (Tabla 4).
Tabla 4. - Porcentaje de preferencia de las especies epifitas hacia hospederos en la parcela permanente del bosque andino del PUFVC
| Especies epífita | Hospedero | % Preferencia |
|
|
22,52 | |
|
|
21,62 | |
|
|
11,71 | |
|
|
50 | |
|
|
50 | |
|
|
9,91 | |
|
|
38,82 | |
|
|
21,26 | |
|
|
13,68 | |
|
|
100 | |
|
|
100 | |
|
|
73,40 | |
|
|
10,16 | |
|
|
100 | |
|
|
43,87 | |
|
|
12,82 | |
|
|
11,97 | |
|
|
1,14 | |
|
|
11,97 | |
|
|
26,61 | |
|
|
10,09 | |
|
|
11,93 | |
|
|
22,94 | |
|
|
9,17 | |
|
|
43,57 | |
|
|
12,23 | |
|
|
16,30 | |
|
|
10,03 | |
|
|
3,14 | |
| Morella interrupta (Benth.) Lægaard | 0,31 | |
|
|
16,30 | |
|
|
21,32 | |
|
|
22,57 | |
|
|
50,80 | |
|
|
19,47 | |
|
|
0,06 | |
|
|
0,17 | |
| Oreopanax andreanus Marchal | 16,30 | |
|
|
1,01 | |
|
|
0,92 |
DISCUSIÓN
Composición florística y estructura de las epífitas vasculares
En este estudio se encontraron individuos de especies de epífitas que muestran preferencias por determinados forófitos y secciones dentro de los forófitos, con mayor abundancia en la copa de los árboles. Se determinó la presencia de 12 especies de 8 géneros y 4 familias, registradas en 75 árboles muestreados. Estudios sobre epífitas en bosques andinos ecuatorianos no son tan abundantes en particular por la dificultad del trabajo de campo (Paredes-Ulloa et al., 2021). Al respecto, Henao-Díaz et al. (2012) reportó una diversidad y densidad similar de epífitas vasculares en bosques subandinos y de tierras bajas colombianos y argumentó que esto estaba en relación directa a la altura del hospedero y altitud sobre el nivel del mar. Sin embargo, el presente estudio muestra una menor riqueza de especies, lo cual pudiera deberse al grado de intervención del bosque y su condición en recuperación (Aguirre et al., 2016). Dichas alteraciones a los bosques naturales, que producen cambios en los elementos de su estructura, afectan a todas las comunidades dependientes de esos ecosistemas, principalmente a las epífitas (Bartels 2012).
Las familias con mayor número de especies e individuos son Bromeliaceae y Orchidaceae, esto es similar a lo reportado por Paredes-Ulloa et al. (2021), lo cual parece ser una regularidad de los bosques andinos de Ecuador. Esta representatividad está influenciada por las adaptaciones morfológicas, ecológicas y fisiológicas que estas especies epífitas presentan para poder responder a diversas restricciones ambientales como es la exposición de la luz, baja disponibilidad de nutrientes, cambios de temperatura y en especial la precipitación (Ding et al., 2016).
La alta abundancia de Tillandsia tovarensis en el bosque puede ser explicada por la tendencia de la especie a formar conglomerados los que en ocasiones cubren casi por completo las copas de los forófitos, no permitiendo el crecimiento de otras especies en su alrededor. El género Tillandsia por la característica de formar tanques o reservorios con sus hojas, son la clave para reproducción y el desarrollo a través de la retención de agua y nutrientes (Ding et al., 2016).
En cuanto a la riqueza por secciones en los forófitos muestreados, existe gran similitud en cuanto al número de especies en las tres secciones de los árboles que determina que la variación en la riqueza por sección vertical no es significativa. Este resultado diferente a Paredes-Ulloa et al. (2021) en un bosque siempreverde piemontano donde claramente se concentran las epífitas (orquídeas) en el dosel superior. Sin embargo, con relación a la densidad de epífitas hubo diferencias significativas entre secciones (Figura 6), por lo que se reconoce la existencia de una selectividad de individuos epífitos en los sitios más altos. Este comportamiento de mayor concentración de individuos en dosel superior fue congruente con los estudios de Krömer et al. (2007) y Paredes-Ulloa et al. (2021), lo cual debe relacionarse con la mejor disponibilidad de luz para las actividades vegetativas y fotosintéticas.
De modo que, en la distribución vertical se observa la preferencia de las especies a una sección en especial, un ejemplo de esto son las bromelias que están presentes en todas las secciones, pero en mayor frecuencia y densidad en la superior. Además, en secciones altas la velocidad del viento y la radiación solar aumentan mientras que la humedad relativa disminuye, lo que proporciona la formación de microclimas en las secciones del bosque situación que afecta la presencia de algunos grupos de epífitas vasculares.
En el caso de las orquídeas, no existe una generalidad para algunas especies la densidad aumenta a medida que se asciende por las secciones del hospedero como en Cyrtochilum aureum y Pleurothallis máxima. Sin embargo, esto no ocurrió con Cyrtidiorchis rhomboglossa, Epidendrum sp. y Stelis emarginata, todas estas orquídeas. Al respecto Rasmussen y Rasmussen (2018) y Paredes-Ulloa et al. (2021) reportan que esta familia tiende a tener mayor riqueza en especies, pero con poca densidad de individuos. Otra posible causa del limitado número de individuos pudiera relacionarse a morfología externa ya que estas especies tienen un pobre anclaje Rasmussen y Rasmussen (2018). La mayor frecuencia y densidad de Peperomia alata y Peperomia galioides, en la sección 1 de los forófitos pudiera estar relacionado con la mayor humedad (Mai et al., 2016 y Martínez-Bautista et al., 2019).
La diversidad de especies en las diferentes secciones fue media a baja en términos de composición y abundancia, respectivamente en comparación con los resultados de Rasmussen y Rasmussen (2018), y puede ser la respuesta a alteraciones del bosque en los últimos 30 años ya que la riqueza se asocia positivamente con la edad de la sucesión (Paredes-Ulloa et al., 2021).
Preferencia de hábitat por parte de las epífitas vasculares
Características como presencia de árboles adultos emergentes de ramificaciones abundantes y gruesas, corteza rugosa determinan la presencia de epífitas, en particular su riqueza (Armijos et al., 2017). Al respecto, Krömer et al. (2007) y Rasmussen y Rasmussen (2018) reportan que elementos que faciliten la colonización de epífitas son determinantes en la preferencia del hábitat como el tamaño del árbol, pues regula la intensidad de luz captada por las epífitas del dosel o del interior, la estructura del árbol, presencia del número de ramas gruesas, una corteza rugosa, la presencia en esta de metabolito que favorezcan o inhiban su desarrollo y sin dejar de mencionar la retención de agua y nutrientes. Sin embargo, Paredes-Ulloa et al. (2021) plantean las características más importantes es el diámetro del hospedero facilita la presencia de mayor cantidad de epífitas.
La abundancia de Alnus acuminata como forófito preferido por nueve especies de epífitas vasculares, pudiera relacionarse no solo a la abundancia de las especies arbórea en el área sino además a la fisonomía de los ejemplares, pues en su gran mayoría tenían gran porte muy ramificado, con corteza semirugosa y espacios entre las ramas a modo de hendidura que favorece la preferencia del forófito (Martínez-Meléndez et al., 2008 y Salazar-Ramírez et al., 2014). Por su parte, Cedrela montana, con menor abundancia en el área, tuvo un valor alto como especie de preferencia, lo cual se relaciona con la presencia de fisuras longitudinales irregulares en su tronco que determina la preferencia por su corteza y estructura del árbol.
CONCLUSIONES
En el bosque andino se registraron 7 610 individuos de epífitas vasculares en cuatro familias y 12 especies, donde Tillandsia tovarensis, Pleurothallis maxima y Cyrtochilum aureum fueron las más abundantes.
Existe un patrón de distribución en especies y abundancia en las familias estudias, Bromeliaceae y Orchidaceae hacia la sección 3, mientras que Piperaceae y Araceae se concentraron hacia las secciones 1 y 2 de los forófitos por la mayor humedad.
Se prefieren dos especies de forófitos: Alnus acuminata y Cedrela montana por su diámetro, ramificación y características de la corteza.
En el bosque andino del Parque Universitario "Francisco Vivar Castro", las epífitas vasculares tienen diversidad baja y media en las tres secciones reflejo del proceso de recuperación y crecimiento de este.

