










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936
cultrop vol.34 no.1 La Habana ene.-mar. 2013
Estudios preliminares referidos a la herencia de la resistencia a la Piriculariosis de líneas isogénicas de arroz obtenidas por cultivo de anteras
Piriculariosis resistance inheritance of rice isogenic lines obtained from anthers culture
Noraida de J. Pérez LeónI y Dr.C. RodolfoI. Castro MenduiñaI, Dra.C. Maria Caridad González CeperoII y Dra.C. Marta Álvarez GilII, y Dr.C. Manuel Aguilar PorteroIII
IInvestigadores Auxiliares de la Unidad Científico Tecnológica de Base Los Palacios.
II Investigadoras Titulares del departamento de Genética y Mejoramiento Vegetal, Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, Mayabeque, Cuba, CP 32 700.
III Investigador del Departamento de Arroz y Maíz del Centro de Investigación y Formación Agraria (CIFA) Las Torres Tomejil, Alcalá del Río, Sevilla, España.
RESUMEN
En el campo experimental de la Unidad Científico Tecnológica de Base «Los Palacios» fueron sembradas semillas de las plantas cosechadas de la primera generación de líneas isogénicas de arroz, procedentes del cultivo in vitro de anteras de plantas F2 de los cruces Amistad´82/2077, 2077/Amistad´82, Amistad´82/IR759-54-2-2 y Moroberekan/Amistad´82, con el objetivo de evaluar su comportamiento frente a infecciones naturales de P. grisea. Al parecer, no hay presencia de genes dominantes actuando en la herencia de la resistencia a la Piriculariosis y el comportamiento pudiera estar regido por la acción de genes mayores en homocigosis y de genes menores con efecto aditivo, lográndose la selección de cuatro líneas isogénicas resistentes a la enfermedad y de buen comportamiento agronómico, las que constituyen la base para la obtención de nuevos cultivares.
Palabras clave: arroz, cultivo de anteras, resistencia a la enfermedad, herencia genética.
ABSTRACT
In the Base Technological Scientist Unit «Los Palacios» seeds first generation of harvested rice isogenic lines, coming from F2 in vitro anthers culture from crossings Amistad´82/2077, 2077/Amistad´82, Amistad´82/IR759-54-2-2 and Moroberekan/Amistad´82, were sowed in field, with the objective to evaluate their behaviour in natural Blast field infection. The absence of dominant genes in heterocigosis involved in the inheritance to Blast was confirmed and, apparently, the behaviour could be governed by major genes in homocigosis and small genes with additive effect. Four isogenic lines wit good agronomic behaviour and Blast resistant were selected. The same will be very useful to obtain new varieties.
Key words: rice, anther culture, disease resistance, genetic inheritance.
INTRODUCCIÓN
El arroz (Oryza sativa L.) es uno de los cereales de mayor producción a nivel mundial y, junto con el trigo y la carne o el pescado, constituye la base de la alimentación humana. El 75 % de la población mundial lo incluye en su dieta alimenticia diaria y puede superar, en algunos casos, el consumo de otros cereales como el maíz y el trigo (1, 2).
En Cuba, representa la principal fuente de carbohidratos en la alimentación de la población, con un consumo aproximado de 670 000 toneladas al año pero, hasta el momento, la producción nacional solo satisface un poco más del 50 % de las necesidades; lo que implica que el país se vea obligado a cubrir mediante importaciones. El rendimiento agrícola promedio se mantiene cercano a las 3 t.ha-1; inferior a la media mundial, lo que es motivado por diferentes causas, entre las que se encuentran: siembras fuera de la época óptima, malas atenciones culturales, disponibilidad de agua limitada, la continua salinización de los suelos y la incidencia de plagas (3).
Entre las últimas se destaca el hongo Magnaporthe grisea Barr (Pyricularia grisea Sacc) que provoca la Piriculariosis, enfermedad considerada como la más devastadora a nivel mundial, debido a su amplia distribución (4). Por otro lado, el mecanismo de resistencia genética a la enfermedad ha sido atribuido a grupos de genes de efectos menores (5) y, en otros casos, a genes dominantes de efectos mayores (6, 7); sin embargo, esta resistencia es superada, debido a cambios y evolución del patógeno que da origen a nuevos haplotipos en el ecosistema, compatibles con los cultivares resistentes.
Teniendo en cuenta estos antecedentes se llevó a cabo el presente trabajo cuyo objetivo fue evaluar el comportamiento de cuatro poblaciones de líneas isogénicas, procedentes del cultivo in vitro de anteras de plantas F2 en su segunda generación, frente a infecciones naturales en campo de P. grisea.
MATERIALES Y MÉTODOS
En el campo experimental de la Unidad Científico Tecnológica de Base «Los Palacios» (UCTB Los Palacios) fueron sembradas a chorrillo en surcos de 5 m de largo semillas de las plantas cosechadas de la primera generación de líneas isogénicas (Tabla I), procedentes del cultivo in vitro de anteras de plantas F2 de los cruces Amistad´82/2077, 2077/Amistad´82, Amistad´82/IR759-54-2-2 y Moroberekan/Amistad´82, junto al progenitor comercial Amistad´82 y los progenitores resistentes a la Piriculariosis 2077, IR759-54-2-2 y Moroberekan. Las labores fitotécnicas se realizaron según el Instructivo Técnico para el cultivo del arroz (8).

Cada línea isogénica (surco) fue evaluada frente a la Piriculariosis por infección natural en campo, en las hojas, durante la fase vegetativa de las plantas, entre 25 y 35 días después de la germinación y en el cuello de la panícula, en la fase reproductiva, de acuerdo con las escalas propuestas por el IRRI (9), las cuales consideran los grados entre 0-3 como resistentes y entre 4-9 como susceptibles, para la evaluación en hojas y entre 5-9 como susceptibles para la evaluación en el cuello de la panícula (Tabla II).

Con los datos obtenidos se realizó una distribución de frecuencias a las poblaciones y la prueba de ajuste de distribución normal según Kolmogorov-Smirnov. Cada clase se correspondió con el valor de la escala de nueve grados empleada para evaluar la respuesta a la enfermedad.
RESULTADOS Y DISCUSIÓN
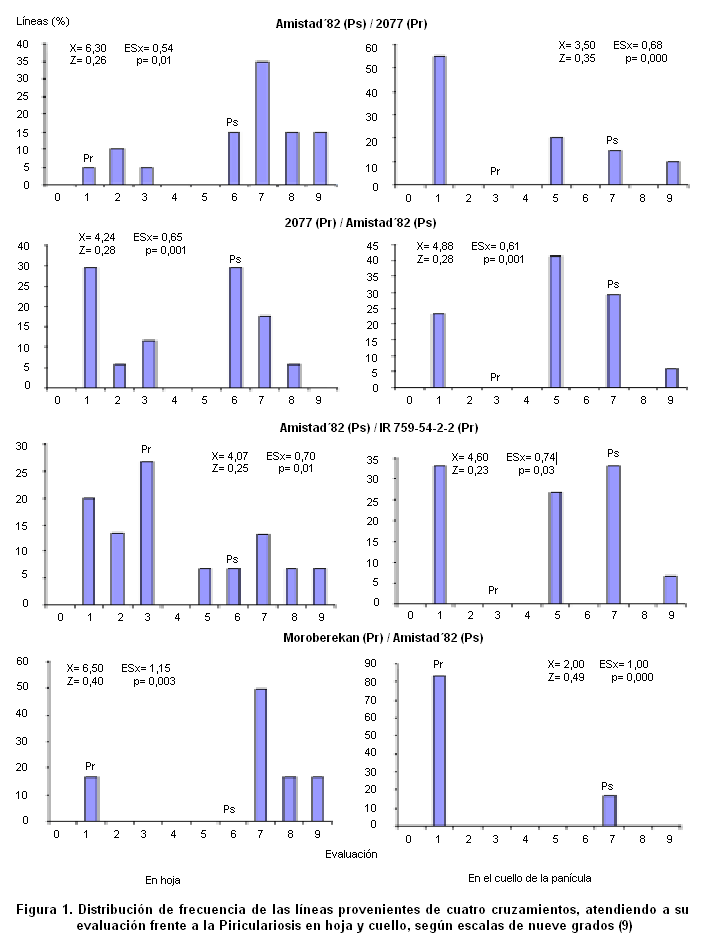
En la Figura 1 se presentan los resultados de la distribución de frecuencias realizadas a los datos del comportamiento frente a la Piriculariosis de las líneas obtenidas procedentes de cuatro poblaciones del progenitor comercial Amistad´82 (susceptible), combinado con tres progenitores resistentes (Amistad´82/2077, 2077/Amistad´82, Amistad´82/IR759-54-2-2 y Moroberekan/Amistad´82).
Teniendo en cuenta que estas líneas fueron obtenidas a partir del cultivo de anteras de plantas F2, constituyen una representación de la segregación en dicha generación, de los cruces que les dieron origen, pudiéndose identificar los recombinantes que regeneraron y constituyeron la base genética para la selección de la resistencia a P. grisea.
En ninguno de los cruzamientos las poblaciones siguieron una distribución normal (p<0,05), según la prueba de Kolmogorov-Smirnov.
No existió solapamiento en el comportamiento de los progenitores, pues se agruparon en clases diferentes y la población de líneas F2 segregó, por su reacción frente a la Piriculariosis, con rangos comprendidos entre las clases uno y nueve.
Es decir las líneas se comportaron desde resistentes hasta susceptibles, pero con ausencia de la clase cuatro y, en algunos casos, también estuvo ausente la clase cinco y otras clases intermedias. De lo anterior se deduce que, dentro de la segregación obtenida en la mayoría de los cruces, no se obtuvieron líneas con una respuesta intermedia a la enfermedad, si se compara con la de los progenitores. El hecho de que no se obtuvieran líneas con un fenotipo intermedio, respecto al de los progenitores, evidencia que ninguna de las líneas fue heterocigótica para el carácter, lo que está en correspondencia con el doblamiento cromosómico del polen haploide que ocurre mediante el cultivo in vitro de anteras, que hace posible la producción de líneas homocigóticas.
En los dos momentos evaluados se presentaron individuos de la progenie con grados susceptibles superiores a los obtenidos por los progenitores y también individuos que originaron clases de resistencia diferentes a la que corresponde al progenitor resistente. Esta aparición en la progenie, de clases nuevas, evidencia los resultados de la recombinación de genes presentes en los progenitores, que dieron lugar a fenotipos nuevos para este carácter o bien a variaciones genéticas que tuvieron su origen en el proceso del cultivo in vitro.
Por otra parte, obtener líneas con una expresión de la resistencia a la piriculariosis, igual o mayor que la del progenitor resistente, indica que en la expresión de la resistencia estuvieron involucrados genes mayores de efecto aditivo, cuya acción fue incrementada, posiblemente por genes menores, también con efecto aditivo, explicándose así los mayores niveles de resistencia que tuvieron algunas de las líneas obtenidas.
En la evaluación de la resistencia a la Piriculariosis en hoja, las poblaciones presentaron una distribución de frecuencias mayor de líneas susceptibles, por ejemplo Amistad´82/2077 y Moroberekan/Amistad´82, o de similar proporción de líneas resistentes y susceptibles (2077/Amistad´82 y Amistad´82/IR 759-54-2-2), lo cual indica que, al parecer, no hay presencia de genes dominantes actuando en la herencia de este carácter.
Todo lo anterior evidencia que el método empleado para la obtención de líneas isogénicas, además de agilizar el proceso, elimina de la segregación aquellos genotipos en heterocigosis y por ello la selección por el fenotipo resistente se hace muy efectiva.
En la evaluación de la resistencia a la Piriculariosis en el cuello de la panícula, se aprecian también poblaciones que presentaron una distribución de frecuencias mayor de líneas susceptibles (2077/Amistad´82 y Amistad´82/IR 759-54-2-2), o de similar proporción de líneas resistentes y susceptibles (Amistad´82/2077), pero apareció además, una población con mayor proporción de líneas resistentes (Moroberekan/Amistad´82).
El comportamiento diferente de las líneas, en los dos momentos evaluados, pudiera estar condicionado por la presencia en el campo de diferentes haplotipos del hongo o deberse a que la resistencia a la enfermedad esté determinada por genes diferentes, de acuerdo a la edad de la planta y al órgano que la misma afecta (hoja o cuello de la panícula).
Teniendo en cuenta la segregación para susceptibilidad en líneas derivadas de plantas resistentes que son heterocigotos para uno o más genes, algunos autores proponen que tres o más genes dominantes deben combinarse en una condición homocigota para conferir resistencia duradera (6).
Otros autores plantean que la resistencia a la Piriculariosis es un carácter de herencia compleja en la que están involucrados, al menos, tres genes mayores independientes en su control (7).
Por su parte otro autor, al evaluar la descendencia de un cruce entre el cultivar ‘Katy’, que posee el gen Pi-ta para resistencia a P. grisea y otro que no lo posee, el mismo no se encontró en los individuos susceptibles y sugiere que se trata de un gen dominante (10). En trabajos posteriores comentó la presencia de este gen en un grupo de cultivares estadounidenses, así como en el cultivar ‘Tetep’ de origen vietnamita, el que posee también los genes Pi-1 y Pi-kh (11). Por su parte, la resistencia a la Piriculariosis en el cultivar ‘Moroberekan’ fue atribuida a la presencia de un grupo de genes mayores en su composición genética (12) y por otro lado se han obtenido líneas resistentes provenientes de cruces donde se emplea el cultivar ‘IR 759-54-2-2’ (13), pero en la literatura consultada no se define que genes posee ni el mecanismo de resistencia que actúa.
Sobre la base de los resultados de la caracterización de las poblaciones del hongo presentes en Uruguay, Brasil y Argentina, se señaló que el alelo de resistencia del gen Pi-ta, presente en los cultivares ‘Yashiro mochi’ y ‘K1’, les confirió incompatibilidad con el linaje A y únicamente la combinación de los genes Pi1, Pi2 y Pi33 otorgan una resistencia completa a todos los linajes presentes en la región (5).
Estudios realizados en el Centro Internacional de Agricultura Tropical de Colombia han demostrado que la combinación de los genes de resistencia Pi-1 (cromosoma 11), Pi-2 (cromosoma seis), y el gen Pi-33 (cromosoma ocho) confieren resistencia a muchas poblaciones de P. grisea en América Latina (5, 14).
El número total de líneas resistentes identificadas en la evaluación efectuada, tanto en la hoja como en el cuello de la panícula, por cruzamientos, se presentan en la Tabla III. Solamente la población proveniente del cruce 2077/Amistad´82, no aportó líneas que combinaron resistencia en la hoja y el cuello de la panícula, aunque fueron obtenidas 12 líneas que mostraron resistencia a la enfermedad cuando esta fue evaluada en uno de los dos momentos.
No obstante, es importante señalar que la obtención de altos rendimientos de los cultivares de arroz está muy asociado con su resistencia a la Piriculariosis en ambos momentos, ya que una plantación que haya escapado a los daños ocasionados por esta enfermedad en su etapa de plántula, puede ser destruida totalmente si se presenta un ataque severo en el cuello de la panícula.
Cuando no hay destrucción total del cultivo es bastante difícil hacer una estimación exacta de las pérdidas, pero se considera que estas son proporcionales al porcentaje del área foliar o del cuello de la panícula afectados (15).
Solo cuatro líneas (A/V-L4, A/I-L11, A/I-L15 y M/A-L6) mostraron resistencia a la Piriculariosis en la hoja y cuello de la panícula, dos de ellas tuvieron como progenitor resistente al cultivar IR 759-54-2-2. Coincide que las cuatro líneas, en un trabajo anterior y con la utilización de un Análisis de Conglomerados (16), fueron ubicadas en la clase donde se lograron los valores más altos de rendimiento y sus componentes principales, comparadas con otras líneas obtenidas por cultivo de anteras y sus progenitores.
Al comparar las reacciones frente a P. grisea en hoja y en el cuello de poblaciones de arroz, desarrolladas por cultivo de anteras y pedigrí, un grupo de autores concluyeron que el cultivo de anteras incrementó la eficiencia de la selección debido a la mayor varianza aditiva, ausencia de dominancia, ausencia de variación intrafamiliar y competencia entre plantas, facilitando la identificación de cultivares superiores con respecto a la selección en generaciones tempranas de un programa de pedigrí, y los valores de heredabilidad sugieren que el avance genético por selección podría lograrse en pocas generaciones (7).
El mejoramiento tradicional ha jugado un papel importante en el incremento del rendimiento y la calidad de los cultivares de arroz en el mundo y la utilización de los métodos biotecnológicos puede asistir el desarrollo de cultivares con rendimiento más alto y resistencia a factores bióticos y abióticos, a la vez que acelera el proceso de obtención de nuevos cultivares debido a la producción rápida de líneas homocigóticas y, por consiguiente, la retención de alelos recesivos o mutaciones útiles. Existen en la actualidad un gran número de cultivares desarrollados combinando las técnicas convencionales de cruzamientos con el cultivo in vitro de anteras (17, 18).
Los resultados de este trabajo han confirmado que, al parecer, no hay presencia de genes dominantes actuando en la herencia de la resistencia a la Piriculariosis y el comportamiento pudiera estar regido por la acción de genes mayores en homocigosis y de genes menores con efecto aditivo. Se logró la selección de cuatro líneas isogénicas (A/V-L4, A/I-L11, A/I-L15 y M/A-L6) resistentes a la Piriculariosis y de buen comportamiento agronómico, las que constituyen la base para la obtención de nuevos cultivares.
REFERENCIAS
1. Álvarez, Rosa M.; Pérez, Mónica; Reyes, Edicta; Moreno, O. J.; Delgado, N.; Torrealba, G. T.; Acevedo, M. A.; Castrillo, W. A.; Navas, María I.; Salazar, Marbelys; Torres, O. J.; Torres, E. A.; García, P. J. y Pérez, A. Evaluación comparativa de híbridos y variedades de arroz en los llanos centroccidentales de Venezuela. Agronomía Tropical, 2008, vol. 58, no. 2, p. 101-110.
2. Méndez, P. Arroz: ¿estabilidad o nueva alza de los precios mundiales? [en línea]. Informativo mensual del mercado mundial del arroz, (84) Febrero 2011. Infoarroz, Centro de Cooperación Internacional en Investigación Agronómica para el Desarrollo (CIRAD). [Consultado: 16 junio 2011]. Disponible en: <http://www.infoarroz.org>.
3. MINAG. Modificaciones al Instructivo Técnico para el cultivo del arroz. Instituto de Investigaciones del Arroz. La Habana, 2011. 30 p.
4. Castejón-Muñoz, M. The effect of temperature and relative humidity on the airbone concentration of Pyricularia oryzae spores and the development of rice blast in southern Spain. Spanish Journal of Agricultural Research, 2008, vol. 6, no. 1, p. 61-69.
5. Livore, A. Desarrollo de una estrategia para la obtención de resistencia durable a Pyricularia grisea en arroz en el cono sur. [en línea]. Informe técnico final-Proyecto FONTAGRO-convenio iica/bid ftg/rf-99-02-rg, INTA. Instituto Interamericano de Cooperación para la Agricultura (IICA). [Consultado: 15 junio 2008]. Disponible en: <http://www.procisur.org.uy/online/DOCS/ pyricularia22.pdf>.
6. Berrio, L. E.; Jenning, P. R.; Torre, E. A. y Cruz, Maribel. Perspectiva del programa de Mejoramiento Varietal del FLAR para la zona tropical", [en línea]. Revista Foro Arrocero Latinoamericano FLAR, [Consultado: 13 julio 2008]. Disponible en: <http://www.flar. org/foroarrocero.htm> .
7. Pérez-Almeida, Iris; Lentini, Zaida y Guimaraes, E. P. El cultivo de anteras en el desarrollo de germoplasma resistente al añublo del arroz (Pyricularia grísea). [en línea]. Fitopatología Venezolana, vol. 8, no. 1, p. 11-14, (1995) [Consultado: 17/01/2009]. Disponible en: <http://www.redpav-fpolar.info.ve/fitopato/v081/ v081f030. htm>.
8. MINAG. Instructivos Técnicos para el cultivo del arroz. Instituto de Investigaciones del Arroz. La Habana, 2008. 115 p.
9. IRRI. Standard Evaluation System for Rice. International Rice Research Institute, 2002. 56 p.
10. Jia, Y. Artificial introgression of a large chromosome fragment around the rice blast resistance gene Pi-ta in backcross progeny and several elite rice cultivars. Heredity, 2009, no. 103, p. 333-339.
11. Séré, Y.; Onasanya, A.; Afolabi, A.; Mignouna, H. D. y Akator, K. Genetic diversity of the blast fungus, Magnaporthe grisea (Hebert) Barr, in Burkina Faso. African Journal of Biotechnology, 2007, vol. 6, no. 22, p. 2568-2577.
12. McNally, K. et al. Sequencing multiple and diverse rice varieties. Connecting whole-genome variation with phenotypes. Plant Physiology, 2006, no. 141, p. 26-31.
13. Hernández, J. Mejoramiento para resistencia a plagas con énfasis en Tagosodes orizicolus, Piricularia grisea y Steneotarsonemus spinki. FAO, 2006
14. Fuentes, J. L.; Correa-Victoria, F. J.; Escobar, F.; Prado, G.; Aricapa, Girlena; Duque, Myriam C. y Tohme, J. Identification of microsatellite markers linked to the blast resistance gene Pi-1(t) in rice. Euphytica, 2008, vol. 160, no. 3, p. 295-304.
15. Zambrano, A.; Vegas, A.; Cardona, R.; Gutiérrez, Z. y Demey, J. R. Estructura genética y diversidad de linajes de Pyricularia grisea en la zona arrocera venezolana. Interciencia, 2006, vol. 31, no. 1, p. 62-66.
16. Pérez, Noraida de J.; González, Maria C.; Castro, R. I. y Aguilar, M. Nuevos genotipos de arroz resistentes a la Piriculariosis obtenidos por cultivo de anteras. Revista Colombiana de Biotecnología, 2012, vol. XIV, no. 1.
17. Chen, C.; Xiao, H.; Zhang, W.; Wang, A.; Xia, Z.; Li, X.; Zhai, W.; Cheng, Z. y Zhu, L. Adapting rice anther culture to gene transformation and RNA interference. Science in China Series C: Life Sciences, 2006, vol. 49, no. 5, p. 414-428.
18. Silva, T. D. Indica rice anther culture:can the impasse be surpassed?. Plant Cell Tissue Organ Culture, 2010, vol. 100, p. 1-11.
Recibido: 21 de febrero de 2012
Aceptado: 30 de agosto de 2012
Noraida de J. Pérez León, Investigador Auxiliar de la Unidad Científico Tecnológica de Base "Los Palacios". Email: nory@inca.edu.cu

{kind=link}
{kind=link}