










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Medicina Tropical
versión On-line ISSN 1561-3054
Rev Cubana Med Trop v.52 n.2 Ciudad de la Habana mayo-ago. 2000
CENTRO DE INVESTIGACIONES CIENTÍFICAS DE LA DEFENSA CIVIL
Anticuerpos neutralizantes frente a 5 cepas de VIH-1 en macacos inmunizados con el polipéptido multiepitópico TAB9
Dra. Leonor M. Navea Leyva,1 Dra. Leonor Lobaina Bartelemy,1 Lic. Marta Dubed Echevarría,2 Lic. Giselle Álvarez Seguí,3 Lic. Carmen Gómez Rodríguez4 y Dr. Carlos Duarte Cano5
RESUMEN
Se estudió y comparó la presencia de anticuerpos neutralizantes (AcN) frente a 5 cepas de laboratorio del virus de la inmunodeficiencia humana tipo 1 (VIH-1) en el suero de 8 macacos (Macaca fascicularis) después de la tercera y cuarta inmunización con el polipéptido multiepitópico TAB9, emulsificado con el adyuvante Montanide ISA 720; 4 animales se inocularon con 1 mg y los otros con 200 µg, como control se inyectaron 2 animales sólo con el adyuvante. Aunque se comprobó la presencia de anticuerpos neutratizantes en la mayoría de los animales, después de las inmunizaciones frente a cepas homólogas del grupo B del VIH-1 por el método ELISA de captura de antígeno p24 (DAVIH-Ag p24, Cuba), no se detectaron diferencias estadísticamente significativas en los títulos provocados por las concentraciones de antígeno ni en las respuestas después de la tercera y cuarta dosis inmunizante. Los animales controles no desarrollaron anticuerpos neutralizantes.
Descriptores DeCS: VIH-1/inmunologia;VIH-1/sangre; ANTICUERPOS VIRALES/inmunologia; ANTICUERPOS VIH/inmunología; PEPTIDOS; EPITOPES;MACACA; INMUNIZACION PASIVA.
La pandemia del síndrome de inmunodeficiencia adquirida (SIDA) ha impulsado la necesidad urgente del desarrollo de vacunas para que constituyan la estrategia preventiva más importante contra el virus de inmunodeficiencia humana (VIH), por lo que se llevan a cabo grandes esfuerzos para obtener un preparado vacunal seguro y eficaz, tanto en Cuba como en otros países. En los últimos años, se han estudiado varios candidatos en modelos animales y humanos.1,2 En algunos experimentos en macacos y chimpancés se ha observado protección contra el reto viral.3 La mayoría de estos biopreparados se basan en la producción de anticuerpos neutralizantes (AcN) con el objetivo de que éstos puedan ejercer un papel importante en la protección; aunque tienen la desventaja de que, en aquellas vacunas conformadas con las secuencias del lazo V3 de la gp 120, estos anticuerpos son cepa-específicos y por lo tanto no son capaces de neutralizar aislamiento de VIH-1 divergentes.4,5
Una de las estrategias vacunales empleadas para buscar respuesta de AcN con mayor espectro es el desarrollo de polipéptidos multiepitópicos (PME) del VIH-1.6,7 Estas son proteínas recombinantes quiméricas que expresan la región del lazo V3 de la gp120 de diferentes aislamientos del VIH-1. El PME TAB9 que contiene 6 regiones V3 provoca una potente respuesta de anticuerpos en conejos y ratones, cuando se usa en combinación con el adyuvante Montanide ISA 720 (M-ISA 720) (Quintana D. Centro de Ingeniería Genética y Biotecnología [comunicación personal].1997).
En este trabajo se determinó y comparó la presencia de AcN frente a 5 cepas de referencia del VIH-1 en sueros de macacos (Macaca fascicularis) después de la tercera y cuarta dosis de inmunizaciones con el PME TAB 9.
MÉTODOS
Cepas virales de laboratorio: Se emplearon 5 cepas VIH-1 para la realización del estudio: MN, LAI, RF, y III-B que se obtuvieron por una donación del Medical Research Council -MRC- AIDS Reagent Program (Reino Unido de la Gran Bretaña -RUGB-) y la cepa 92UG046, del Programa de la OMS para la lucha contra el SIDA (ONUSIDA). Esta última presenta una gran homología con el aislamiento JY1, aunque en la cepa 92UG046m, adaptada en nuestro laboratorio a la línea celular MT4, se encontraron cambios en 3 aminoácidos dentro de la región V3 (Quintana D. Centro de Ingeniería Genética y Biotecnología [comunicación personal].1997).
PME TAB9: El PME TAB9 contiene la secuencia central de 15 aminoácidos correspondiente a 6 aislamientos de VIH-1 (LR150, JY1, RF, MN, BRVA y IIIB). Se obtuvo en la División de Desarrollo de Procesos y se unió al adyuvante MI-ISA 720 en la División de Formulaciones, ambas dependencias del Centro de Ingeniería Genética y Biotecnología (CIGB).
Sueros hiperinmunes: Se inmunizaron 8 macacos con la tercera y cuarta dosis de 1 mL de PME TAB9/M-ISA 720, según un esquema que aplicó el inmunógeno en los meses 0, 1, 6 y 12. Se inocularon 4 animales con 1 mg (sueros 1-4) y los otros con 200 µg (sueros 5-8). Además, 2 macacos se inmunizaron solamente con el adyuvante M-ISA 720 (sueros 9 y 10). Un mes después de concluido el esquema de inmunización se obtuvieron muestras de sangre por punción venosa, los sueros se filtraron a través de membranas de acetato de celulosa de 0,2 µm (Nalgene, EE. UU.) y se realizaron diluciones seriadas de 1:20-1:320 en medio RPMI-1640 con 10 % de suero fetal bovino (SFB) (Gibco Ltd., RUGB).
Ensayo de neutralización: Se emplearon placas de 96 cavidades donde se mezclaron 50 µL de las diferentes diluciones del suero con igual volumen de cada uno de los aislamientos virales (100 TCID50), por separado. Después de incubar durante 1 h a 37 °C, se adicionaron 100 µL de células MT4 (1 X 105) en medio RPMI-1640 con 10 % de SFB y antibióticos (penicilina: 100 UI/mL - estreptomicina: 100 µg/mL). A las 24 h y al cuarto día se efectuó cambio de medio, con la adición de 150 µL y 100 µL de medio fresco, respectivamente. Al séptimo día se midió la multiplicidad viral mediante un sistema inmunoenzimático (ELISA) de captura de antígeno p24 (DAVIH-Ag) p24) (DAVIHLab., Cuba) en los sobrenadantes de cultivo. Los experimentos se repitieron por triplicado. Se utilizaron los controles siguientes: celulas (células solamente), viral (células y virus), positivo (anticuerpo monoclonal -AcM- con actividad neutralizante conocida) y negativo (suero conocido sin actividad neutralizante). El título de AcN se determinó como el inverso de la mayor dilución donde se produjo una inhibición de 50 ó 90 % del Ag p24.
Análisis estadístico: Para comparar los títulos de AcN después de la tercera y cuarta inoculación y entre la dosis máxima y la dosis mínima, se aplicó una prueba t de Student para los datos transformados; para la comparación de los títulos de AcN frente a las diferentes cepas se realizó un análisis de varianza ANOVA para ambas inmunizaciones; siempre se consideró significativa una diferencia cuando p ?_0,05.
RESULTADOS
Todos los animales inoculados con el PME TAB9//M-ISA 720, tanto con 200 µg como con 1 mg, desarrollaron AcN contra los aislamientos LAI, IIIB, MN y RF de VIH-1, después de la tercera inmunización; sin embargo, frente a la cepa 92UG046m sólo 2 sueros (5 y 7) reaccionaron en respuesta a la dosis de 200 µg. Cuando se comparó la respuesta para una neutralización de 50 % frente a todas las cepas, los títulos de AcN fueron significativamente menor frente a la cepa 92UG046m (p < 0,05), pero no hubo diferencias estadísticas significativas para una neutralización de 90 % (tabla 1).
TABLA 1. Títulos de anticuerpos neutralizantes de sueros de macacos frente a cepas de referencia del VIH-1 un mes después de la tercera dosis
| Neutralización de cepas de laboratorio de VIH-1 | |||||||||||
| Dosis PME TAB9 | Suero | LAI | IIIB | MN | 92UG046m | RF | |||||
| No. | 50 % | 90 % | 50 % | 90 % | 50 % | 90 % | 50 % | 90 % | 50 % | 90 % | |
|
| 1 | 320 | < 20 | 284 | 71 | 99 | 25 | < 20 | < 20 | 320 | 320 |
| 1 mg | 2 | 320 | 48 | 320 | < 20 | 157 | 39 | < 20 | < 20 | 320 | 320 |
|
| 3 | 320 | 24 | 120 | < 20 | 50 | < 20 | < 20 | < 20 | < 20 | < 20 |
|
| 4 | 32 | < 20 | 196 | < 20 | < 20 | 320 | < 20 | < 20 | < 20 | < 20 |
| TPG |
| 179 | 12 | 215 | 13 | 62 | 27 | - | - | 320 | 320 |
| F |
|
| 4/4 | 2/4 | 4/4 | 1/4 | 3/4 | 0/4 | 0/4 | 2/4 | 2/4 |
|
| 5 | 320 | < 20 | 320 | <20 | < 20 | 320 | 160 | < 20 | 320 | 40 |
| 200 µg | 6 | 320 | < 20 | < 20 | 320 | < 20 | 320 | < 20 | < 20 | 80 | < 20 |
|
| 7 | 211 | 26 | 195 | 24 | 320 | 81 | 94 | 24 | 320 | 320 |
|
| 8 | 24 | 320 | 27 | 320 | 76 | < 20 | < 20 | < 20 | 40 | < 20 |
| TPG |
| 144 | 8 | 76 | 22 | 78 | 80 | 67 | 22 | 100 | 23 |
| F |
| 4/4 | 2/4 | 3/4 | 3/4 | 2/4 | 3/4 | 2/4 | 1/4 | 4/4 | 2/4 |
| Adyu- | 9 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 |
| vante | 10 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 |
< 20: no hubo neutralización; TPG: título promedio geométrico; F: frecuencia de neutralización.
Por otra parte, después de la cuarta inmunización, también se encontraron títulos neutralizantes en todos los animales frente a las cepas LAI, IIIB, MN y RF; pero frente a la cepa 92UG046m sólo se identificaron 4 sueros reactivos, 2 en la dosis mayor y 2 en la dosis menor. Sin embargo, no hubo diferencias significativas para la actividad neutralizante ni de 50 ni de 90 % (tabla 2).
TABLA 2. Títulos de anticuerpos neutralizantes de sueros de macacos frente a cepas de referencia del VIH-1 un mes después de la cuarta dosis
| Neutralización de cepas de laboratorio de VIH-1 | |||||||||||
| Dosis PME TAB9 | Suero | LAI | IIIB | MN | 92UG046m | RF | |||||
| No. | 50 % | 90 % | 50 % | 90 % | 50 % | 90 % | 50 % | 90 % | 50 % | 90 % | |
|
| 1 | 180 | 28 | 320 | 320 | 104 | 56 | < 320 | 207 | 333 | 104 |
| 1 mg | 2 | 372 | 225 | 320 | 320 | < 160 | 110 | < 20 | < 20 | 422 | 226 |
|
| 3 | 167 | 27 | 431 | 225 | < 20 | < 20 | < 20 | < 20 | 53 | < 20 |
|
| 4 | < 20 | < 20 | 47 | 28 | 20 | < 20 | 46 | < 20 | < 20 | < 20 |
| TPG |
| 46 | 11 | 211 | 57 | 23 | 8 | 10 | 16 | 51 | 12 |
| F |
| 3/4 | 3/4 | 4/4 | 4/4 | 3/4 | 2/4 | 2/4 | 1/4 | 3/4 | 2/4 |
|
| 5 | 431 | 225 | 417 | 226 | 23 | < 20 | <20 | < 20 | < 20 | < 20 |
| 200 µg | 6 | 43 | < 20 | 410 | 56 | < 20 | < 20 | 26 | < 20 | < 20 | < 20 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
| 7 | < 20 | < 20 | 50 | 28 | 171 | 57 | 204 | 28 | 417 | 208 |
|
| 8 | < 20 | < 20 | 104 | 57 | 20 | < 20 | < 20 | < 20 | 24 | < 20 |
| TPG |
| 20 | 17 | 170 | 66 | 16 | 12 | 8 | 10 | 17 | 16 |
| F |
| 2/4 | 1/4 | 4/4 | 4/4 | 3/4 | 1/4 | 2/4 | 1/4 | 2/4 | 1/4 |
| Adyu- | 9 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 |
| vante | 10 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 | < 20 |
< 20: no hubo neutralización; TPG: título promedio geométrico; F: frecuencia de neutralización.
Cuando se compararon los títulos de AcN provocados por el inóculo de 200 µg y 1 mg para cada cepa viral, no se encontraron diferencias estadísticamente significativas en la respuesta a ambas dosis, tanto después de la tercera como de la cuarta inmunización. Los sueros de los macacos inoculados sólo con el adyuvante nunca desarrollaron AcN.
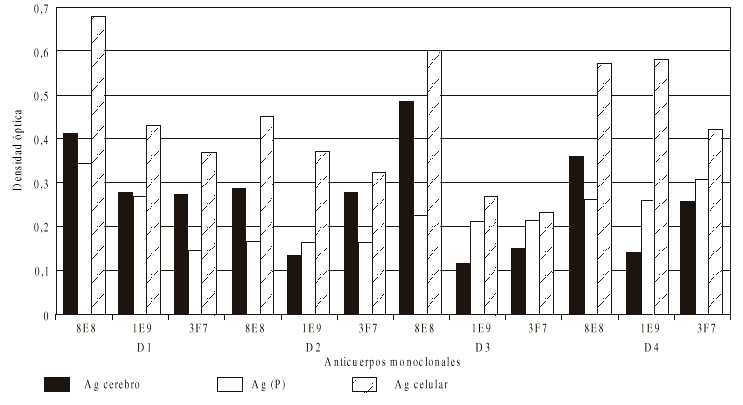
Fig. Patrón de reactividad de los anticuerpos monoclonales (AcMs) por Elisa-da.
DISCUSIÓN
Aun cuando no se deben hacer comparaciones entre las diferentes preparaciones virales por la conocida complejidad biológica que dificulta la normalización, estos resultados permiten evaluar la inmunogenicidad general del TAB9 en macacos. Ambas concentraciones (200 µg y 1 mg) resultaron efectivas para la inducción de AcN en la mayoría de los animales frente a cepas del grupo B (LAI, IIIB, MN y RF), todas representadas en el PME. La menor respuesta frente a la cepa 92UG046m se podría explicar por las diferencias genéticas entre ésta y los epitopes de la cepa JY1 representada en el TAB9. Otros autores que obtuvieron AcM contra VIH-1 coinciden en el predominio de actividad neutralizante cepa-específica frente a las cepas homólogas.8-10
Se confirmó que después de la tercera y de la cuarta inmunización con TAB9, este PME fue capaz de inducir AcN contra cepas de laboratorio de VIH-1. Es importante estudiar la actividad neutralizante frente a aislamientos primarios de VIH-1, obtenidos a partir de cocultivos de células mononucleares de sangre periférica de pacientes o portadores del virus, pues los candidatos vacunales desarrollados hasta la fecha no han sido capaces de inducir este tipo de anticuerpos.1 Por esa razón, la relevancia que podría tener este ensayo como indicador de protección en los humanos es objeto de intensa discusión y se requieren otros experimentos para dilucidarla.
No obstante, la actividad neutralizante depende de varios factores: el adyuvante, la presentación del antígeno, la combinación de epitopes y el ajuste de dosis inoculadas en el esquema de inmunización. Si la calidad de la respuesta de AcN se evalúa por la especificidad neutralizante de éstos, una mejor comprensión del mecanismo de la neutralización permitirá modificar la formulación del candidato vacunal para lograr una actividad neutralizante óptima contra el virus. El mejoramiento de los ensayos biológicos para evaluar la actividad neutralizante, junto a otras que evalúen la inmunidad celular inducida por el candidato, permitiría en el futuro la selección de los mejores candidatos vacunales para el VIH-1.
SUMMARY
Presence of neutralizing antibodies(Nab) to 5 laboratory strains of human immunodeficiency virus type-1 (HIV-1) was studied and compared in the sera of eight macaca (Macaca fascicularis) after the third and fourth immunizations with multi-epitope polypeptide TAB9, emulsified with adjuvant Montanide ISA 720. Four animals were innoculated 1mg and the others 200 µg, 2 animals were used as controls and injected only with adjuvant. Although the presence of neutralizing antibodies to homologous strains of HIV-group group B was confirmed in most animals after immunization through antigen p24 capture ELISA (DAVIH-Agp24, Cuba), no statistically significant differences were found neither in titers caused by antigen concentrations nor in the responses after the third and fourth immunizing dosis. Controls did not develop neutralizing antibodies.
Subject headings: HIV-1/immunology; HIV-1 blood; ANTIBODIES, VIRAL/immunology; HIV ANTIBODIES/immunology; PEPTIDES; EPITOPES; MACACA; IMMUNIZATION, PASSIVE.
REFERENCIAS BIBLIOGRÁFICAS
- Esparza J, Heyward WL, Osmanov S. HIV Vaccine development: from basic research to human trials. A/DS 1996;10(Suppl A): S123-S132.
- Emini E, Nara PO, Schlif W A, Lewis JA, Davide JP, Lee DR, et.al. Antibody mediated in vitro neutralization of human immunodeficiency virus type l abolishes infectivity for chimpanzees. J Virol 1990;64(8):3674-8.
- Girard M, Kieny MP, Pinter A, Barre-Sinoussi F, Nara P, Kolbe H, et al. Immunization of chimpanzees confers protection against challenge with human immunodeficiency virus. Proc Nat Acad SciUSA 1991;88:542-6.
- Letvin NL. Progress in the development of an HIV-1 vaccine. Science 1998;280:1875-80.
- Hellman CA, Baltimore DHW. Vacine-where are we going?. Nature Med Vaccine 1998;4(Suppl 5): 532-4.
- Duarte CA, Montero M, Seralena A, Valdés R, Jiménez V, Benítez J, et al. Multi-epitope polypeptide (MEP) containing epitopes of HIV-1 envelope induces neutralizing monoclonal antibodies against the V3 loop.AIDS Res Hum Retroviruses 1994;10(3):235-43.
- Montero M, Menéndez A, Domínguez MC, Navea L, Vilarrubia OL, Quintana D, et al . Broadly reactive antibodies against a gp 120 V3-loop multi-epitope polypeptide neutralize different isolates on human immunodeficiency virus type 1 (HIV-1). Vaccine 1997;15/(11):1200-8.
- Steinman RM, Germain RM. Antigen presentation and related immunological aspects of HIV-1 vaccines. AIDS 1998;12(Suppl A): S97-S112.
- Gauduin MC, Parren PWHI, Weir R, Barbas CF, Burton DR, Koup RA. Passive immunization with a potent neutralizing human monoclonal antibody HU-PBL-SCID mice against challenge by primary isolates of human immunodeficiency virus type 1. Nature Med 1997;3:1389-93.
- Kostrikis LG, Cao Y, Ngai H, Moore JP, Ho DD. Quantitative analysis of serum neutralization of human immunodeficiency virus type 1 from subtypes A,B,C,D,E,F and l: lack of direct correlation between neutralization serotypes and genetic subtypes and evidence for prevalent serum-dependent infectivity enhancement. J Virol 1996;70 (1):445-58.
Recibido: 2 de agosto de 1999. Aprobado: 16 de marzo del 2000.
Dra. Leonor M. Navea Leyva. Centro de Investigaciones Científicas de la Defensa Civil. Carretera de Tapaste y Ocho Vías. San José de las Lajas, La Habana, Cuba.
1 Especialista de I Grado en Microbiología. Investigadora Auxiliar.
2 Máster en Virología. Licenciada en Ciencias Biológicas. Investigadora Auxiliar.
3 Licenciada en Ciencias Biológicas. Investigadora Agregada.
4 Licenciada en Bioquímica. Aspirante a Investigadora.
5 Doctor en Ciencias Biológicas. Investigador Auxiliar.

