










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Medicina Tropical
versión On-line ISSN 1561-3054
Rev Cubana Med Trop v.63 n.1 Ciudad de la Habana ene.-abr. 2011
ARTÍCULO ORIGINAL
Estrategia cubana de caracterización molecular del virus influenza A/H1N1pdm
Cuban strategy for the molecular characterization of the pandemic influenza A virus (H1N1)
Alexander Piñón Ramos,I Belsy Acosta Herrera,II Odalys Valdés Ramírez,III Amely Arencibia García,IV Clara Estela Savón Valdés,V Grehete González Muñoz,VI Suset Isabel Oropesa Fernández,VII Elías Quilarte García,VIII Guelsys González Baez,IX Bárbara Hernández Espinosa,X Ángel Goyenechea Hernández,XI María Guadalupe Guzmán Tirado,XII Alina Llop Hernández,XIII Vivian Kourí CardelláXIV
I Licenciado en Microbiología. Máster en Virología. Investigador Agregado. Instructor. Departamento de Virología, Instituto de Medicina Tropical "Pedro Kourí" (IPK). La Habana, Cuba.
II Doctor en Medicina. Especialista de II Grado en Microbiología. Investigador Auxiliar. Asistente. La Habana, Cuba.
III Licenciada en Bioquímica. Doctora en Ciencias de la Salud. Investigadora Titular. Asistente. Departamento de Virología, IPK. La Habana, Cuba.
IV Licenciada en Microbiología. Reserva Científica. Departamento de Virología, IPK. La Habana, Cuba.
V Licenciada en Ciencias Biológicas. Doctora en Ciencias Biológicas. Investigadora Titular. Profesora Titular. Departamento de Virología, IPK. La Habana, Cuba.
VI Licenciada en Microbiología. Máster en Virología. Investigadora Auxiliar. Asistente. Departamento de Virología, IPK. La Habana, Cuba.
VII Doctor en Medicina. Médico Especialista de II Grado en Microbiología. Investigador Auxiliar. Profesor Auxiliar. Departamento de Virología, IPK. La Habana, Cuba.
VIII Doctora en Medicina. Especialista de I Grado en Medicina General Integral. Residente de Microbiología, IPK. La Habana, Cuba.
IX Técnico Medio en Química Farmacéutica. Auxiliar Técnico Docente. Departamento de Virología, IPK. La Habana, Cuba.
X Técnico Medio en Microbiología. Auxiliar Técnico Docente. Departamento de Virología, IPK. La Habana, Cuba.
XI Especialista de II Grado en Microbiología. Investigador Titular. Profesor Titular. Departamento de Virología, IPK. La Habana, Cuba.
XII Doctora en Medicina. Doctora en Ciencias Médicas. Doctora en Ciencias. Investigadora Titular. Profesora Titular. Departamento de Virología, IPK. La Habana, Cuba.
XIII Doctora en Medicina. Doctora en Ciencias Médicas. Investigadora Titular. Profesora Titular. Subdirección de Microbiología, IPK. La Habana, Cuba.
XIV Doctora en Medicina. Doctora en Ciencias Médicas. Investigadora Titular. Profesora Auxiliar. Departamento de Virología, IPK. La Habana, Cuba.
RESUMEN
INTRODUCCIÓN: en Abril de 2009 se identificó una variante del virus influenza A/H1N1 de origen porcino, lo cual determinó que fuese declarada rápidamente la primera pandemia del siglo XXI.
OBJETIVO: establecer una estrategia de secuenciación nucleotídica que permitiera diagnosticar diferencialmente los virus influenza A estacionales del nuevo virus pandémico, así como obtener la mayor cantidad de información posible desde el punto de vista molecular de los genes hemaglutinina y neuraminidasa, tanto de pacientes que sufrieron una enfermedad tipo influenza como los que padecieron de una infección respiratoria aguda grave y los que fallecieron.
MÉTODOS: se diseñaron e implementaron tres estrategias de secuenciación que brindaron información importante acerca del nuevo virus en Cuba.
RESULTADOS: a través de la tercera estrategia se obtuvieron los resultados más completos: diagnóstico diferencial, vigilancia de las mutaciones D222G/E en la hemaglutinina y las variantes virales H275Y resistentes al Tamiflu. A pesar de no haber detectado las mutaciones mencionadas, no se puede descartar su presencia en población cubana, debido a que estas estrategias no fueron diseñadas con ese fin. Se impone diseñar un estudio para cumplir con ese objetivo.
CONCLUSIONES: las estrategias de secuenciación aplicadas en nuestro algoritmo permitieron realizar el diagnóstico diferencial de los virus influenza estacional del pandémico y su caracterización molecular.
Palabras clave: influenza pandémica, secuenciación, caracterización molecular, Cuba.
ABSTRACT
INTRODUCTION: in April 2009, there was identified a variant of the A/H1N1 influenza virus of swine origin, and shortly after the first pandemic in XXI century was declared.
OBJECTIVES: to establish a nucleotide sequencing strategy for the differential diagnosis of the seasonal and pandemic influenza A viruses, and to obtain as much molecular information as possible about hemagglutinin and neuraminidase genes in patients with influenza-like illnesses, in those with severe respiratory infection and in patients who died.
METHODS: three sequencing strategies were designed and implemented, which also offered important information about the new virus in Cuba.
RESULTS: the third strategy provided the most comprehensive results such as differential diagnosis, the surveillance of the D222G/E mutation in hemagglutinin and Tamiflu-resistant H275Y viral variants. In spite of the fact that the mentioned mutations were not detected, their presence in the Cuban population can not be ignored since these strategies were not designed for this end. It is imperative to design a study to fulfill this objective.
CONCLUSIONS: the sequencing strategies in our algorithm allowed the differential diagnosis of the seasonal and the pandemic viruses, and their molecular characterization.
Key words: pandemic influenza, sequencing, molecular characterization, Cuba.
INTRODUCCIÓN
En abril de 2009 fue identificada una variante del virus influenza A/H1N1 de origen porcino conocida como la «influenza mexicana», la que fue declarada rápidamente como la primera pandemia del siglo XXI. Muchos países del mundo tenían sus reservas de drogas antivirales para el tratamiento de los pacientes infectados y para minimizar la diseminación del virus durante la primera fase de la pandemia, período en el cual no hay disponibilidad de vacunas efectivas contra la cepa pandémica.1 La detección rápida de variantes de la influenza A/H1N1 pandémica (A/H1N1pdm) y la identificación de variantes resistentes a los antivirales es de gran importancia para la vigilancia pandémica y en el control de casos.
Las variaciones genéticas, predominantemente en la forma de mutaciones puntuales, son una propiedad inherente de los virus influenza y otros virus cuyo genoma se compone de ácido ribonucleico (ARN). La mayoría de las sustituciones de aminoácidos no están ligadas a alteraciones en las propiedades del virus y las variantes virales que contienen tales sustituciones son descritas como variantes neutrales. Su significación (si es que la tienen), permanece desconocida. Sin embargo, en algunas ocasiones pueden emerger variantes con algunas de sus propiedades alteradas, como pueden ser el rango de hospedero, tropismo, potencial de replicación, antigenicidad, entre otras.2
A través de la Red de Vigilancia Global de Influenza (RVGI)3 rectorada por la Organización Mundial de la Salud (OMS), varios países han logrado la secuenciación nucleotídica de los virus A/H1N1pdm. Gran parte de este trabajo se centra en los resultados de la caracterización molecular de regiones del genoma de los virus influenza que determinan alteraciones en las propiedades antigénicas y perfiles de susceptibilidad a las drogas. Los cambios antigénicos pueden alterar la efectividad de las vacunas actuales que obligan a realizar selecciones y recomendaciones de nuevos virus para la composición de esta. Por otro lado, hay cambios que pueden provocar la aparición de variantes resistentes a las drogas anti-influenza disponibles, que restringen las opciones de tratamiento específico y su uso como medida de control y prevención. La mayoría de los virus A/H1N1 pdm son resistentes a los antivirales conocidos como adamantanos, sin embargo, solo existen reportes de algunos casos individuales y pequeños grupos de virus pandémicos resistentes al oseltamivir (inhibidor de la neuraminidasa= INA). Todos los virus oseltamivir-resistentes son susceptibles a otra alternativa de INA, el zanamivir. Hasta la fecha no han emergido virus que hayan experimentado cambios antigénicos de interés en comparación con la cepa de referencia A/California/7/2009 y sus similares.4
Los virus influenza inician el proceso de infección a través de la unión de la hemaglutinina (HA) a las moléculas de azúcar del ácido siálico (receptores) ubicadas en la superficie de las células hospederas. La infección por el virus influenza es generalmente restringida al tracto respiratorio superior (TRS), localización donde predominan los receptores de ácido siálico de unión a2-6.5 Existen varios reportes de cambios a nivel molecular que afectan la unión a los receptores de la superficie celular a la cual van a infectar. La variante D222G ha sido descrita en la HA de un número limitado de virus A/H1N1 pdm, y se ha sugerido la posibilidad de que afecte la unión de la HA a su receptor, pudiendo ser la responsable de un cambio en la preferencia de unión al receptor de ácido siálico de a2-6 a una preferencia dual a2-3/a2-6.6 Las células del tracto respiratorio que expresan receptores de unión a2-3 son más abundantes en el tracto respiratorio inferior (TRI) de las vías respiratorias del humano.7 Esta observación ha llevado a la hipótesis de que los virus con especificidad de receptor dual pueden replicarse y producir títulos virales elevados en los pulmones, que dan como resultado un curso más severo de la enfermedad. Sin embargo, no han sido publicados datos experimentales con cepas del virus nuevo y recientes estudios con el virus pandémico A/H1N1 de 1918 no apoyan esta hipótesis.8
Todas las razones expuestas antes apoyan la importancia de la vigilancia de la evolución molecular de estos virus. Teniendo en cuenta que las secuencias nucleotídicas generan cantidades enormes de información sobre la variación de los virus influenza y que constantemente son descritas mutaciones nuevas, se considera esta como una vía importante de vigilar las huellas moleculares de estos virus y su posible impacto en la salud humana.
En Cuba se diagnosticaron los primeros casos en el Centro Nacional de Influenza (CNI), ubicado en el Instituto de Medicina Tropical "Pedro Kourí" (IPK), desde el mismo inicio de la pandemia. Sin embargo, además de diseñar y aplicar una estrategia diagnóstica específica para el nuevo virus, fue necesaria la implementación de una estrategia que permitiera la caracterización y vigilancia molecular del virus nuevo A/H1N1 pdm.
MÉTODOS
Muestras
Las muestras que formaron parte de este estudio resultaron ser muestras positivas al virus influenza A, tanto por un protocolo TR-RCP múltiple,9 como por el protocolo publicado por el Centro de Control de Enfermedades de Atlanta (CDC de Atlanta).10
Con la finalidad de obtener una elevada eficiencia en la amplificación de los segmentos de los genes deseados, de las muestras positivas solo fueron seleccionadas aquellas cuyos valores de ciclo (a partir del cual la intensidad de la fluorescencia emitida sobrepasa la basal [CT]) eran menores que 30. Además, las muestras positivas al virus pandémico pertenecían a diferentes grupos de pacientes: los que evolucionaron satisfactoriamente después de presentar una enfermedad tipo influenza (ETI), los que evolucionaron padeciendo una infección respiratoria aguda grave (IRAG), y los que evolucionaron tórpidamente hasta su fallecimiento.
Obtención de los productos de los genes de la hemaglutinina y la neuraminidasa
Con la finalidad de obtener fragmentos de calidad y de gran tamaño para la secuenciación nucleotídica, se trazaron varias estrategias de secuenciación que se fueron perfeccionando en el transcurso de la pandemia. Las tres estrategias consistieron en:
1. Utilización de un protocolo TR-RCP anidada del Instituto de Salud Carlos III, Majadahonda, España11 para secuenciar un segmento del gen de la HA de aproximadamente unos 1 000 nucleótidos de los virus influenza A/H1N1 e influenza A/H3N2.
2. Introducción del protocolo de TR-RCP empleado por el Centro de Referencia de la OMS en Londres para secuenciar los genes completos de la HA y la neuraminidasa (NA).12
3. Combinación de ambos métodos con la finalidad de obtener productos de mejor calidad para secuenciar la mayor longitud posible de los genes de la HA y la NA, realizado por nuestro laboratorio.
Estrategia 1
Transcripción reversa-reacción en cadena de la polimerasa (TR-RCP)
La TR-RCP se realizó utilizando el estuche comercial OneStep RT-PCR siguiendo las instrucciones del productor (catálogo 210212, QIAGEN, Alemania). Brevemente, se añadieron 5 µL del ARN extraído de las muestras de exudado nasofaríngeo a la mezcla de reacción (de volumen final 45 µL) que contenían 30 µL de agua libre de ARNasa, 10 µL de solución tampón contenida en el estuche comercial, 2 µL dNTPs (10 mM cada uno), 0,5 µL de cada cebador descritos antes por Tenorio y otros11 a 10 µM cada uno y 2 µL del reactivo EMix contenido en el estuche comercial. Los cebadores utilizados en este paso permiten amplificar la HA de los virus influenza H1N1 estacional y pandémico, así como la HA de los virus estacionales H3N2.
Reacción en cadena de la polimerasa anidada (RCP anidada)
La RCP anidada se realizó utilizando la enzima Amplitaq DNA polimerasa (catálogo F00759, Applied Biosystem, USA). En esta reacción se utilizan 2 µL del producto obtenido en la reacción de TR-RCP en las dos mezclas que se preparan por separado. Estas dos mezclas permiten la amplificación por separado de un segmento de la HA de los virus H1N1 estacional y pandémico por un lado y del virus H3N2 por otro. Los detalles de las mezclas se muestran en la tabla, los cebadores utilizados fueron anteriormente descritos.11
Las longitudes de los segmentos obtenidos de estas reacciones son de 980 pb para el virus influenza A/H1N1 y 1 100 pb para el virus influenza A/H3N2.
Estrategia 2
Transcripción reversa-reacción en cadena de la polimerasa (TR-RCP)
La TR-RCP se realizó según las indicaciones del Centro de Referencia de la OMS de Londres y las normas para el diagnóstico de laboratorio publicadas por la OMS. Los cebadores utilizados en este protocolo permiten amplificar la HA y NA de los virus influenza A/H1N1 estacional y pandémico, así como de los virus estacionales del subtipo A/H3N2.12 Las combinaciones de los cebadores permiten obtener productos que se superponen, lo cual garantiza obtener mayor información y secuencias de mayor confiabilidad.
Estrategia 3
Para esta estrategia se utilizó el producto obtenido en la reacción de TR-RCP de la estrategia 1. Las mezclas de la RCP anidada fueron realizadas con combinaciones diferentes de los cebadores de la estrategia 2, que contenían 31,5 µL de agua libre de ARNasas, 10 µL de solución tampón, 2 µL de dNTPs (10 mM cada uno), 2 µL de cada cebador a 10 µM cada uno y 2 µL de la enzima Amplitaq DNA polimerasa (catálogo F00759, Applied Biosystem, USA).
Purificación de los productos de reacción en cadena de la polimerasa obtenidos
Los productos se purificaron utilizando el estuche comercial QIAquick PCR Purification (catálogo 28106, QIAGEN, Alemania) siguiendo las especificaciones del productor. El proceso se realizó de forma automatizada con la utilización del QIAcube (QIAGEN, Alemania), mediante las instrucciones del productor.
Reacciones de secuencia y purificación
Las reacciones de secuencia y la purificación se realizaron utilizando el estuche comercial Dye Terminator Cycle Sequencing (DTCS) Quick Start (catálogo PN 608120, Beckman Coulter, USA), siguiendo las indicaciones del productor.
La secuenciación nucleotídica de los productos purificados se realizó en el secuenciador automático CEQ 8800 Genetic Analyzer System (Beckman Coulter, USA) mediante el método de terminación de cadena por dideoxinucleótidos.
Análisis de las secuencias nucleotídicas
Las secuencias de los virus pandémicos y estacionales obtenidas directamente de las muestras clínicas de los pacientes cubanos infectados por estos virus, se publicaron en la base de datos del banco de genes (GENBANK) de libre acceso en internet, bajo los números de acceso: HQ159391 al HQ159418; HM176606 al HM176614 y HQ190930 al HQ190946.
Las secuencias se editaron utilizando los programas EditSeq y SeqMan del paquete DNASTAR (Lasergene, Inc., Madison, WI) y los alineamientos múltiples y las secuencias consenso se obtuvieron de los análisis con el editor de alineamientos de secuencias BioEdit v.7.0.1.13 La traducción a aminoácidos para la búsqueda de mutaciones se realizó en el programa MEGA v.4.0.14 Los árboles filogenéticos se construyeron por dos programas diferentes, el MEGA v.4.0 y BEAST.15 Los árboles filogenéticos construidos por el programa BEAST se visualizaron y editaron en el programa FigTree v.1.3.1.
RESULTADOS
La OMS ha estimado cada año que la influenza contabiliza entre un cuarto y medio millón de muertes en todo el mundo. Actualmente, la vacunación es el único medio práctico de reducir o contrarrestar la carga de morbilidad y mortalidad en la comunidad.16 Sin embargo, la producción de una vacuna anti-influenza óptima requiere de un monitoreo global continuo de los virus influenza, para detectar la emergencia de virus que son diferentes de los que circularon antes. La habilidad de los virus influenza para evadir la inmunidad adquirida en respuesta a la infección o vacunación por las derivas antigénicas progresivas, exige una actualización regular de la composición de las vacunas anuales, que reflejen a los virus contemporáneos.17
La necesidad de la vigilancia global de los virus influenza fue reconocida en 1947 y propició la organización de la Red de Vigilancia Global de Influenza (RVGI) de la OMS, de la que son parte importante los Centros Nacionales de Influenza (CNI). En conjunto, los laboratorios miembros de la RVGI procesan alrededor de 500 000 muestras respiratorias por año, para monitorear la actividad del virus influenza en todo el mundo. Alrededor de 8 000 virus de los aislados por los CNI son enviados a los Centros Colaboradores de la OMS (CCOMS) para una caracterización antigénica y genética más extensiva.18 En Cuba, el CNI ubicado en el IPK, es el encargado de la caracterización antigénica y genética de los virus influenza.
Recientemente la humanidad se vio afectada por la primera pandemia del siglo XXI, detectada primero en México.19 Cuba no fue la excepción, razón por la cual el CNI de Cuba se dio a la tarea de trazar estrategias que permitieron la caracterización molecular de los virus influenza que circularon en este período.
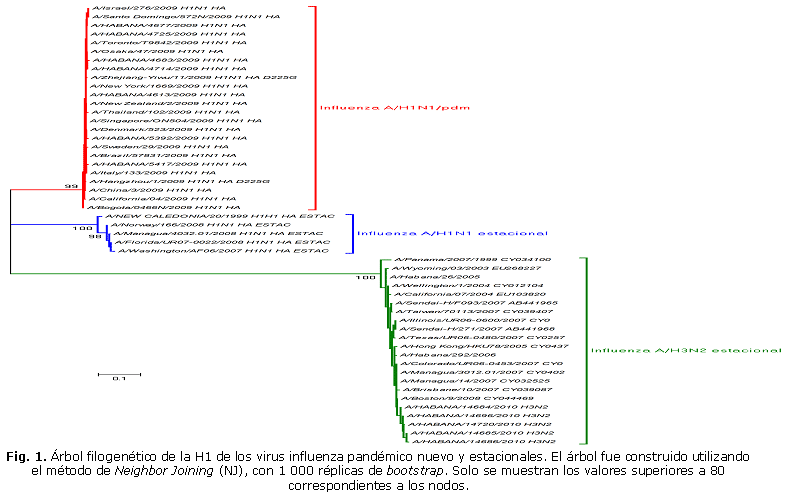
La estrategia 1 posibilitó obtener las primeras secuencias de un segmento del gen de la HA de los virus influenza A, incluido el virus pandémico. Aunque los segmentos obtenidos mediante esta estrategia eran de un tamaño alrededor de los 1 000 pb, después de la edición de las secuencias se obtenía un segmento útil de aproximadamente 600 pb. Estos resultados fueron valiosos en cuanto a la diferenciación del virus A/H1N1 estacional del pandémico, sobre todo en los primeros meses de la pandemia, cuando no contábamos con el diagnóstico por RCP en tiempo real, confirmando su valor como confirmatorio (Fig. 1).
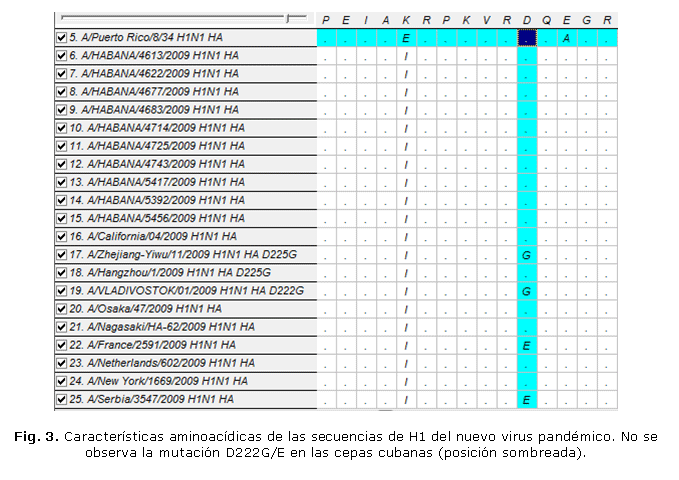
La estrategia 2 permitió obtener las secuencias de la HA y por primera vez de la NA de los virus influenza A en Cuba, incluido el virus pandémico (Fig. 2). En este caso se pudieron obtener los segmentos casi completos de los mencionados genes, aun después de la edición de las secuencias. Esta estrategia, al igual que la 1, permitió la vigilancia de la mutación D222G/E (Fig. 3). Sin embargo, para las muestras con valores de CT>30 no fue posible obtener un producto de RCP de calidad, por lo que no se pudo obtener sus secuencias por esta estrategia. Este problema fue visto sobre todo con las muestras de tejido de pulmón proveniente de pacientes fallecidos. Esta estrategia permitió además, iniciar la vigilancia de la posible emergencia de cepas mutantes H275Y resistentes a los INA, específicamente al tamiflu (Fig. 4).
Con las reacciones de la estrategia 3 se lograron obtener productos de gran calidad en cuanto a concentración y pureza para ambos genes analizados. Los productos obtenidos a partir de las muestras de tejido de pulmón resultaron óptimos y esto permitió lograr secuencias de alta calidad a partir de esas muestras tan valiosas.
DISCUSIÓN
Aunque las estrategias 1 y 2 no fueron las más adecuadas, se constató que cada una tuvo su importancia en un contexto determinado.
A inicios de la pandemia, la OMS publicó un listado de países con la capacidad diagnóstica para el nuevo virus.20 Esta lista se confeccionó teniendo en cuenta los resultados del Proyecto de Evaluación de la Calidad Externa de la OMS (PECE/OMS),21 para lo cual era necesario haber obtenido 100 % de resultados positivos en los dos últimos paneles. Si bien los resultados que permitieron a Cuba estar en esta lista fueron logrados con los sistemas de RCP múltiple con los que contaba el CNI, cuando fue incluido el nuevo virus en el panel, la estrategia 1 permitió confirmar su detección. Auún así, esta estrategia no se consideró como la óptima porque el segmento no era lo suficiente grande como para obtener toda la información posible sobre el nuevo virus y, además, no se contaba con las secuencias del gen de la NA para los análisis de resistencia.
Es en este punto donde cobra importancia la estrategia 2, que al ser introducida, permite obtener secuencias más largas de los genes HA y las primeras secuencias de la NA. Esto permitió introducir la vigilancia de mutantes H275Y entre los virus pandémicos.22
Por su parte, la estrategia 3 posibilitó obtener los mismos resultados que la 1 y la 2. Sin embargo, permitió también la obtención de secuencias a partir de las muestras de tejido de pulmón y con mejor calidad, objetivo que no se alcanzó con las estrategias 1 y 2.
Al realizar el análisis de las secuencias de un segmento del gen de la HA obtenidas en cualquiera de las tres estrategias, se pudo diferenciar entre los virus influenza A estacionales y el pandémico, que se demuestra su utilidad como método confirmatorio de los casos sospechosos (Fig. 1). Cuando se quiere realizar el diagnóstico diferencial entre los virus influenza, se debe tener en cuenta que la región amplificada por los cebadores pueda establecer la diferencia entre los subtipos.23
Los segmentos del gen de la HA obtenidos mediante la estrategia 2 fueron mayores que los de la 1, lo cual permite obtener mayor información sobre este gen en el nuevo virus pandémico. Por ejemplo, se pudo comprobar que al menos la mutación D222G/E,24 presente en algunos casos con diagnóstico clínico de IRAG y fallecidos, no fue encontrada en las muestras cubanas analizadas (Fig. 3). Las secuencias para el caso de las muestras de tejido de pulmón, se obtuvieron utilizando la estrategia 3, porque al aplicar una TR-RCP anidada como método, se ganó en sensibilidad y se obtuvieron segmentos con una mayor calidad y concentración de ácido desoxirribonucleico (ADN). Sin embargo, en las muestras cubanas se hallaron otros cambios importantes, los cuales merecen un análisis más profundo.
Cuando se analiza un segmento del gen de la NA, de igual forma se pudo realizar el diagnóstico diferencial de los virus estacionales con el pandémico (Fig. 2), apoyado también por la selección correcta de los cebadores.23
La obtención de estos segmentos resultó posible por las estrategias 2 y 3, sin embargo, solo la estrategia 3 permitió obtener los segmentos de calidad mayor en el caso de las muestras de tejido de pulmón. Al contar con los segmentos de la NA se inició en nuestro laboratorio la vigilancia de cepas emergentes resistentes al tamiflu (INA).22 Los análisis de las muestras cubanas no mostraron ningún caso de resistencia al tamiflu, incluidos los pacientes que padecieron ETI, IRAG y pacientes fallecidos (Fig. 4). Sin embargo, hay que ser cuidadosos a la hora de analizar este resultado porque se han encontrado variantes del virus A/H1N1 pdm mutantes H275Y que aparecen de forma natural y no se trasmiten, es decir, aparecen de forma puntual en determinados pacientes. Existen reportes en la literatura internacional que apoyan esta observación, donde expresan que los virus influenza resistentes a los INA pueden diferenciarse en cuanto a la fijación y transmisibilidad, en dependencia del nivel de pérdida de la funcionalidad de la NA.25 No obstante, se ha podido ver lo contrario en pacientes que padecen de influenza A/H1N1 estacional.26
Las tres estrategias utilizadas en el CNI de influenza en Cuba permitieron caracterizar molecularmente al nuevo virus influenza A/H1N1pdm. Cada una permitió realizar el diagnóstico diferencial entre los virus estacionales y pandémicos. La estrategia considerada como la más completa fue la tercera porque además de permitir el diagnóstico diferencial, posibilitó obtener secuencias de HA mayores que la estrategia 1 y de mayor calidad que la 1 y la 2 para los genes HA y NA en el caso de muestras de tejido de pulmón. Las secuencias de HA en las cepas cubanas analizadas no presentaron la mutación D222G/E, mientras que las secuencias de NA no mostraban la mutación H275Y que es un marcador molecular de resistencia al tamiflu. No obstante, se recomienda realizar estudios de un número mayor de muestras para poder llegar a una conclusión definitiva.
AGRADECIMIENTOS
A los colegas miembros del CNI del Instituto de Salud Carlos III y a los del Centro Colaborador de la Organización Mundial de la Salud de Londres por su cooperación con los protocolos para la obtención de los productos de secuencia. A la oficina de la Organización Panamericana de la Salud (OPS) en Cuba por su colaboración constante.
REFERENCIAS BIBLIOGRÁFICAS
1. Lackenby A, Thompson CI, Democratis J. The potential impact of neuraminidase inhibitor resistant influenza. Curr Opin Infect Dis. 2008;21(6):626-38.
2. 2009 pandemic influenza A(H1N1) virus mutations reported to be associated with severe disease [cited 2009]. Available in: http://www.ecdc.europa.eu/en/activities/sciadvice
3. Global influenza surveillance network: laboratory surveillance and response to pandemic H1N1 2009. Wkly Epidemiol Rec. 2009;84(36):361-5.
4. Fiore AE, Uyeki TM, Broder K, Finelli L, Euler GL, Singleton JA, et al. Prevention and control of seasonal influenza with vaccines: recommendations of the Advisory Committee on Immunization Practices (ACIP), 2010. MMWR Recomm Rep. 2010;59(RR-08):1-62.
5. Kumlin U, Olofsson S, Dimock K, Arnberg N. Sialic acid tissue distribution and influenza virus tropism. Influenza Other Respi Viruses. 2008;2(5):147-54.
6. Stevens J, Blixt O, Paulson JC, Wilson IA. Glycan microarray technologies: tools to survey host specificity of influenza viruses. Nat Rev Microbiol. 2006;4(11):857-64.
7. Shinya K, Ebina M, Yamada S, Ono M, Kasai N, Kawaoka Y. Avian flu: influenza virus receptors in the human airway. Nature. 2006;440(7083):435-6.
8. Qi L, Kash JC, Dugan VG, Wang R, Jin G, Cunningham RE, et al. Role of sialic acid binding specificity of the 1918 influenza virus hemagglutinin protein in virulence and pathogenesis for mice. J Virol. 2009;83(8):3754-61.
9. Coiras MT, Perez-Brena P, Garcia ML, Casas I. Simultaneous detection of influenza A, B, and C viruses, respiratory syncytial virus, and adenoviruses in clinical samples by multiplex reverse transcription nested-PCR assay. J Med Virol. 2003;69(1):132-44.
10. World Health Organization (WHO). CDC protocol of realtime RTPCR for influenza A (H1N1). [cited 2009]. Available in: http://www.who.int/csr/resources/publications/swineflu /realtimeptpcr/en/index.html
11. Tenorio-Abreu A, Eiros JM, Rodriguez E, Bermejo JF, Dominguez-Gil M, Vega T, et al. Influenza surveillance by molecular methods. Rev Esp Quimioter. 2009;22(4):214-20.
12. WHO information for laboratory diagnosis of pandemic (H1N1) 2009 virus in humans - revised. [cited 2009]. Available in: http://www.who.int/csr/resources/publications/swineflu/diagnosticrecommendations/en/index.html
13. Hall TA. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp Ser (Oxf). 1999;41:95-8.
14. Tamura K, Dudley JP, Nei M, Kumar S. MEGA4: Molecular Evolutionary Genetics Analysis (MEGA) Software Version 4.0. Mol Biol Evol. 2007;24(8):1596-9.
15. Drummond A, Rambaut A. BEAST: Bayesian evolutionary analysis by sampling trees. BMC Evol Biol. 2007;7:214.
16. Barr IG, McCauley J, Cox N, Daniels R, Engelhardt OG, Fukuda K, et al. Epidemiological, antigenic and genetic characteristics of seasonal influenza A(H1N1), A(H3N2) and B influenza viruses: basis for the WHO recommendation on the composition of influenza vaccines for use in the 2009-2010 Northern Hemisphere season. Vaccine. 2009;28(5):1156-67.
17. Salomon R, Webster RG. The influenza virus enigma. Cell. 2009;136(3):402-10.
18. WHO influenza surveillance. Wkly Epidemiol Rec. 1996;71(47):353-7.
19. Schnitzler SU, Schnitzler P. An update on swine-origin influenza virus A/H1N1: a review. Virus Genes. 2009;39(3):279-92.
20. Countries able to perform PCR to diagnose influenza A (H1N1) virus infection in humans. [cited 2009]. Available in: http://www.who.int/csr/resources/publications/swineflu /country_pcr_capacity/en/index.html
21. WHO external quality assessment project for detecting influenza virus subtype A by polymerase chain reaction - summary analysis, 2009. Wkly Epidemiol Rec. 2009;84(48):493-500.
22. van der Vries E, Jonges M, Herfst S, Maaskant J, Van der Linden A, Guldemeester J, et al. Evaluation of a rapid molecular algorithm for detection of pandemic influenza A (H1N1) 2009 virus and screening for a key oseltamivir resistance (H275Y) substitution in neuraminidase. J Clin Virol. 2009;47(1):34-7.
23. Chander Y, Jindal N, Stallknecht DE, Sreevatsan S, Goyal SM. Full length sequencing of all nine subtypes of the neuraminidase gene of influenza A viruses using subtype specific primer sets. J Virol Methods. 2010;165(1):116-20.
24. Kilander A, Rykkvin R, Dudman SG, Hungnes O. Observed association between the HA1 mutation D222G in the 2009 pandemic influenza A(H1N1) virus and severe clinical outcome, Norway 2009-2010. Euro Surveill. 2010;15(9).
25. Update on oseltamivir-resistant pandemic A (H1N1) 2009 influenza virus: January 2010. Wkly Epidemiol Rec. 2009;85(6):37-40.
26. Schilling M, Gravenstein S, Drinka P, Cox N, Krause P, Povinelli L, et al. Emergence and transmission of amantadine-resistant influenza A in a nursing home. J Am Geriatr Soc. 2004;52(12):2069-73.
Recibido: 19 de octubre de 2010.
Aprobado: 20 de noviembre de 2010.
Alexander Piñón Ramos. Instituto de Medicina Tropical "Pedro Kourí". Autopista Novia del Mediodía Km 6 1/2, Lisa. La Habana, Cuba. Correo electrónico: alexpr@ipk.sld.cu


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}