










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkVaccimonitor
versión On-line ISSN 1025-0298
Vaccimonitor v.18 n.1 Ciudad de la Habana ene.-abr. 2009
ARTICULOS ORIGINALES
Obtención y caracterización parcial de un extracto lipídico de la membrana externa de Mycobacterium smegmatis
Obtainment and partial characterization of a lipidic fragment of the outer membrane of Mycobacterium smegmatis
Nadine Alvarez1, Reinier Borrero1, María de los Ángeles García1, Ileana Martínez1, Michel Acosta1, María de los Ángeles Padrón1, Miguel Echemendía2, Ernesto Montoro2, Norazmi Mohd3, Maria Elena Sarmiento1 y Armando Acosta 1.
1. Instituto Finlay. Centro de Investigación–Producción de Vacunas. Ave. 27 No. 19805, La Lisa. Ciudad de la Habana, Cuba.
2. Instituto de Medicina Tropical «Pedro Kourí « (IPK). Apdo Postal 601, Marianao 13, La Habana, Cuba.
3. School of Health Sciences, University Sains Malaysia, 16150 Kubang Kerian, Malaysia.
email: nalvarez@finlay.edu.cu
RESUMEN
En la actualidad, los antígenos lipídicos de las micobacterias constituyen blancos atractivos para el desarrollo de nuevas formulaciones vacunales contra la tuberculosis. En nuestro trabajo se realizó la caracterización parcial de un extracto lipídico de pared celular de Mycobacterium smegmatis mediante cromatografía de capa delgada y Dot blot frente a gammaglobulina humana. Se identificó, fundamentalmente, la presencia de fosfolípidos y ácidos micólicos en el extracto lipídico y se observó un elevado reconocimiento de los mismos por la gammaglobulina humana, lo cual indica la importancia de continuar los estudios de inmunoprotección empleando antígenos lipídicos de micobacterias.
Palabras clave: Tuberculosis, Mycobacterium smegmatis, lípidos.
ABSTRACT
Currently, lipid antigens of mycobacteria are attractive targets for the development of new tuberculosis vaccinal formulations. A lipid extract of Mycobacterium smegmatis cell wall was characterized using a Thin Layer Chromatography and Dot blot with human gammaglobulin. Mainly we identified the presence of phospholipids and micolic acids in the lipid extract showing a high recognition by human gammaglobulin. These results indicate the relevance of continuing immunoprotection studies with mycobacterial lipid antigens.
Keywords: Tuberculosis, Mycobacterium smegmatis, lipids.
INTRODUCCION
Por décadas la tuberculosis (TB) ha constituido una de las enfermedades que mayor número de muertes ha causado al hombre. En la actualidad, a pesar de la vacunación con BCG y los tratamientos con antibióticos combinados, varios factores promueven el desarrollo de numerosas investigaciones con el único y común propósito de obtener una nueva vacuna contra la TB (1). Entre estos factores se encuentran: la coinfección de VIH con Mycobacterium tuberculosis, la aparición de cepas resistentes a múltiples fármacos antituberculosos y las condiciones socioeconómicas desfavorables. La TB continúa siendo un serio problema de salud pública en la actualidad, no sólo en los países en vías de desarrollo, sino también en los países industrializados.
BCG es la única vacuna disponible contra la TB, sin embargo la misma ha mostrado alta variabilidad en cuanto a eficacia se refiere, con rangos que oscilan entre un 0-80% (2), protege fundamentalmente contra las formas más severas de la enfermedad y no contra la forma pulmonar en la población adulta. Por ello, la obtención de una vacuna más eficaz contra la TB se ha convertido en un objetivo primordial para la comunidad científica mundial.
La gran mayoría de los antígenos empleados en los estudios encaminados a la obtención de vacunas contra la TB son de naturaleza proteica y muy pocas veces se ha explorado la incorporación de antígenos lipídicos. Sin embargo, una de las características que contribuye, en mayor medida, al elevado índice de patogenicidad de M. tuberculosis, es la pared celular impermeable y altamente hidrofóbica que posee (3). La pared celular de M. tuberculosis consiste en una capa de ácidos micólicos, arabinogalactano y peptidoglicano, provisto de una estructura que permite la inserción de lípidos que contienen tiocerol, micolatos de trealosa, fosfolípidos como los fosfatidilinositol manósidos y sus derivados glicosilados, tales como el lipomanano y lipoarabinomanano. Todos estos compuestos contribuyen de una forma u otra a la fisiología característica y a la capacidad que poseen las especies del género Mycobacterium, de producir enfermedad (4).
Los ácidos micólicos, en específico, poseen funciones biológicas importantes, entre las que se encuentra el papel que desempeñan en la persistencia de la micobacteria dentro del macrófago (5). Se conoce, además, que la presencia de niveles elevados de IgG circulantes contra varios antígenos micobacteriales, en especial el lipoarabinomanano, indican una menor incidencia de TB diseminada en niños (6), así como que la presencia de IgA secretora contra lipoarabinomanano en la saliva de contactos sanos de pacientes leprosos, se asocia con protección contra la TB (7).
Reportes recientes han demostrado que diferentes lípidos provenientes de la pared celular están implicados en algunos de los mecanismos de evasión e inhibición al sistema inmune del hospedero (8). En los últimos años, varios de estos lípidos se han convertido en moléculas diana para el desarrollo de nuevas vacunas debido a que han sido identificadas células Ta/b humanas que responden a antígenos lipídicos o glicolipídicos de micobacteria que son presentados por diferentes moléculas CD1, las cuales son no polimórficas (9). Por esta razón, los lípidos de micobacteria han sido empleados como constituyentes de adyuvantes y en inmunoterapias antitumorales (4), además de que se ha evidenciado en modelo experimental de curiel, que la inmunización con formulaciones que contienen lípidos de micobacteria disminuyen la aparición de lesiones a nivel pulmonar (10). Los lípidos de M. tuberculosis son importantes factores de virulencia (11), de manera que una buena estrategia consiste en desarrollar una respuesta inmune que los neutralice.
Mycobacterium smegmatis es una micobacteria no patógena de crecimiento rápido que ha sido empleada como modelo de estudio para la TB (12), ya que presenta muchas características en común con M. tuberculosis, a partir de comparaciones realizadas entre los genomas de ambas especies que han indicado la presencia de una elevada homología (13). Se especula que esto se debe, en mayor medida, a la presencia de rutas metabólicas muy conservadas, lo cual indica que comparten genes importantes en el metabolismo y exportación a la pared celular de componentes moleculares que pueden poseer un efecto inmunomodulador (14-16).
También se ha identificado la presencia de glicopeptidolípidos y fosfolípidos glicosilados en la pared celular de M. smegmatis que tienen la capacidad de inhibir la interacción de macrófagos humanos con la micobacteria, similares a los presentes en micobacterias patógenas como M. tuberculosis (17).
Teniendo en cuenta las características de M. smegmatis en cuanto a patogenicidad y crecimiento se refiere, resulta de interés utilizar esta especie como modelo de estudio para conocer la composición lipídica de las micobacterias, por lo que en el presente trabajo nos propusimos realizar la caracterización del perfil de un extracto de lípidos totales de M. smegmatis y su reactividad contra una formulación comercial de gammaglobulina humana.
Materiales y Métodos
Cepa bacteriana y condiciones de crecimiento
Para los experimentos se empleó la cepa de M. smegmatis mc2155 (perteneciente a la colección del Laboratorio Nacional de Referencia de Tuberculosis del Instituto “Pedro Kourí”), conservada a -70 °C, en glicerol al 15%. Para la obtención de la biomasa, se inocularon 5 µL de M. smegmatis en precultivos que contenían 5 mL de medio Luria Bertani (LB) y se pusieron a crecer durante 48 h en zaranda con agitación a 5,75 x g y 37 °C. Luego de transcurrido este tiempo, se realizó la técnica de Zielh-Neelsen (18) para comprobar la pureza y el crecimiento micobacteriano. Posteriormente, se inocularon, por duplicado, 100 µL de los cultivos crecidos en dos Erlenmeyer que contenían 250 mL de medio LB cada uno y se pusieron a crecer bajo las mismas condiciones descritas anteriormente.
Intacglobín
Formulación comercial de gammaglobulinas humanas producida en la Empresa de Sueros y Hemoderivados, Ciudad de La Habana, para uso endovenoso con una concentración de 50 mg/mL.
Extracción de fosfolípidos
Una vez crecidos los cultivos, se recolectó la biomasa mediante centrifugación a 135 x g durante 30 min. El sobrenadante fue desechado y el botón celular fue almacenado a – 20 °C hasta el día siguiente. Ambos botones celulares fueron tratados con lisozima (7,3 mg de lisozima en 1 mL de solución reguladora Tris-HCl 0.02 mol/L, EDTA 1 mmol/L, pH 8,0). Luego se dejó reposar durante 1 h a 37 °C y posteriormente se sometió a sonicación durante 6 tiempos, alternando sonicación e incubación en hielo (10 segundos de sonicación y 10 segundos de incubación en hielo). Se adicionaron dos volúmenes de la mezcla de cloroformo-metanol (2:1) y mezclamos en agitador de tubos durante 20 min, seguido de un paso de centrifugación durante 30 min a 750 x g. El sobrenadante se colectó y el precipitado se volvió a tratar con cloroformo-metanol en igual proporción, se mezcló en agitador de tubos y se centrifugó bajo las mismas condiciones del paso anterior. Ambos sobrenadantes se mezclaron y se adicionaron dos volúmenes del reactivo de Folch (cloroformo 0,3 mL, metanol 4,8 mL, agua 4,7 mL y cloruro de potasio 72,5 mg). La muestra se sometió a secado a una temperatura de 50 °C hasta lograr la máxima evaporación.
Caracterización de los fosfolípidos extraídos
Con el objetivo de visualizar y caracterizar los lípidos presentes en la muestra de M. smegmatis, se desarrollo la técnica de Cromatografía de Capa Delgada unidimensional (del inglés Thin Layer Chromatographic, TLC) para fosfolípidos. La muestra se disolvió en 5 µL de cloroformo y se aplicó en una placa de sílica gel 60 en soporte de vidrio, en forma de punto, en el extremo inferior izquierdo de la placa. La placa se introdujo en una cámara con una atmósfera que se dejó saturar con una mezcla de cloroformo-metanol-agua (65:25:4) durante 1 h. Luego de la corrida se secó la placa y se reveló utilizando reactivo Ditmer (óxido de molibdeno 4 g y molibdeno 0,18 g en 100 mL de H2SO4) y sulfato de cobre al 10% en ácido fosfórico al 8%, con un paso de secado al aire después de rociar con Ditmer. La placa se calentó a 110 °C durante 10 min después de rociar con ácido fosfórico.
Dot blot
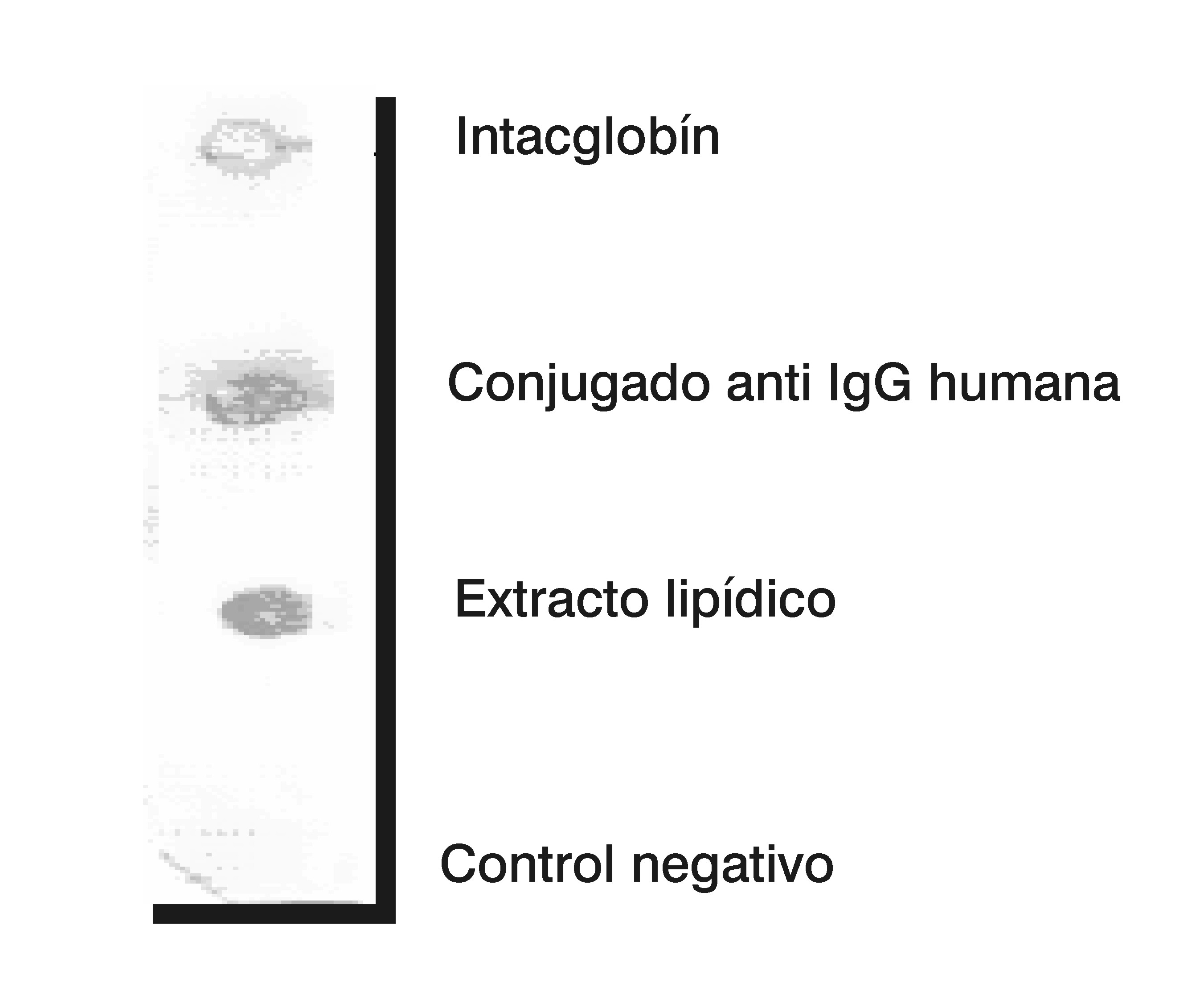
Se aplicaron 10 µL del extracto lipídico, 10 µL de solución salina tamponada con fosfato 0,15 M pH 7,2 (SSTF) como control negativo, 10 µL de conjugado anti IgG humana a la dilución de trabajo, como control positivo, y 10 µL de Intaglobín, colocados en diferentes puntos del papel de nitrocelulosa. Se bloqueó con 10 mL de leche descremada al 3% en SSTF durante 1 h a 37 °C sin agitación. Se realizaron tres lavados con SSTF y posteriormente se incubó durante 2 h con 10 mL de leche descremada e Intacglobín (1/50), previamente disuelto en leche descremada con SSTF-Tween 20 al 0,1% y se incubó durante 4 h en agitación y temperatura ambiente.
Posteriormente, se realizaron lavados de 3 min con SSTF-Tween 20 al 0,1%. Se adicionaron 10 mL de leche descremada 1,5%, 0,1 mL del conjugado 1/100 (anti IgG humana, Fc específica, desarrollada en carnero y conjugado a peroxidasa, SIGMA), y se incubó durante 1 h en agitación, a temperatura ambiente. Nuevamente, se efectuaron lavados con SSTF-Tween 20 al 0,1% de 3 a 4 veces durante 5 min y posteriormente, tres lavados con SSTF. Finalmente, la muestra fue revelada en la oscuridad con diaminobenzidina en SSTF y se detuvo la reacción con agua destilada.
Resultados y Discusión
Entre los principales lípidos que se localizan en la superficie de la pared celular de las micobacterias se encuentran los fosfolípidos (19), los cuales se ha observado que contribuyen, en cierta medida, a la patogenicidad de diversas micobacterias patógenas, ya que se ha demostrado que los mismos participan en diversos procesos que posibilitan la evasión de la micobacteria a los elementos del sistema inmune a través de la inhibición de la fagocitosis por los macrófagos (20, 21).
En el presente trabajo se procedió a caracterizar el perfil de fosfolípidos presentes en un extracto total de lípidos de pared celular de M. smegmatis, obtenido mediante un método estándar de extracción con cloroformo-metanol. La composición lipídica del extracto fue analizada mediante TLC, empleando fosfolípidos conocidos como estándares para la comparación. Los lípidos polares identificados fueron correspondientes a fosfatidil colina, fosfatidil serina y fosfatidil etanolamina (Figura 1).
Además, se observaron manchas correspondientes a moléculas apolares, las cuales estimamos que se correspondan con los ácidos micólicos presentes en el extracto de pared celular, cuya hidrofobicidad es muy elevada y de acuerdo con estudios realizados previamente en Mycobacterium habana (20), la migración de estos ácidos grasos es mayor que el resto de los lípidos presentes en la muestra (22).
El análisis de ácidos micólicos mediante TLC ha sido empleado por diversos laboratorios como un método rápido para la identificación de aislamientos clínicos y ambientales de micobacterias (23).
Adicionalmente, nos propusimos evaluar la reactividad de la población cubana frente a lípidos de micobacteria, para lo cual se empleó el Intacglobín por el hecho de ser una formulación que representa la reactividad de la población cubana en general.
Como resultado, se observó reactividad frente al extracto lipídico obtenido de M. smegmatis (Figura 2), lo que nos sugiere que una población de individuos sanos presenta anticuerpos capaces de reconocer antígenos de naturaleza lipídica de micobacterias pertenecientes al género Mycobacterium, a partir del hecho de que el Intacglobín es un compuesto hemoderivado obtenido del suero de individuos sanos de la población cubana.
Dicha formulación ha mostrado protección pasiva en modelos experimentales de ratón frente a la infección por M. bovis (24).
Figura 1. Crometografia en capa delgada de fosfolipidos extraidos de Mycobacterium smegmatis. Revelado con Ditmer.FC:estandar de fosfatidil colina. FE:estandar de fosfatodil etanolamina.FS:estandar de fosfatidil serina. Msm:extracto de M. smegmatis(2,5 uL).
Figura 2. Dot blot de fosfolipidos extraidos de Mycobacterium smegmatis.
Este reconocimiento, pudiera deberse en cierta medida a que la mayor parte de la población cubana es vacunada con M. bovis (BCG) al nacer. Colateralmente, consideramos que la exposición continua a micobacterias ambientales no patógenas puede influir en este tipo de reconocimiento. Las micobacterias ambientales están ampliamente diseminadas y raramente infectan a humanos inmunocompetentes. Estas pueden producir enfermedad pulmonar indistinguible, clínicamente, de las producidas por el bacilo tuberculoso. Recientemente, el aislamiento de micobacterias ambientales se ha incrementado, e incluso en algunos países ha superado el diagnóstico de M. tuberculosis y su identificación es frecuente en individuos inmunosuprimidos como es el caso de los pacientes con SIDA (25).
En nuestro país, se han realizado diversos estudios de identificación de especies micobacterianas a las cuales se encuentran expuesta la población cubana. Tal es el caso del estudio desarrollado por Mederos y cols. (26), en el cual se aislaron diferentes especies de micobacterias (Mycobacterium avium intracellulare, Mycobacterium fortuitum, Mycobacterium chelonae y Mycobacterium malmoense), a partir de muestras clínicas pulmonares y extrapulmonares procedentes de pacientes sintomáticos de diferentes provincias de nuestro país, con vistas a analizar su frecuencia de aislamiento según los diferentes grupos establecidos para la clasificación micobacteriana.
De acuerdo con los resultados obtenidos mediante el empleo de la cromatografia en capa delgada y el Dot blot, se puede afirmar que se obtuvo un extracto de pared de M. smegmatis que contiene fosfolípidos y posiblemente ácidos micólicos. Además, dicho extracto es reconocido por los anticuerpos presentes en la formulación comercial de gammaglobulina humana, Intacglobín.
Consideramos necesario profundizar en la caracterización de lípidos de este extracto empleando otras técnicas como espectrometría de masa, análisis espectroscópico mediante resonancia magnética nuclear, combinado con diferentes métodos de degradación química, así como profundizar en los estudios sobre el papel inmunoprotector de los lípidos de pared celular de M. smegmatis, en aras de comprobar la factibilidad de su uso como candidato en la investigación de nuevas vacunas de uso humano contra la tuberculosis.
Referencias
1. Skeidy YA, Sadoff JC. Advances in tuberculosis vacine strategies. Nat Rev Microbiol 2006; 4:469-76.
2. Cegielski JP, Chin DP, Espinal MA, Frieden TR, Cruz RR, Talbot EA, et al. The global tuberculosis situation: progress and problems in the 20th century, prospects for the 21st century. Infect Dis Clin N Am 2002; 16:1-7.
3. Brennan PJ, Nikaido H. The envelope of mycobateria. Annu Rev Biochem 1995;64:29-63.
4. Gorocica P, Jiménez-Martínez M del C, Garfias Y, Sada I, Lascurain R. Componentes glicosilados de la envoltura de Mycobacterium tuberculosis que intervienen en la patogénesis de la tuberculosis. Rev Inst Nal Enf Resp Mex 2005; 18(2):142-53.
5. Glickman GS, Jacobs Jr WR. Microbial pathogenesis of Mycobacterium tuberculosis: dawn of a discipline. Cell 2001;104:477-85.
6. Costello AM, Kumar A, Narayan V, Akba MS, Ahmed S, Abou-Zeid C, et al. Does antibody to mycobacterial antigens, including lipoarabinomannan, limit dissemination in childhood tuberculosis? Trans R Soc Trop Med Hyg 1992; 86:686–92.
7. Ramaprasad P, Fernando A, Madhale S, Rao JR, Edward VK, Samson PD, et al. Transmission and protection in leprosy: indications of the role of mucosal immunity. Lepr Rev 1997; 68(4):301-15.
8. Kaur D, Berg S, Dinadayala P, Gicquel B, Chatterjee D, McNeil MR, et al. Biosynthesis of mycobacterial lipoarabinomannan: Role of a branching mannosyltransferase. Proc Natl Acad Sci USA 2006;103:13664-9.
9. Grant EP, Degano M, Rosat JP, Steffen S, Modlin RL, Wilson I A, et al. Molecular Recognition of Lipid Antigens by T Cell Receptors. J Exp Med 1999;189:195-205.
10. Dascher CC, Hiromatsu K, Xiong X, Morehouse C, Watts G, Liu G, et al. Immunization with a mycobacterial lipid vaccine improves pulmonary pathology in the guinea model of tuberculosis. International Immunology 2003;15: 915-25.
11. López LM, Díaz F, Vallecillo AJ, Esquivel H, Gutiérrez JA. Tuberculosis humana y bovina en Latinoamérica: De estudios sobre virulencia hacia herramientas para su control. Rev Latinoam Microbiol 2006; 48(2):173-78.
12. Chaturvedi V, Dwivedi N, Tripathi RP, Sinha S. Evaluation of Mycobacterium smegmatis as a possible surrogate screen for selecting molecules active against multi-drug resistant Mycobacterium tuberculosis. J Gen Appl Microbiol 2007;53:333–7.
13. Barry C. Mycobacterium smegmatis: an absurd model for tuberculosis? Trends in Microbiology 2001;9(10):473–474.
14. Converse SE, Cox JS. A Protein Secretion Pathway Critical for Mycobacterium tuberculosis Virulence Is Conserved and Functional in Mycobacterium smegmatis. J Bacteriol 2005;187:1238–45.
15. Henry SG, WolschendorfF, Abshire M, Niederweis M, Braunstein M. Identification of Two Mycobacterium smegmatis Lipoproteins Exported by a SecA2-Dependent Pathway. J Bacteriol 2007; 189:5090–100.
16. McCarthy TR, Torrelles JB, MacFarlane AS, Katawczik M, Kutzbach B, DesJardin LE. Overexpression of Mycobacterium tuberculosis manB, a phosphomannomutase that increases phosphatidylinositol mannoside biosynthesis in Mycobacterium smegmatis and mycobacterial association with human macrophages. Molecular Microbiology 2005; 58:774–90.
17. Villeneuve C, Etienne G, Abadie V, Montrozier H, Bordier C, Laval F, et al. Surface glycopeptidolipids of M. smegmatis specifically inhibit the phagocytosis of mycobacteria by human macrophages. J Biol Chem 2003; 278:51291-300.
18. Bishop PJ, Neumann G. The history of the Ziehl-Neelsen stain. Tubercle 1970; 51:196-206.
19. Ortalo-Magné A, Lemassu A, Lanéelle MA, Bardou F, Silve G, Gounon P, et al. Identification of the surface-exposed lipids on the cell envelopes of Mycobacterium tuberculosis and other mycobacterial species. J Bacteriol 1996;178:456–61.
20. Mederos LM, Valdivia JA, Valero-Guillen PL. Lipids of Mycobacterium habana, a synonym of Mycobacterium simiae with vaccine potential. Tuberculosis 2006; 86:324-9.
21. Thorson LM, Doxee D, Scott MG, Wheeler P, Stokes RW. Effect of Mycobacterial phospholipids on interaction of Mycobacterium tuberculosis with macrophages. Infection and Immunity 2001; 69:2172-9.
22. Fujimura Leite CQ, Oliveira de Souza CW, de Andrade Leite SR. Identification of Mycobacteria by Thin Layer Chromatographic Analysis of Mycolic Acids and Conventional Biochemical Method: Four Years of Experience Mem Inst Oswaldo Cruz 1998; 93(6): 801-5.
23. Falcão DP, Valentini SR, Leite CQF. Pathogenic and potentially pathogenic microorganisms as contaminants of fresh water from different sources. Water Res 1993; 27:1737-41.
24. Olivares N, León A, López Y, Puig A, Cádiz A, Falero G, et al. The effect of the administration of human gamma globulins in a model of BCG infection in mice. Tuberculosis 2006;86:268–72[ STANDARDIZEDENDPARAG]
25. Rodríguez C, Martínez JL. Micobacterias ambientales estudio de 10 años. Revista Ciencias.com. 2006. [Disponible en: http://www.revistaciencias.com/publicaciones/EEVpEFupAkFbrgJiFs.php].
26. Mederos LM, Fofana AS, Perovani MA, García G, Montoro EH. Identificación de especies micobacterianas en Cuba. Archivos venezolanos de farmacología y terapéutica 2007;26(2): 142-5.
Recibido: Noviembre de 2008
Aceptado: Enero de 2009


{kind=link}
{kind=link}