










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl vol.32 no.4 La Habana oct.-dic. 2015
TECHNIQUE
Development of a method to detect three frequent mutations in the CFTR gene using allele-specific real time PCR
Desarrollo de un método para la detección de tres mutaciones frecuentes en el gen CFTR mediante PCR en tiempo real alelo-específica
Juan E Figueredo-Lago1, Anny Armas-Cayarga1, Yaimé J González-González1, Teresa Collazo-Mesa2, Iria García de la Rosa1, Yenitse Perea-Hernández1, Elvia N Santos-González2
1 Departamento de Diagnóstico Molecular, Centro de Inmunoensayo, CIE. Calle 134 y Ave 25, Cubanacan, Playa, PO Box 6653, CP 11600, Cuba.
2 Centro Nacional de Genética Médica, CNGM, La Habana, Cuba.
ABSTRACT
Cystic fibrosis (CF) is an autosomal recessive disease caused by mutations in the cystic fibrosis transmembrane conductance regulator (CFTR) gene. A method was developed to detect three frequent CFTR mutations (F508del, 3120+1G>A and N1303K), using allele-specific real time PCR (ASPCR). Genomic DNA samples from nine patients carrying different CFTR mutations were evaluated in triplicate. The Ct variation (ΔCt) between wild type and mutant ASPCRs, and the mean value ± standard deviation (SD) were determined. The specificity of allele-specific primers was confirmed by High Resolution Melting (HRM) analysis. The results validated this test for genotype determination; the tested mutations were clearly identified in all the samples and in excellent agreement with the commercial panel StripAssay (Viennalab, Austria). The described method provides the necessary selectivity and specificity as required for clinical screening of CF population, mainly for F508del mutation due to its high prevalence among the Cuban population.
Keywords: CFTR gene mutations, allele-specific real time PCR, Ct variation, cystic fibrosis.
RESUMEN
La fibrosis quística (FQ) es una enfermedad autosómica recesiva provocada por mutaciones en el gen regulador de la conductancia transmembranal de la fibrosis quística (CFTR). Se desarrolló un método para detectar tres mutaciones frecuentes en el gen CFTR (F508del, 3120+1G>A and N1303K), mediante PCR en tiempo real alelo-específica (ASPCR). Se evaluaron por triplicado las muestras de ADN genómico de nueve pacientes que portaban las mutaciones CFTR mencionadas, y se determinó la variación de Ct (ΔCt) entre las reacciones de ASPCR para los genotipos mutado y salvaje, expresada como media ± la desviación estándar. La especificidad de los cebadores por los alelos se confirmó mediante análisis de Fusión de Alta Resolución (HRM). Los resultados validaron la utilidad del método implementado para el genotipado, y se identificaron las tres mutaciones, según se pudo corroborar al comparar los resultados obtenidos con las determinaciones hechas mediante el panel comercial StripAssay (Viennalab, Austria). El método descrito mostró la selectividad y la especificidad necesarias para la pesquisa clínica poblacional de la FQ, principalmente de la mutación F508del, dada su alta prevalencia entre la población cubana.
Palabras clave: mutaciones en el gen CFTR, PCR en tiempo real alelo-específica, variación de Ct, fibrosis quística.
INTRODUCTION
Mutations in the Cystic Fibrosis Transmembrane Regulator (CFTR) gene are associated with cystic fibrosis (CF), as the most common life-threatening autosomal recessive genetic disorder [1]. But also male infertility due to congenital bilateral absence of the vas deferens (CBAVD), idiopathic chronic pancreatitis and bronchiectasis are CFTR-related disorders. One mutation (F508del) accounts for about 70 % of CF alleles worldwide [2], and diverse heritages are reflected for the CFTR gene and distributed with varying frequencies among populations often complicating genetic analysis. Worldwide, most mutations are rare, with frequencies under 0.1 %, and just four mutations have relative frequencies of 1.2-2.4 % [3]. Regardless the frequency, F508del is always screened when testing for CFTR mutations [4].
Different detection methods have been implemented to detect the CFTR gene, among them: single-strand conformation polymorphism analysis (SSCP) [5], restriction fragment length polymorphism analysis, amplification refractory mutation system (ARMS) and real-time PCR systems using SYBR Green [6]. Commonly, exon analysis using SSCP or denaturing gradient gel electrophoresis (DGGE) are performed as indirect detection methods, but they are very laborious and time-consuming techniques, making them incompatible for the screening of a large number of samples.
Among the systems developed, allele-specific PCR (ASPCR) is an advantageous strategy which combines amplification and detection, and the successfully amplified product directly indicates the occurrence of a particular allele when present. The use of ASPCR together with real-time PCR techniques allows monitoring template amplification, consequently improving the interpretation of PCR results [7]. A typical ASPCR assay uses a specific primer to detect both the wild type (wt) and the mutant (mut) DNA sequences, and a common reverse primer. By these means, individuals can be genotyped by analyzing the resulting amplification products [8].
Theoretically, individuals without mutation generate a PCR product only in the wild type reaction, heterozygotes yields both the wild type and mutant reaction products, and homozygotes only in the mutant reaction. However, ASPCR involves a relative rather than an absolute phenomenon, hence, the mismatched template could also be amplified, although at much lower efficiency as compared with the matched one [9].
Since amplification of mismatched template occurs, the genotype was determined by comparing the Ct values (the number of PCR cycles at which reporter fluorescence becomes significant or is distinguishable from the background noise at a given threshold) of the sample in two parallel ASPCRs (wt and mut). The difference in Ct (ΔCt) could be used to predict the assay selectivity [10]. Similar Ct values could imply that both alleles, wt and mut, were present in the sample. There could be predicted the only allele (wt or mut) present in a sample just by comparing their Ct values, indicating a non-carrier or homozygous individual, depending on the earliest curve corresponding to the lower Ct. To attain such results, genomic DNA (gDNA) templates should be kept as homogeneous as possible to avoid variations between tubes corresponding to the same sample, which lead to false-positive results produced by the uneven distribution of different alleles of the same gene in the sample [11].
Based on these assumptions, here we describe the development of a fast and cost-effective method to detect three frequent CFTR mutations in Cuban population using ASPCR: F508del (c.1521-1523delCTT), 3120+1G>A (c.2988+1G>A) and N1303K (c.3909C>G). These three mutations change the polypeptide sequence of CFTR: phenylalanine is deleted at position 508, glycine is replaced by alanine at the first nucleotide in the intron following nucleotide 3120 and asparagine at position 1303 is replaced by lysine respectively [12]. The selectivity and specificity of the allele-specific real time PCR was confirmed through the cycle threshold variation (ΔCt) between wild type and mutant PCR products. The described method also provides the necessary selectivity and specificity re-quired for the analysis of clinical samples. Furthermore, it reduces the possibilities of sample cross-contamination by avoiding post-hybridization reactions and it is easily reproducible by real-time PCR technology, with potential for the detection of other CFTR mutations.
MATERIALS AND METHODS
Primers design
Primers were designed with their 3´ nucleotide overlapping the polymorphic residue. Three primers per target were designed, one pair of primers targeting the mutant or a wild type allele, respectively, using the default parameters as recommended for allele discrimination, and a common primer by following the same recommendations as those for allele-specific primers. The 3´ terminal base of each allele-specific primer was adapted according to its corresponding mutation. Primers sequences for the detection of 3120+1G>A and N1303K polymorphisms were designed at the Center of Immunoassay using the CFTR sequence data available on the Cystic Fibrosis Mutation Database [13]. Primer sequences for detection of F508del were taken from Ferrie et al. [14]. Primer sequences are described in Table 1.
DNA extraction
Human gDNA samples from nine CF patients carrying the most frequent mutations reported in Cuba were kindly provided by the National Center of Medical Genetics (CNGM, Cuba) [15]. The gDNA was isolated from 10 mL of peripheral blood treated with ethylenediaminetetraacetic acid (EDTA) by salting out and samples were subsequently stored at –20 °C until use. Prior to the analysis, the gDNA quality and concentration was spectrophotometrically determined (optical density at 260 nm; OD260nm), its final concentration in the range of 5-40 μg/mL.
Genotyping
Genotype was previously characterized in all the samples by using the commercial panel StripAssay 4-410 (Viennalab, Austria). Five out of nine samples tested were single heterozygous (only one mutation detected): G85E, I507del, R553X, 3120+1G>A and R1162X, three as compound heterozygous (two mutations detected): R334W/F508del, G542X/ F508del and N1303K/F508del; and one homozygous, F508del/F508del.
Real-time PCR assays
Three CFTR mutations were evaluated (F508del, 3120+1G>A and N1303K). Two PCR reactions per sample were run for every target (wt and mut). The wt allele-specific primer (ASP) of the corresponding target was added to one reaction and the mut ASP to the other. The common reverse primer was added to both reactions (wt and mut). Reactions were run in triplicate. The wt amplification curve was used as reference for the mut one and viceversa. All reactions were run in a reaction volume of 25 μL in 96-well plates with adhesive films and by using the SLAN 96-P real-time PCR system (Shanghai Honshi Medical Technology Co., Ltd; China). Each reaction mixture contained 1× Absolute qPCR SYBR Green Mix (Thermo Scientific, Liege, Belgium). During the optimization steps, primer concentrations were set to 0.3 μM; the volume was completed to 20 μL with DNase-free water and 5 μL of template gDNA (25-200 ng) was added for each sample. The thermal PCR profile was: 15 min at 95 °C for Thermo Start DNA Polymerase activation, followed by 38 amplification cycles (95 °C for 30 s, 59 °C for 30 s and 72 °C for 30 s), and a final extension step at 72 °C for 3 min.
The test threshold was set at 0.12. The average Ct values for the three replicas were determined and ΔCt calculated for each sample: ΔCt = Average Ct (mutant ASO) – Average Ct (wild type ASP). High Resolution Melting (HRM) analysis was performed to evaluate the specificity of the mutant allele-specific primers. PCR products were melted by increasing the temperature from 60 to 90 °C at a programmed rate of 0.1 °C/s, and melting curves analyzed with the aid of the commercial SLAN 96-P software, version 8.2 (Shanghai Honshi Medical Technology Co., Ltd; China).
RESULTS
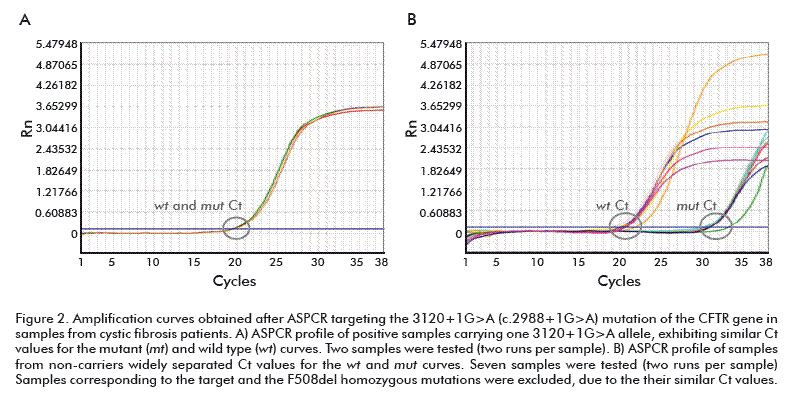
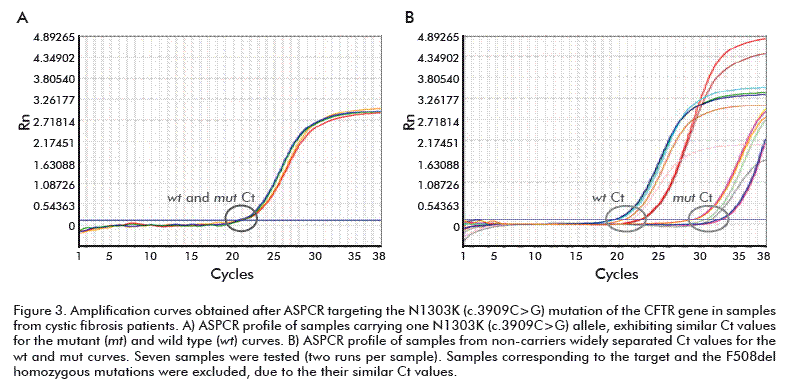
ASPCR reactions were performed to evaluate if the experimental conditions were capable to discriminate samples containing the mutations variants F508del, 3120+1G> or N1303K of CFTR, the typical amplification curves of ASPCRs containing wt or mut ASP targeting mutations are shown in figure 1, figure 2 and figure 3, respectively.
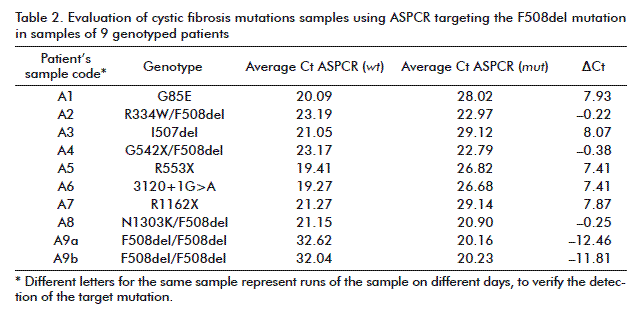
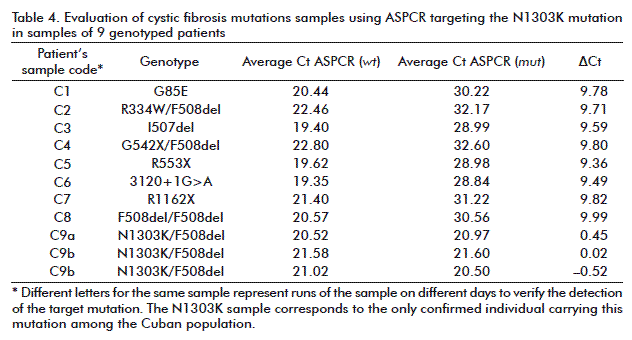
In the case of target F508del (Figure 1A), the amplification product in the reaction containing the mut ASP (red) was detected significantly earlier than that for the wt reaction, in the homozygous F508del sample. The similar accumulation of both ASPCR products allows identifying the heterozygote F508del sample (Figure 1B). Moreover, non-carrier F508del genotypes were identified (Figure 1C), inversely as compared with that previously seen for the homozygous sample, the amplification product in the wt ASP reaction been detected significantly earlier than the mut product. The same interpretation was given to 3120+1G>A and N1303K targets profiles shown in figure 2 and figure 3, respectively. Data resulting from PCR analysis of the three mutations analyzed are shown in table 2, table 3 and table 4.
The ΔCt values were widely separated to accurately determine the non-carrier and homozygoous genotypes in every sample. Similar ΔCt values were obtained for all heterozygous samples (–1 ≤ ΔCt ≤ 1). The ΔCt ranges for non-carriers of each mutation (mean value ± SD) were: 7.4 to 8.1 for F508del; 10.26 to 11.16 for 3120+1G>A; and 9.58 to 9.98 for N1303K. On the contrary, heterozygous carriers developed ΔCt ranges as follows: F508del, –0.20 to –0.36; 3120+1G>A, –0.14 to –0.18; and N1303K, –0.14 to –0.18. Homozygous samples (F508del) showed a mean ΔCt value of –12.13 ± 0.45, confirming the reliable detection reported for this frequent genotype (see table 2, table 3 and table 4).
All heterozygous samples demonstrated ΔCt values lower than 1 cycle, whereas non-carriers samples showed ΔCt > 7 values, the homozygous showing ΔCt < –11 (negative values). The specificity of ASPCR was verified using the derivative melting curve. As shown in figure 4, the amplification reactions were specific since only one peek per target was obtained. The melting temperatures of the generated amplicons were 80.0 °C (F508del), 77.55 °C (3120+1G>A) and 76.44 °C (N1303K).
DISCUSSION
Studies based on ASPCR have been developed for clinical research or testing of the IVS8 (TG) m(T)n locus in the CFTR gene [16], common mutations in KRAS and BRAF [7] and hepatitis B virus-resistant strains [17]. This study shows the development of a highly specific ASPCR method targeting three of the most common CFTR mutations (F508del, 3120+1G>A and N1303K) in the same IVS8 (TG)m(T)n locus among the Cuban population.
ASPCR is based on Taq polymerase lacking 3´ to 5´ exonuclease activity, the resulting mismatch between the 3´ end of the PCR primer and the template leading to highly reduced amplification efficiency [9]. As expected, a difference in the ΔCt values between the wt and mut ASPCR was high enough to correctly discriminate the presence of wt from the mut allele for the three mutations tested. This is in agreement with control determinations by using commercial panel StripAssay (Viennalab, Austria), with neither false positive nor doubtful result obtained.
Since CFTR mutations are variations at the germline level, individuals can be non-carriers (mutation absent), heterozygous (50 % frequency, in a single allele) or homozygous (100 %). For these reasons, genotyping assays just require 50 % selectivity; that is, assays must be able to detect one mutant template in the presence of one wild type template [9]. In this sense, 50 % selectivity was achieved for two targets (3120+1G>A and N1303K) and 100 % for F508del due to its homozygocity, in fact, the only one homozygous mutation. Homozygous combinations are rare [18], and neither 3120+1G>A nor N1303K homozygocity has not been reported in Cuba to date. But, significantly, the method developed is designed to be able to detect them in case that they would appear among the screened populations.
Regarding ΔCt values, Ct is sensitive to the initial amount of templates. If initial gDNA concentrations were too low, the first curve could emerge after 30 cycles and the second curve could not be visible, possibly making the analysis unreliable. So far, we established the reproducibility of the assay in the range of 25-200 ng of gDNA to avoid the displacement of the amplification curves to left or right, which could generate misleading data, in spite of not determining the sensitivity for these assay. Therefore, further studies will be conducted to establish the limits of sensitivity (i.e., lowest amount) of gDNA needed to attained reproducible results. This element is relevant for the implementation of high throughput screening and simultaneous determinations from single samples.
Finally, the specificity of these assays was demonstrated, with not any unspecific PCR products been amplified when samples carrying other related CFTR mutations were tested. The HRM analysis of mutant ASPCRs confirmed the presence of specific PCR products when at least one of the alleles in the sample was affected (Figure 4). The derivative melting curves showed only one peak, indicating a single amplified target on each reaction. Since HRM analysis was performed, therefore, a post-PCR melting analysis was not required. Nevertheless, Nagy et al. [19] suggested that a post-PCR melting analysis could complement data and lead to more accurate results, aiding to detect possible unspecific amplification signals. Using HRM data only, all heterozygotes and most homozygotes could be detected without mixing [20], although F508del is sometimes not distinguished from wild type [21]. The ΔCt values analysis used in this work solved this problem; therefore, avoiding the need for complementary analysis such as HRM to detect mutations.
Overall, the method developed provides some valuable advantages as compared with other methodologies. These assays do not require internal amplification controls. If no product is generated in the allele-specific reaction this can be due to the presence of inhibitors in the sample. Both amplification curves, wt and mut, should always be present. Our ASPCR-based method loses its reliability when both reactions from the same sample are not carried out at the same time. The assays should also be performed in a calibrated PCR system to avoid variations especially among tubes or wells.
Another relevant feature of this assay is that interpretation of the results is based on ΔCt differences between two reactions, easy to visualize and does not require additional software or the use of any complex algorithms for results discrimination. The method described herein allows fast confirmation (in less than 2 h) as compared with other more laborious standard procedures, of three CFTR mutations, and not requiring solid-phase hybridization or any other expensive sequencing process. Moreover, additional contamination risks are minimized since there is no need for additional processing after PCR, the samples never leaving the amplification plate.
Regarding CF, the identification of F508del and its association with the clinical severity of the disease supports a better understanding of its influence on clinical manifestations of the disease [22]. In fact, considering the high prevalence of the F508del mutation [12] and the advantages of the developed method, we could recommend to implement it as the primary massive screening tool, mainly in prenatal diagnosis and also for carrier screening. Due to the low prevalence of 3120+1G>A and N1303K, these methods should be used for confirmation after previous exon scanning techniques like DGGE or SSCP.
ACKNOWLEDGEMENTS
The authors thank to Dr. Fidel Rodríguez Cala for his support in the collection of the samples and Dr. Boris Acevedo Castro for his relevant comments and suggestions for this paper.
REFERENCES
1. Farrell PM. The prevalence of cystic fibrosis in the European Union. J Cyst Fibros. 2008;7(5):450-3.
2. Bobadilla JL, Macek M, Fine JP, Farrell PM. Cystic fibrosis: a worldwide analysis of CFTR mutations–correlation with incidence data and application to screening. Hum Mutat. 2002;19:575-606.
3. Kammesheidt A, Kharrazi M, Graham S, Young S, Pearl M, Dunlop C, et al. Comprehensive genetic analysis of the cystic fibrosis transmembrane conductance regulator from dried blood specimens – Implications for newborn screening. Genet Med. 2006;8:557-62.
4. Poulou M, Destouni A, Kakourou G, Kanavakis E, Tzetis M. Prenatal diagnosis for CF using High Resolution Melting Analysis and simultaneous haplotype analysis through QF-PCR. J Cyst Fibros. 2014;13(6):617-22.
5. Billard A, Laval V, Fillinger S, Leroux P, Lachaise H, Beffa R,et al. The allele-specific probe and primer amplification assay, a new real-time PCR method for fine quantification of single-nucleotide polymorphisms in pooled DNA. Appl Environ Microbiol. 2012;78(4):1063-8.
6. Dempsey E, Barton DE, Ryan F. Detection of five common CFTR mutations by Rapid-Cycle Real-Time Amplification refractory mutation system PCR. Clin Chem. 2004;50(4):773-5.
7. Lang A, Drexel H, Geller-Rhomberg S, Stark N, Winder T, Geiger K, et al. Optimized Allele-Specific Real-Time PCR Assays for the Detection of Common Mutations in KRAS and BRAF. J Mol Diagn. 2011;13(1):23-8.
8. Chen N, Prada AE. Development of allele-specific multiplex PCR to determine the length of poly-T in intron 8 of CFTR. PeerJ. 2014;2:e468.
9. Dennis YML. The amplification refractory mutation system. In: Dennis Lo YM, editor. Clinical applications of PCR. Totowa NJ: Humana Press Inc; 1998. p. 61-9.
10. Morlan J, Baker J, Sinicropi D. Mutation detection by real-time PCR: a simple, robust and highly selective method. PLoS ONE. 2009;4(2):e4584.
11. Nasis O, Thompson S, Hong T, Sherwood M, Radcliffe S, Jackson L, et al. Improvement in sensitivity of allele-specific PCR facilitates reliable noninvasive prenatal detection of Cystic fibrosis. Clin Chem. 2004.50(4):694-701.
12. Eshaque B, Dixon B. Technology platforms for molecular diagnosis of cystic fibrosis. Biotechnol Adv. 2006;24:86-93.
13. Cystic Fibrosis Mutation Database. 2015 [cited 2015 Sep 16]. Available from: http://www.genet.sickkids.on.ca/cftr
14. Ferrie RM, Schwarz MJ, Robertson NH, Vaudin S, Super M, Malone G, et al. Development, multiplexing and application of ARMS tests for common mutations in the CFTR gene. Am J Hum Genet. 1992;51:251-62.
15. Collazo T, Bofill AM, Clark Y, Hernández Y, Gómez M, Rodríguez F, et al. Common mutations in Cuban cystic fibrosis patients. J Cyst Fibros. 2009;8:47-9.
16. Costa C, Costa JM, Martin J, Boissier B, Goossens M, Girodon E. Multiplex allele-specific fluorescent PCR for haplotyping the IVS8 (TG)m(T)n locus in the CFTR gene. Clin Chem. 2008;54:1564-7.
17. Ntziora F, Paraskevis D, Haida C, Manesis E, Papatheodoridis G, Manolakopoulos S, et al. Ultrasensitive Amplification Refractory Mutation System Real-Time PCR (ARMS RT-PCR) assay for detection of minority hepatitis B virus-resistant strains in the era of personalized medicine. J Clin Microbiol. 2013;51(9):2893-900.
18. Hantash FM, Rebuyon A, Peng M, Redman JB, Sun W, Strom CM. Apparent homozygosity of a novel frame shift mutation in the CFTR gene because of a large deletion. J Mol Diagn. 2009;11(3):253-6.
19. Nagy B, Nagy RG, Lázár L, Bán Z, Papp Z. Detection of ΔF508del using quantitative real-time PCR, comparison of the results obtained by fluorescent PCR. Fetal Diagn Ther. 2007;22:63-7.
20. Liew M, Pryor R, Palais R, Meadows C, Erali M, Lyon E, et al. Genotyping of single-nucleotide polymorphisms by high-resolution melting of small amplicons. Clin Chem. 2004;50:1156-64.
21. Montgomery J, Wittwer CT, Kent JO, Zhou L. Scanning the cystic fibrosis transmembrane conductance regulator gene using high-resolution DNA melting analysis. Clin Chem. 2007;53(11):1891-8.
22. De Lima FA, Bertuzzo CS, Gonçalves de Oliveira MA, Ribeiro AF, Dirceu J. Screening for F508del as a first step in the molecular diagnosis of cystic fibrosis. J Bras Pneumol. 2013;39(3):306-16.
Received in September 2015.
Accepted in December 2015.
Juan E Figueredo-Lago. Departamento de Diagnóstico Molecular, Centro de Inmunoensayo, CIE. Calle 134 y Ave 25, Cubanacan, Playa, PO Box 6653, CP 11600, Cuba. E-mail: juan.figueredo@cie.cu.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}