










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl vol.35 no.2 La Habana abr.-jun. 2018
RESEARCH
Analysis of genetic polymorphism in wild Nicotiana species and Cuban cultivated tobacco (Solanaceae) through AFLP
Análisis de polimorfismo genético en especies salvajes de Nicotiana y tabaco cultivado en Cuba mediante AFLP
Yoannis Domínguez1, Sandra Peréz-Álvarez2, Marco A Magallanes-Tapia2, Jesús Alicia Chávez-Medina2, Eduardo F Héctor-Ardisana3
1 Universidad Agraria de La Habana Fructuoso Rodríguez Pérez. Km 23 1/2 Autopista Nacional, San José de las Lajas, CP 32700, Mayabeque, Cuba.
2 Departamento de Biotecnología Agrícola, Unidad Sinaloa, Instituto Politécnico Nacional, CIIDIR-IPN. Blvd. Juan de Dios Bátiz Paredes No 250. Guasave, Sinaloa, CP 81101, México.
3 Facultad de Ingeniería Agronómica, Universidad Técnica de Manabí, Portoviejo, CP EC130105, Ecuador.
ABSTRACT
Wild Nicotiana species are commonly used in tobacco’s breeding programs to obtain cultivars of enhanced productivity, and to improve its tolerance or resistance to diseases or different types of stress. In Cuba, tobacco production is one of the main sources of economic income and during the last decades several new tobacco varieties have been generated, being essential to study their genetic background for better crop management. In this work,the genetic polymorphism of four Cuban varieties of Nicotiana tabacum L. and six wild species used in Cubantobacco breeding programs were assessed through AFLP analysis. Polymorphic profiles were obtained with four selective primers combinations (EcoR I/Mse I) among the studied accessions. A total of 203 polymorphic bands (57.79 %) were used for cluster analysis (UPGMA) based on genetic similarity and genetic distance matrices. A common group was detected, comprising all the cultivated varieties that showed high genetic similarity (0.87446-0.93920) according to the Nei and Li’s distance measure, whereas wild species showed the highest genetic diversity. It was also possible to identify some bands shared among cultivated accession as specific markers for the analyzed Cuban cultivated tobacco. Our results indicate that AFLP analysis effectively detects the genetic diversity at levels enough to differentiate wild species of Nicotiana from Cuban varieties of N. tabacum, even though a narrow genetic diversity was present in Cuban varieties. All the accessions were distinguished through AFLP analysis.
Keywords: Genetic diversity, molecular markers, Nicotiana, polymorphism, tobacco.
RESUMEN
Las especies salvajes de Nicotiana se utilizan comúnmente en los programas de mejoramiento genético, para obtener cultivares con nuevas características que lleven al incremento de la productividad, y a la tolerancia o resistencia a enfermedades o a diferentes tipos de estrés. En Cuba, la producción de tabaco es una de las principales fuentes de ingreso económico, de ahí que en la pasada década se obtuvieran nuevas variedades del cultivo. En este estudio se analizó el polimorfismo genético de cuatro variedades cubanas de Nicotiana tabacum L. y seis especies salvajes mediante AFLP, dichas variedades empleadas en los programas de mejoramiento genético. Se utilizaron cuatro combinaciones selectivas de cebadores (EcoR I/Mse I) (EcoR I: E-ACT combinado con Mse I: M-CAC; M-CAG, M-CTC y M-CAT) para generar los perfiles polimórficos entre los genotipos estudiados. Se obtuvo un total de 203 bandas polimórficas (57.79 %), suficientes para el análisis de clúster (UPGMA) basado en similitudes genéticas y matrices de distancias genéticas. Esto permitió la distinción de un grupo común que comprende todas las variedades analizadas, los que mostraron una alta similitud genética (0.87446-0.93920) mediante la medida de distancia de Nei y Li, mientras las especies salvajes mostraron la más alta diversidad genética. También fue posible identificar algunas bandas similares como marcadores específicos para las variedades cultivadas de tabaco cubano analizadas. Nuestros estudios indican que existe suficiente diversidad genética entre las variedades cubanas de N. tabacum estudiadas y las especies salvajes como para diferenciarlas, y que, a su vez, la diversidad genética entre las variedades cubanas es estrecha. El análisis mediante AFLP permitió distinguir a todos los genotipos estudiados.
Palabras clave: Diversidad genética, marcadores moleculares, Nicotiana, polimorfismo, tabaco.
INTRODUCTION
The genus Nicotiana L. is the fifth largest genus within Solanaceae, including up to 75 species [1], but only two of them are cultivated (Nicotiana tabacum L. and Nicotiana rustica L.) which are commonly known as tobacco [2]. It is known that in the early evolution of a genus, related species share close genetic affinity and gene exchanges are frequent. Chromosome number in Nicotiana is very variable; according to Goodspeed [3], the basic chromosome number is n = 6, but further studies have revealed variations from n = 9 to n = 32, with some species regarded as amphidiploids or polyploids (n = 24) which arose from interspecific hybridization (allopolyploidy). And this is the case of N. tabacum and its varieties. In fact, genetic relationships have been found for Nicotiana species [4, 5], demonstrating that genetic processes and geographic isolation have driven important changes in genome organization and divergence in nuclear DNA sequences, which resulted in the evolution and diversification of the genus.
Particularly, Cuban tobacco is famous worldwide and almost the whole production is exported as cigars [6], with large plantations in the country and being an economically relevant crop. This has fostered the development of new varieties and hybrids in an attempt to minimize the damage caused by diseases, environmental stress and to increase production yields [7, 8]. The main varieties of tobacco grown in Cuba are Corojo and Criollo, although some new hybrids have been introduced due to its resistance to some typical tobacco diseases as the blue mold, with Habana-92, Habana 2000 and Burley Habana 13 (BH-13) among them [9, 10].
Due to the importance of tobacco for Cuban economy, some studies had focused on the analysis of genetic diversity and relationships among Nicotiana germplasm based on isozymes variation [4] and randomly amplified polymorphic DNA (RAPD) analysis [5]. However, Cuban cultivated varieties remain to be studied. According to Julio et al. [11], the few cultivars resistant to tobacco’s diseases are consistent with the low genetic diversity found within N. tabacum. On the other hand, several studies have described genetic diversity among wild and cultivated Nicotiana [12] or just among cultivars [13, 14] using amplified fragment length polymorphism (AFLP) and randomly amplified polymorphic DNA (RAPD), but just a poor sampling of Cuban varieties have been included in such studies.
It is of general knowledge that N. tabacum varieties have low genetic diversity [12-14]. Therefore, a better knowing of the available germplasm of Cuban tobacco will contribute to its preservation. This will also aid for better planning of Cuban tobacco breeding programs, especially to select the most appropriate parental genetic background able to provide the most advantageous genetic properties of cultivated varieties and wild species. Hence, this work was aimed to analyze the genetic polymorphism in plants of six wild Nicotiana species and in four cultivated varieties of N. tabacum germplasm used for tobacco breeding in Cuba. The cultivated varieties are among the most cultivated ones. Their germplasms were screened through amplified fragment length polymorphism (AFLP) analysis.
MATERIALS AND METHODS
Plant material and DNA extraction
Six wild species of Nicotiana and four Cuban varieties of N. tabacum (Habana 2000, Corojo 99, Criollo 98, Burley Habana 13) were studied (Table 1). Plant material was provided by the Tobacco Research Institute (IIT; Havana, Cuba). Samples of 5 g of fresh leaves from each species and variety were processed and the respective total genomic DNA was extracted by the CTAB method [15], being further purified as described by Ausubel et al. [16]. The integrity and quality of the DNA obtained was verified by electrophoresis in 1.8 % agarose gels.
AFLP analysis
AFLP analysis was done following the standard procedure described by Vos [17] and using the IRDye® Fluorescent AFLP Kit® for large plant genome analysis (LI-COR® Biosciences, USA) as recommended by the manufacturer. High-quality DNA was digested with a pair of restriction enzymes (EcoR I/Mse I) and then ligated to adapters. The assembled fragments were pre-amplified with non-selective primers EcoR I (5´-GACTGCGTACCAATTCA-3´) and Mse I (5´- GATGAGTCCTGAGTAAC-3´). Subsequently, four selective primer sets chosen to generate the AFLP profiles: EcoRI + ACT/Mse I + CAC (E-ACT/M-CAC); EcoR I + ACT/Mse I + CAG (E-ACT/M-CAG); EcoR I + ACT/Mse I + CTC (E-ACT/M-CTC); EcoRI + ACT/Mse I + CAT (E-ACT/M-CAT) with EcoR I fluorescent labeled primers. Amplification products were separated in 6.5 % polyacrylamide gels using IRDye® 800-labeled molecular weight marker (LI-COR®) as a size standard (50-700 bp). Band patterns were detected using a LI-COR® DNA Analyzer model 4200 and photographed with SAGAMX (LI-COR®).
Data analysis
The patterns of the amplified fragments were analyzed and scored as discrete characters states (presence/absence of bands: +/–) with the software SAGAMX (LI-COR®) to produce a binary matrix from primary data. A matrix of genetic similarity was calculated using the metric of Nei and Li [18]. Additionally, a matrix of genetic distances [19] was generated by the Simple Matching coefficient [20] between genotypes based on the number of shared bands, and further subjected to cluster analysis in FreeTree [21]. A dendrogram was constructed based on the un-weighted pair-group method with arithmetic averages (UPGMA). For the validation of particular branches, bootstrap analysis [22] was carried out using 1000 replicates.
RESULTS
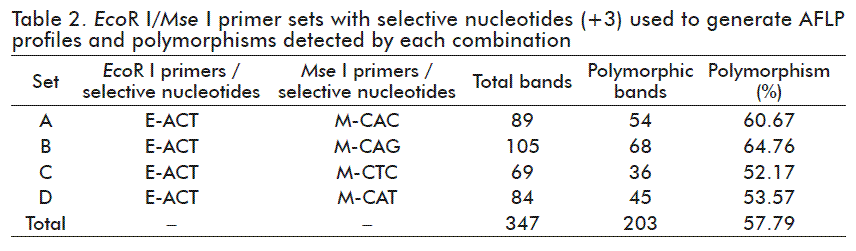
AFLP profiles revealed a large number of bands per primer combination which gave fragments of different size within the range of 200-700 bp. A total of 347 amplification bands were obtained for the whole set of primers, of which 57.79 % were polymorphic (203). High levels of polymorphisms were also detected for each primer pair separately (Table 2). The maximum number of polymorphic fragments (68 bands, 64.76 %) was amplified by the primer pair E-ACT/M-CAG (Figure 1) whereas the minimum (36, 52.17 %) was obtained with the pair E-ACT/M-CTC. Since all primer pair’s amplifications resulted in more than 50 % polymorphisms, these results demonstrated the usefulness of these sets for diversity analysis in Nicotiana spp.
Several fragments obtained by each primer combination were shared exclusively among N. tabacum accessions (Figure 1); therefore, they were identified as species’ specific markers.
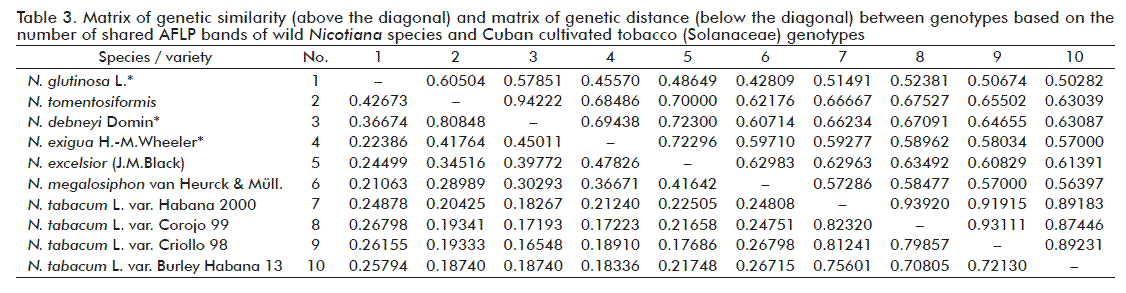
Based on the 203 polymorphic fragments, pairwise comparison among the ten accessions studied revealed genetic similarities from 0.42809 to 0.94222; whereas genetic distance ranged from 0.16548 to 0.82320 (Table 3).
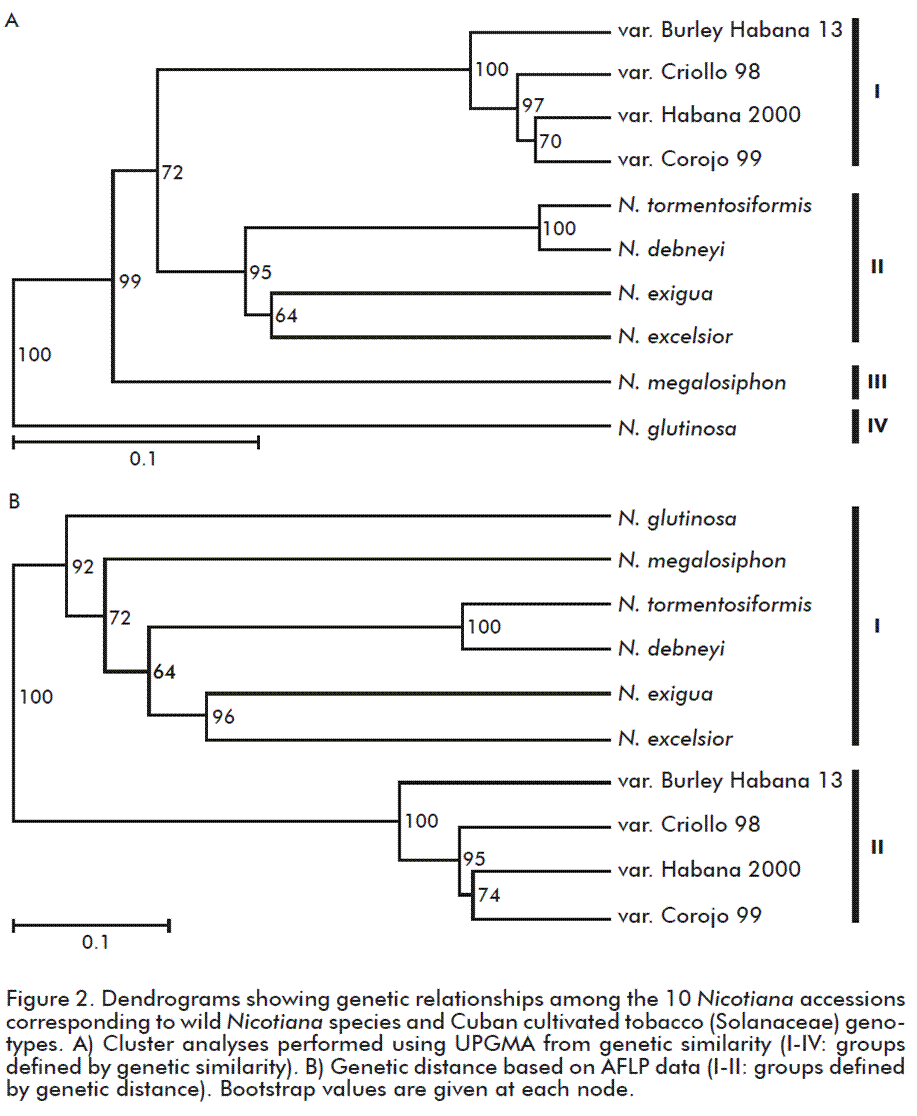
The results of cluster analyses varied according to the measure used. Based on genetic similarity, all accessions were grouped into four main groups (Figure 2 A). Attending to genetic distance analysis, accessions were separated into two main groups (Figure 2 B). Nevertheless, all cultivars of N. tabacum remained separated from wild species since they were clearly grouped together with maximum bootstrap support in both analyses, regardless the measure used for cluster analysis (Figure 2). On the other hand, the greatest differences were observed among wild species, their grouping patterns are in disagreement with their geographic origin or phylogenetic relationships previously described.
DISCUSSION
The results concern the reliability of the four primer combinations used for assessing genetic diversity in the genus Nicotiana since all sets provided sufficient number of polymorphic fragments (> 50 %) to differentiate among studied accessions (Table 2). Similar results were previously obtained with other combinations of EcoR I/Mse I primers, which found higher levels of polymorphism in Nicotiana than those obtained with other molecular markers such as Sequence-Specific Amplification Polymorphism (SSAP) [23] and RAPD [24].
The highest diversity rates were found among wild species of Nicotiana (Table 3) which is consistent with previous interspecific AFLP variation, as reported by Ren et al. [12]. As suggested by Mehrotra and Goyal [25], nuclear DNA in Nicotiana has experienced several reorganizations during the evolution and diversification of the genus, supporting the high levels of polymorphism observed among wild species. Phylogenetic reconstructions based on plastid and nuclear DNA data have established that the genus Nicotiana is monophyletic and supports the understanding of species relationships and patterns of divergence according to geographical distribution [1, 26, 27]. Besides, in this study it was not possible to recognize a congruent pattern between genetic diversity as revealed by AFLP and geographic origin of the analyzed species. N. megalosiphon showed genetic affinity with N. glutinosa meanwhile N. tomentosiformis showed affinity with other Australian species (Figure 2). Most of the presumed relationships between diploid and polyploidy Nicotiana species have been confirmed using molecular cytogenetics and mapping of repetitive DNA [28, 29]. The basic chromosome number of the genus is n = 6, but more than 30 species (including the entire Australian section Suaveolentes) are allopolyploids, although some species in section Suaveolentes show secondary chromosome number reductions in the range of 32 to 48 [3; 27], such as some of the species from this section analyzed (Table 1).
Regarding N. tabacum, it is known that about 77 % of the genome is made up of repetitive DNA sequences [30] which are regarded as responsible for most variations among cultivars. The high genetic similarity observed among cultivated accessions (0.87446-0.93920, Table 3) may be related to inbreeding in Cuban tobacco germplasm, due to the aim of maintaining the desired agronomic and quality traits during breeding programs. Some studies have pointed out low levels of genetic diversity among Nicotiana species [31], mainly on cultivated plants. Zhang et al. [24] selected 28 accessions with similar agronomic characteristics for an AFLP study. They used 14 selective primers to screen against all 28 accessions and it resulted in 154 polymorphic fragments with an average of 15.4 (27.45 %) per primer pair. They also found that high similarity measures revealed by AFLP in cultivars, which shares gene pools, were linked to morphological similarities. Such low genetic diversity detected by AFLP is common among cultivated plants as reported for acid lime [32], soybean [33] and others.
Regarding the cultivated varieties, the presence of amplification fragments of similar size shared exclusively among the four cultivated accessions (Figure 1) may be useful as species’ specific markers. They also might be related to inbreeding processes induced by the use of a limited gene pool in Cuban tobacco germplasm.
Specifically for the AFLP analysis, it is highly efficiently compared to morphological analyses and other molecular markers since it covers the entire genome [34]. Other groups had used it to study genetic polymorphism in tobacco as we implemented it. For instance, Ren and Timko [12] recommended five combinations, because they were suitable to detect genetic polymorphism between cultivars (46 lines of N. tabacum) and seven wild species (N. sylvestris, N. tomentosiformis, N. otophora, N. glutinosa, N. suaveolens, N. rustica, and N. longiflora). These authors recommended the use of the combinations E-ACT + M-CAG and E-ACT + M-CAC which were able to detect the highest percentage of polymorphism in the genotypes evaluated. Denduangboripant et al. [35] reported a similar outcome; the highest percentage of polymorphism (87 %) found among 19 tobacco cultivars by using the combination E-ACT + M-CAG. This precedent, together with these results showing the maximum number of polymorphic fragments with the E-ACT + M-CAG combination, lead us to propose that these AFLP markers could be converted into Sequence Characterized Amplified Region (SCAR), specific locus markers, prior confirmation that they can be used for the identification of cultivars and the studied species. SCAR markers are simpler and less expensive to use for cultivar identification studies than AFLP markers.
On the other hand, the lowest percentage of polymorphism was obtained with the E-ACT + M-CTC combination, in agreement with reports by Zhang et al. [24] on the obtention of just monomorphic amplification products using this combination. Dadras et al. [36] also used AFLP markers to analyze 50 N. tabacum genotypes with 21 primer combinations, and high polymorphic rates varying from 52.63 to 92.59 % were found, demonstrating that the AFLP technique can be a powerful and valuable tool in the breeding program of N. tabacum. Overall, these findings indicate that AFLP analysis effectively detects the genetic diversity enough to differentiate wild species of Nicotiana from Cuban varieties of N. tabacum. Moreover, they also remark the usefulness of AFLP for studying the polymorphism of tobacco genome.
Apart from the narrow genetic diversity present in Cuban varieties, all accessions were identified by the AFLP analysis. The reduced genetic diversity of the Cuban varieties represents an important knowledge for further research on cultivar identification through the utilization of molecular markers. It also emphasizes on the need to focus on breeding programs improvement by introducing more diverse cultivated tobacco or using desirable traits found in wild species for cultivated plants.
ACKNOWLEDGEMENTS
We thank Yosvanis Acanda Artiga and Humberto García Cruz for providing samples of Cuban tobacco germplasm bank. We are also grateful to Katia Gil-Vega from CINVESTAV (Mexico) and Miriam Isidrón Pérez from the Agrarian University of Havana (Cuba) for their assistance with the analyses, and Vitor Fernandes Oliveira de Miranda (Brazil) for comments on the early manuscript.
CONFLICTS OF INTEREST STATEMENT
The authors declare that there are no conflicts of interest.
REFERENCES
1. Clarkson JJ, Knapp S, Garcia VF, Olmstead RG, Leitch AR, Chase MW. Phylogenetic relationships in Nicotiana (Solanaceae) inferred from multiple plastid DNA regions. Mol Phylogenet Evol. 2004;33(1):75-90.
2. Gerstel DU. Evolutionary problems in some polyploid crop plants. Hereditas. 1966;2:481-504.
3. Goodspeed TH. The genus Nicotiana. Chronica Botanica. 1954;16:1-536.
4. Valdés M, González C, Lara RM, Román MI, Hernández Y, Hernández RM, Cabrera M, et al. Diversidad genética de especies silvestres del género Nicotiana I: caracterización mediante marcadores bioquímicos. Rev Protección Veg. 2010;25:88-97.
5. Valdés M, Hernández Y, Vázquez A, Piñero D, González L. Diversidad genética de especies silvestres del género Nicotiana II: caracterización molecular mediante marcadores RAPD. Rev Protección Veg. 2010;25:166-73.
6. Pérez S, Cabezas D, Domínguez Y, Coto O, García H. Evaluación genómica de los genes pr1 y tctp en especies y variedades detabaco (Nicotiana tabacum L.). Rev Protección Veg. 2011;26:164-9.
7. García V, Santana N, García H, Mena E. ‘Corojo 2006’: nueva variedad cubana de tabaco negro (Nicotiana tabacum L.). Cult Trop. 2013;34(2):45.
8. García V, Santana N, Mena E. ‘Burley Pinar 2010’: primer cultivar cubano de tabaco Burley resistente al marchitamiento por Fusarium (Fusarium oxysporum) y a otras enfermedades de importancia en el cultivo. Cult Trop. 2015;36(2):91.
9. García H, García V, Pino LA, Espino E. ‘Burley Habana-13’ primera variedad comercial obtenida en Cuba por cultivo de anteras con resistencia al moho azul (Peronospora tabacina), pata prieta (Phytophtora parasitica) y al virus del mosaico del tabaco (VMT). Cubatabaco. 1999;1:49-54.
10. Espino E. Resultados de los experimentos de regionalización de variedades (2001-2002). Cubatabaco. 2003;4:61-76.
11. Julio E, Verrier JL, Dorlhac de Borne F. Development of SCAR markers linked tothree disease resistances based on AFLP within Nicotiana tabacum L. Theor Appl Genet. 2006;112(2):335-46.
12. Ren N, Timko MP. AFLP analysis of genetic polymorphism and evolutionary relationships among cultivated and wild Nicotiana species. Genome. 2001;44(4):559-71.
13. Zhang HY, Liu XZ, Li TS, Yang YM. Genetic diversity among flue-cured tobacco (Nicotiana tabacum L.) revealed by amplified fragment length polymorphism. Bot Stud. 2006;47:223-9.
14. Sarala K, Rao RVS. Genetic diversity in Indian FCV and burley tobacco cultivars. J Genet 2008;87(2):159-63.
15. Doyle JJ, Doyle JL. A rapid DNA isolation procedure for small quantities of fresh leaf tissue. Phytochem Bull. 1987;19:11-5.
16. Ausubel FM, Brent R, Kingston RE, Moore DD, Seidman JG, Smith JA, et al. Short protocols in molecular biology: a compendium of methods from Current Protocols in Molecular Biology. 5th ed. New York, NY, USA: Wiley; 2002. p. 250.
17. Vos P, Hogers R, Bleeker M, Reijans M, Lee TVD, Hornes M, et al. AFLP: a new technique for DNA fingerprinting. Nucleic Acids Res. 1995;23:4407-14.
18. Nei M, Li WH. Mathematical model for studying genetic variation in terms of restriction endonucleases. Proc Natl Acad Sci USA.1979;76:5269-73.
19. Sokal RR. Principles and practices of numerical taxonomy. Taxon. 1963;2(5):190-9.
20. Skroch P, Tivang J, Nienhuis J. Analysis of genetic relationships using RAPD markerdata. In: The American Society for Horticultural Science and American Genetic Association, editors. Applications of RAPD technology to plant breeding. Joint plant breeding symposia series. Minneapolis: Crop Science Society of America; 1992. p. 26-30.
21. Hampl V, Pavlícek J, Flegr J. Construction and bootstrap analysis of DNA fingerprinting-based phylogenetic trees with the freeware program FreeTree: application to trichomonad parasites. Int J Syst Evol Micr. 2001;51:731-5.
22. Felsenstein J. Confidence Limits on Phylogenies: An Approach Using the Bootstrap. Evolution. 1985;39(4):783-91.
23. Petit M, Lim KY, Julio E, Poncet C, Dorlhacde Borne F, Kovarik A, et al. Differential impact of retrotransposon populations on the genome of allotetraploid tobacco (Nicotiana tabacum). Mol Genet Genomics. 2007;278(1):1-15.
24. Zhang HY, Liu XZ, He CS, Yang YM. Genetic diversity among flue-cured tobacco cultivars based on RAPD and AFLP markers. Braz Arch Biol Techn. 2008;51:1097-1101.
25. Mehrotra S, Goyal V. Repetitive sequences in plant nuclear DNA: types, distribution, evolution and function. Genomics Proteomics Bioinformatics. 2014;12(4):164-71.
26. Chase MW, Knapp S, Cox AV, ClarksonJJ, Butsko Y, Joseph J, et al. Molecular systematics, GISH and the origin of hybridtaxa in Nicotiana (Solanaceae). Ann Bot. 2003;92(1):107-27.
27. Knapp S, Chase MW, Clarkson JJ. Nomenclatural changes and a new sectional classification in Nicotiana (Solanaceae). Taxon. 2004;53:73-82.
28. Lim KY, Matyasek R, Kovarik A, FulnecekJ, Leitch AR. Molecular cytogenetics and tandem repeat sequence evolution in the allopolyploid Nicotiana rustica compared with diploid progenitors N. paniculata and N. undulata. Cytogenet Genome Res. 2005;109(1-3):298-309.
29. Lim KY, Kovarik A, Matyáçek R, Chase MW, Knapp S, McCarthy E, et al. Comparative genomics and repetitive sequence divergence in the species of diploid Nicotiana section Alatae. Plant J. 2006;48:907-19.
30. Renny-Byfield S, Chester M, Kovarik A, Le Comber SC, Grandbastien MA, Deloger M, et al. Next generation sequencing reveals genome downsizing in allotetraploid Nicotiana tabacum, predominantly through the elimination of paternally derived repetitive DNAs. Mol Biol Evol. 2011;28(10):2843-54.
31. Darvishzadeh R, Gholizadeh S, Hatami-Maleki H, Abdollahi B, Bernousi I. Study on genetic diversity among Iranian water pipe’s tobacco (Nicotiana spp.) varieties by using simple sequence repeat markers. Bulgarian J Agr Sci. 2013;19(3):557-62.
32. Al-Sadi AM, Al-Moqbali HS, Al-Yahyai RA, Al-Said FA. AFLP data suggest a potential role for the low genetic diversity of acid lime (Citrus auriantifolia Swingle) in Oman in the outbreak of witches’ broom disease of lime. Euphytica. 2012;188:285-97.
33. Kondetti P, Jawali N, Apte SK, Shitole MG. Comparative study of genetic diversity in Indian soybean (Glycine max L. Merr.) by AP-PCR and AFLP. Ann Biol Res. 2012;3:3825-37.
34. Zhang Z, van Parijs FR, Xiao B. The status of AFLP in the genomics era and a pipeline for converting AFLPs into single-locus markers. Mol Breeding. 2014;34:1245-60.
35. Denduangboripant J, Piteekan T, Nantharat M. Genetic polymorphism between tobacco cultivar-groups revealed by amplified fragment length polymorphism analysis. J Agri Sci. 2010;2:41-8.
36. Dadras AR, Sabouri H, Nejad GM, Sabouri A, Shoai-Deylami M. Association analysis, genetic diversity and structure analysis of tobacco based on AFLP markers. Mol Biol Rep. 2014;41(5):3317-29.
Received in March, 2018.
Accepted in June, 2018.
Sandra Peréz-Álvarez. Departamento de Biotecnología Agrícola, Unidad Sinaloa, Instituto Politécnico Nacional, CIIDIR-IPN. Blvd. Juan de Dios Bátiz Paredes No 250. Guasave, Sinaloa, CP 81101, México. E-mail: perezalvarezsandra2015@gmail.com.


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}