










 Servicios personalizados
Servicios personalizados Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCTION
The main species of small fruits produced worldwide are the strawberry, the blueberry and the raspberry (Faostat, 2017). Within this group, the strawberries is the one with the greatest economic expression, being appreciated in several regions of the world (Oliveira et al., 2011). The emphasis on the production of the species is based not only on the economic return, but also, as an important source of vitamin C, anthocyanins and antioxidant activities, which can contribute to disease prevention (Cocco et al., 2015).
The production and use of healthy seedlings are the most important factors for obtaining high quality fruits and better responses to the technologies used (Oliveira and Scivittaro, 2009). Thus, tissue culture techniques allow the production of a large number of plants with excellent phytosanitary and genetic quality in a shorter period of time, through several cycles of in vitro multiplication (Fonseca et al., 2013). The use of the micropropagation technique makes it possible to meet the demands of the consumer market by offering high quality seedlings to nurseries and homogeneous seedlings to farmers (Dias et al., 2014).
The culture media composition has often been modified to stimulate the growth and development of specific plant material (Sivanesan and Park, 2014). Among the nutritional components, silicon is considered an element that has not been much studied in the plant micropropagation. However, from the physiological point of view, for plants growth and development, silicon has shown beneficial effect on the increase the production of several crops (Gomes et al., 2008). Culture medium supplemented with silicon can benefit seedlings in vitro cultured, raising the hemicellulose and lignin content and thus increasing the stiffness in the cell wall (Camargo et al., 2007).
Thick cell wall presence is a mechanism of preformed structural resistance of hosts and may contribute to the restriction of plant colonization by phytopathogens (Pascholati, 2011). With this, silicon has provided promising results in the control of plant diseases (Domiciano et al., 2010; Lima et al., 2010).
This paper aimed to determine the effect of potassium silicate added to the MS medium (Murashige and Skoog, 1962) in the strawberry in vitro propagation.
MATERIAL AND METHODS
Trial was carried out at the Laboratory of Plant Micropropagation of Santa Catarina State University (UDESC / CAV) in Lages city, Santa Catarina State (SC), Brazil, in 2016.
Plant material
Strawberry seedlings originating from the regeneration of shoot apex in vitro and grown in MS medium were used as plant material.
In vitro culture
Plantlets approximately 1.5 ± 0.5 cm length were introduced into glass bottles containing 30 ml of the same medium MS with potassium silicate at five different concentrations (0.0, 0.5, 1.0, 1.0 , 5.0 and 2.0 g l-1). The basal medium was composed of 30 g l-1 sucrose, 0.1 g l-1 myo-inositol and 0.5 g l-1 6-benzylaminopurine (BAP) and the pH was adjusted to 5.8 before addition of 6 g l-1 of the solidifying agent (agar). Then, the vials containing the culture medium were sterilized by autoclave at 121 °C and 1.2 atm for 20 minutes.
In a laminar flow chamber, seedlings were introduced into the culture medium and then transferred to a growth room at 25 °C (± 2 °C), photoperiod of 16 hours and light intensity of 27 μmol m-2 s-1 supplied by white fluorescent lamps.
After 45 days of culture, the following variables were evaluated:
a) number of shoots: obtained by separation and manual counting of shoots per explant
b) number of leaves: obtained by the separation and manual counting of leaves per explant
c) shoot length: measured from the neck of the plant to the upper end of the largest leaf, expressed in centimeters
d) SPAD index: quantified using the Minolta® SPAD-502-PLUS portable meter, which performs a sheet absorbance measurement in two regions of wavelength (red and near infrared regions) and from these two transmittances, the equipment calculates the SPAD value proportional to the amount of chlorophyll present in the leaf
e) number of roots: obtained by separation and manual counting of roots per plant, and
f) root length: measured from the neck of the plant to the end of all roots, expressed in centimeters.
Statistical analysis
Data were analysed for normality by the Shapiro-Willk test. Values that were not normal were transformed into √𝑥+ 0.1, where 𝑥 is the average obtained from each variable. They were then submitted to variance analysis and when significant by the F test, regression analysis was performed. In addition, Pearson correlation analysis was performed between the variables number of roots per explant and average length of shoots; also average length of roots and average length of shoots. The statistical program used for data processing was Sisvar (5.6 version).
RESULTS AND DISCUSSION
The concentrations of potassium silicate added to the culture medium had a negative influence on the number of shoots, mean length of shoots and number of leaves (Figure 1).
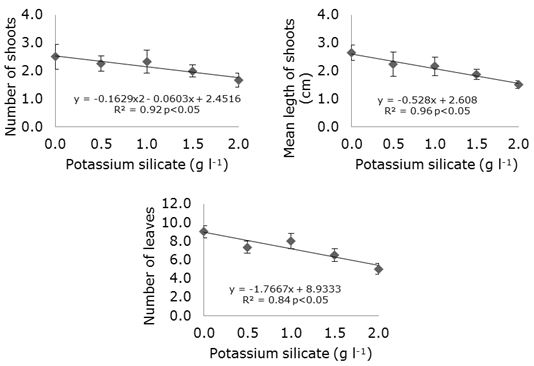
Figure 1. Number of shoots, mean shoot length (cm) and number of leaves as a function of the addition of different concentrations of potassium silicate to MS medium in strawberry cv. ‘Jonica’ in vitro propagation.
In the treatment with 2 g l-1 potassium silicate there was an approximately 80% reduction in the number of leaves when compared to the treatment without silicon. As observed in this study, the addition of potassium silicate to the MS culture medium also resulted in a lower number of leaves in the explants of strawberry cultivar ‘Oso Grande’ (Braga et al., 2009). Besides, studying two species of orchids (Brassavola perrine and Laelia cattleya Culminant ‘Tuilerie’ x Laelia cattleya Sons Atout Rotunda x Brassolaelia cattleya Startifire Moon Beach) Pasqual et al. (2011) reported that the number of leaves decreased with increasing concentrations of calcium silicate added to the culture medium. In the same way, Alves et al. (2017), verified that the increase of the concentrations of calcium silicate caused a reduction in the leaf area and in the height of the orchid aerial part.
According to Soares et al. (2008), the reduction in the number of leaves of plants that receive silicon may be a mechanism to reduce the perspiration. However, Soares et al. (2011) found greater numbers of leaves in Cattleya loddigesii when the specie was grown in Knudson C medium supplemented with 20 mg l-1 calcium silicate and 20 mg l-1 potassium silicate. The adjustment of the osmotic potential in plant cells is regulated by potassium, which also activates enzymes involved in respiration and photosynthesis (Taiz and Zeiger, 2009), so it is possible to attribute the increased leaf growth observed in this study to potassium and not just to silicon.
The absence of responses in relation to the analysed variables can be related to the determination of the optimum concentration and the silicon source used that varies according to the species or plant genotype (Soares et al., 2013). Another explanation is that nutritional excess, as in this case potassium, may have caused an imbalance leading to deficiencies or overaccumulation of nutrients (Malavolta et al., 2006). Moreover, the negative results obtained with the addition of potassium silicate can be attributed to an unfavourable interaction between this and the other components of the medium. In this sense, according to Borgatto et al. (2002), in vitro rowth of plants, organs, tissues and/or cells depends on the perfect interaction between the essential components of the culture medium, such as carbon sources and mineral nutrients, which are limiting factors for in vitro development.
The SPAD index had a polynomial regression curve response, where the inflection point was at 0.67 g l-1 of potassium silicate added to the nutrient medium (Figure 2).
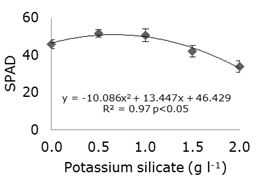
Figure 2. SPAD index as a function of the addition of different potassium silicate concentrations to the MS culture medium in strawberry cv. ‘Jonica’ in vitro propagation.
The inclusion of 0.67 g l-1 of potassium silicate in the culture medium increased the chlorophyll levels, in the 'Jonica' strawberry seedlings, which are the green pigments specialized in light absorption (Taiz et al., 2017). According to Takahashi (1995), the best foliar architecture provided by silicon allows greater penetration of light, greater absorption of CO2 and decrease of excessive transpiration, which causes the increase of the photosynthetic rates. The beneficial action of silicon has been associated with several indirect effects, such as increased photosynthetic efficiency and chlorophyll content (Sivanesan and Park, 2014).
The linear regressions for the variables related to the root system of strawberry cv. 'Jonica' explants (Figure 3) indicated that the Potassium silicate concentrations added to the culture medium did not contribute to the increase of the variables analysed.
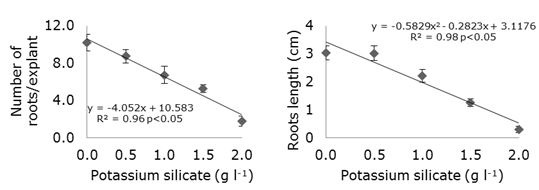
Figure 3. Number of roots and mean root length (cm) as a function of the addition of different potassium silicate concentrations to the MS culture medium in strawberry cv. ‘Jonica’ in vitro propagation.
When the number of roots formed in vitro is increased, the root/culture medium contact area is also increased, reflecting a higher nutrients absorption (Soares et al., 2013). Thus, in the control treatment it was observed that the root number and mean root length were superior related to the other treatment (the control do not increase, just it was not affected by the silicate added), and the addition of any concentrations of potassium silicate decreased the means for these variables. A similar result was observed in coffee (Coffea arabica L.), where the addition of potassium silicate to the nutrient solution did not present promising results, since there was a decrease in the dry mass of the roots (Cunha et al., 2012).
Potassium silicate effects on in vitro root development of strawberry cv. 'Jonica' showed that under the conditions in which this experiment was performed, the number and length of roots decreased with increasing potassium silicate concentrations, indicating that potassium may have been over-offered. According to Malavolta et al. (1997) potassium is involved in the meristematic growth, since the phytohormones acting in this process are put into action by this mineral. When in excess it can inhibit the absorption of other ions, like Calcium and Magnesium, essential for the development of the roots. The lack of Magnesium would prevent the complete development of the roots and the lack of Calcium, besides limiting the growth of the roots leaves them with a darker colour (Nunes, 2016).
A trial carried out with the banana variety ‘Apple’ (Musa acuminata) shows that there was no significant effect of adding different sources of silicon to the MS culture medium on root mean length compared to the control treatment (Asmar et al., 2011). Besides, silicate sources did not affect the in vitro development of orchids (Cattleya forbesii) and there was a decrease in the number and average root length of the plants (Colombo et al., 2016), in agreement with the results obtained in the present trial. The inhibitory effect on root formation due to the addition of potassium silicate to the nutrient medium may be associated with the concentration itself (George et al., 2008), since all nutrients in excess cause a nutritional imbalance in both the seedling and the culture medium (Soares et al., 2013).
In studies of mineral nutrition, it is necessary to consider nutrients as a whole, because in the absorption process, one can exert influence on the other, given the possible interactions that may occur (Malavolta et al., 1997). It is possible that the potassium silicate concentrations caused a nutritional imbalance in the plants inhibiting the nutrients absorption from the culture medium, thus, silicon could not exert beneficial effects on the growth and development of strawberry seedlings.
As roots are responsible for the uptake of water and nutrients, lower emission and lower growth of the root system generally reflect negatively on shoot growth rate (Colombo et al., 2016). This response was assessed because the reduced development of the aerial part was correlated with the number of roots (r = 0.85) and with the average length of roots (r = 0.83) (Figure 4).
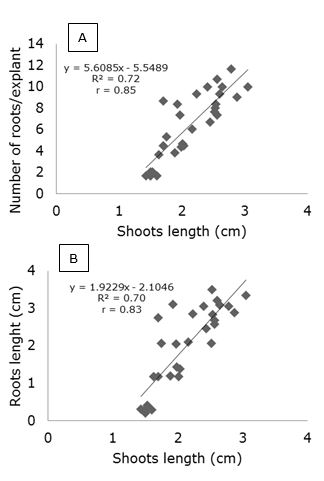
Figure 4. Pearson correlation between the number of roots per explant and the mean of shoots length (A) and between the mean root length and the mean shoot length (B) in strawberry cv. 'Jonica' explants culture with different potassium silicate concentrations in the in vitro medium culture.
In vitro growth of seedlings depends on the optimization of culture media that interact perfectly with essential components such as carbon source, mineral nutrients and growth regulators (Soares et al., 2013). The ideal silicon concentration required for plant growth and development varies between different genotypes within the same species and among plant species (Lim et al., 2012). Due to the lack of data on the influence of silicon on the strawberry micropropagation it is necessary to carry out future trials in order to fill this deficiency of information. Based on the results obtained through this trial, it is not recommended to use potassium silicate for ‘Jonica’ strawberry in vitro multiplication.

