










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión On-line ISSN 2224-4697
Rev. Protección Veg. v.26 n.1 La Habana ene.-abr. 2011
TRABAJO ORIGINAL
CARACTERIZACIÓN molecular de AISLAMIENTOS DE Cladosporium fulvum Cooke PROVENIENTES DE TOMATE EN CONDICIONES DE CULTIVO PROTEGIDO
MOLECULAR CHARACTERIZATION OF Cladosporium fulvum Cooke ISOLATES FROM TOMATO IN SHELTERED CROP CONDITIONS
Belkis Peteira*, A. Bernal Cabrera**, B. Martínez*, Ileana Miranda*
*Centro Nacional de Sanidad Agropecuaria, Autopista Nacional y Carretera de Tapaste, Apartado 10, San José de las Lajas, Mayabeque, Cuba. Correo electrónico: bpeteira@censa.edu.cu; **Facultad de Ciencias Agropecuarias, Universidad Central ¨Marta Abreu¨ de Las Villas, Carretera a Camajuaní. Km. 5½, Santa Clara, Villa Clara, Cuba
RESUMEN
El moho de la hoja, causado por el hongo Cladosporium fulvum Cooke, es una enfermedad muy común y destructiva en el cultivo del tomate. Aunque existen cultivares portadores de uno o más genes de resistencia, el hecho de que el hongo muta fácilmente trae como consecuencia un uso limitado de los mismos. De ahí que, el conocimiento de las poblaciones del patógeno contribuye a los esfuerzos realizados en el programa de mejoramiento y al desarrollo de estrategias que alarguen la vida útil de los materiales obtenidos. El objetivo de este trabajo fue la caracterización molecular de 37 aislados de C. fulvum provenientes de plantas de tomate de diferentes localidades. La caracterización incluyó la identificación de los aislamientos por la amplificación de la región 28S del gen que codifica para el ARNr, la PCR con cebadores específicos dirigidos a diferentes genes relacionados con la patogenicidad (Avr2, Avr4, Avr4E, Avr9, Ecp1, Ecp2, Ecp4 y Ecp5) y los RAPDs. Todos los aislados fueron clasificados como C. fulvum. Ningún aislado amplificó para el gen Avr9, aunque se encontraron elevados porcentajes de amplificación para los genes Avr2, Avr4 y Avr4E. La virulencia de las razas 2, 4, y 2.4 está más relacionada con la presencia de la proteína Ecp2 (100%) que con la Ecp1. Todos los marcadores empleados confirmaron un elevado nivel de variabilidad molecular, aunque en ninguno de los casos hubo relación con el origen geográfico.
Palabras clave: Cladosporium fulvum; caracterización molecular; genes Avr, genes Ecp, RAPDs.
ABSTRACT
The leaf mold caused by Cladosporium fulvum Cooke, is a very common and destructive disease in the tomato crop. Although there are cultivars with one or more resistance genes, the fact that the fungus has a very high mutation level, results in a limited use of such materials. For that reason, the knowledge of the pathogen populations contributes to the breeding program efforts to develope strategies to extent the useful life of the materials obtained. The objective of this work was the molecular characterization of the variability in a group of C. fulvum isolates from tomato plants. The molecular characterization was carried out for 37 isolates, including identification by the 28S region encoding for RNAr amplification, PCR with specific primers directed to different genes related to pathogenicity (Avr2, Avr4, Avr4E, Avr9, Ecp1, Ecp2, Ecp4 y Ecp5) and RAPDs. All the isolates were classified as C. fulvum. Any of them amplified for the Avr9 gene, although there were high amplification percentages for the Avr2, Avr4 and Avr4E genes. The virulence of the races 2, 4, and 2.4 were more related to the presence of the Ecp2 protein (100%) than to Ecp1. All the markers used confirmed a high level of molecular variability, although no relation the geographical origin was found.
Key words: Cladosporium fulvum; molecular characterization; Avr genes, Ecp genes, RAPDs.
INTRODUCCIÓN
El tomate (Solanum lycopersicum, Mill), es una de las hortalizas de mayor distribución geográfica y consumo en el mundo. En Cuba, ocupa un lugar importante y con un desarrollo creciente por el incremento de las casas de cultivo protegido en todo el país.
Bajo estas condiciones, el moho de la hoja, causado por el hongo Cladosporium fulvum Cooke, es una enfermedad muy común y destructiva en el cultivo (1, 2) y ha llegado a convertirse, en nuestro país, en el principal problema fúngico que incide en estas instalaciones de cultivo protegido, sobre todo en la época de primavera-verano (3); aunque ocasionalmente aparece en campos de tomate en condiciones húmedas. En ausencia de medidas de control, grandes porciones de las hojas mueren resultando en una disminución significativa de los rendimientos (4). El manejo de la enfermedad se basa en algunas medidas de control cultural tales como adecuada ventilación del invernadero, control de la temperatura, evitar mojar las hojas durante la irrigación y un adecuado espacio entre plantas y surcos. Existen cultivares portadores de uno o más genes de resistencia, disponibles para su uso contra la enfermedad. Sin embargo, el hecho de que el hongo muta fácilmente trae como consecuencia un uso limitado de estos cultivares (4). De ahí que, el conocimiento de las poblaciones del patógeno contribuye a los esfuerzos realizados en el programa de mejoramiento y al desarrollo de estrategias que prolonguen la vida útil de los materiales obtenidos en el mismo.
La interacción entre C. fulvum y tomate es gobernada por la relación gen a gen. Diez proteínas efectoras de C. fulvum han sido caracterizadas, el mayor número para cualquier hongo filamentoso patógenos de plantas hasta el momento (1). Cuatro de estos efectores son proteínas de avirulencia razas específicas (Avr2, Avr4, Avr4E, y Avr9), seis son proteínas extracelulares (Ecp1, Ecp2, Ecp4, Ecp5, Ecp6, y Ecp7), y la mayoría de los genes correspondientes han sido clonados por diferentes autores (5, 6, 7).
El objetivo de este trabajo fue la caracterización molecular de un grupo de aislados de C. fulvum provenientes de plantas de tomate de cultivos protegidos ubicados en diferentes localidades del país.
MATERIALES Y MÉTODOS
La identidad de los aislamientos se comprobó a partir de la amplificación de la región 28S del gen que codifica para el ARNr mediante la técnica de Reacción en Cadena de la Polimerasa (PCR) y el empleo de cebadores específicos (8).
Para ello, se tomó un disco de micelio de 0.5cm de diámetro de cada uno de los aislados (Tabla 1), provenientes de Placas Petri contentivas de 10mL de medio PDA, 15 días después de la inoculación y se sembraron en erlenmeyers (250mL de capacidad) con 100 mL de medio de cultivo líquido B5 (9) suplementado con Sacarosa 2% (10). Los mismos se colocaron en agitación (zaranda orbital GFL) a 120 rpm y temperatura de 25±1ºC durante tres semanas. Transcurrido este período se colectaron los micelios por filtración y se conservaron en congelación (-20ºC) hasta su utilización. La extracción del ADN genómico total se realizó por el método de Moller et al. modificado por Peteira et al. (11). Las concentraciones de ADN se calcularon por la medición de la absorbancia a longitud de onda de 260nm en un espectrofotómetro (Ultrospec Plus Spectrophotometer Pharmacia, LKB), para el cálculo de la concentración según Sambrook et al. (12). Las muestras fueron conservadas a 4ºC hasta los diferentes análisis por PCR.
La mezcla de reacción de PCR empleada tuvo un volumen final de 25 µL compuesta por Tampon PCR 10X (Tris-HCl 10mM, KCl 50mM., MgCl2 1,5mM), dNTPs 200uM, 100ng de ADN, cebadores específicos 10 µM de cada uno (ARNr-F: 5´ GCT TAA GTT CAG CGG GTA TCC 3´ y ARNr-R: 5´ CGG CAA CGA CCA CCC AGG 3´) (13) y 1U de Taq polimerasa Amplicen (CENSA). Las amplificaciones fueron ejecutadas en un termociclador (PTC-100; MJ Research, Inc), desarrollando el programa siguiente: un paso de desnaturalización inicial a 94°C por 5 minutos, seguido por 29 ciclos de desnaturalización a 94°C (30 segundos), Hibridación a 55°C (30 segundos) y extensión a 72°C (1 minuto) y un paso de extensión final a 72°C por 7 minutos (13).
Para la amplificación de los genes Avr, se emplearon los cebadores específicos diseñados a partir de secuencias nucleotídicas publicadas en el NCBI (GenBank) para Avr2 (AJ421628), Avr4 (Y08356), Avr4E (AY546101), Avr9 (X60284) (Tabla 2).
El programa de PCR consistió de forma general en un paso de desnaturalización inicial a 94°C por 5 minutos, seguido por 34 ciclos de desnaturalización a 94°C (30 segundos), hibridación a 54°C (para Avr2), 49°C (para Avr4), 55°C (para Avr4E y Avr9) (30 segundos) y extensión a 72°C (1 minuto), concluyendo con un paso de extensión final a 72°C por 7 minutos (13).
Para la amplificación de los genes que codifican para las proteínas Ecp se emplearon los cebadores específicos diseñados a partir de secuencias nucleotídicas publicadas en el NCBI (GenBank) para Ecp1 (Z14023), Ecp2 (Z14024), Ecp4 (AJ271890) y Ecp5 (AJ271891) (Tabla 3).
El programa de amplificación consistió en este caso un paso de desnaturalización inicial a 94°C durante 5 minutos, seguido por 34 ciclos de desnaturalización por 94°C (30 segundos), hibridación a 54°C (para Ecp2 y Ecp1), 49°C (para Ecp4 y Ecp5) (30 segundos) y extensión a 72°C (1 minuto), con un paso de extensión final a 72°C por 7 minutos (13).
Para la amplificación con cebadores arbitrarios (RAPD) se emplearon cebadores de los Kit OPA y OPF (de la Firma Operon Technologies, Alameda), en concentración de 5 picomoles, en una mezcla de reacción idéntica a la antes descrita excepto en la concentración de MgCl2, que fue 2mM en este caso. La amplificación se ejecutó a través del programa siguiente: 45 ciclos de 1 minuto a 94ºC, 1 minuto a 36ºC y 2 minutos a 72ºC, y un ciclo de 10 minutos a 72ºC.
Todos los fragmentos amplificados fueron separados por electroforesis en gel de agarosa al 2% (excepto para RAPDs que fue en Agarosa al 1.5%), en TBE 0,5 X a 100 V y visualizados por tinción con Bromuro de Etidio (5mg/mL) con luz UV (transiluminador Pharmacia LKB). La talla del fragmento de ADN amplificado fue estimada por comparación con el marcador de peso molecular 1 Kb Ladder (Fermentas).
El aislamiento raza 0 (R-0), donado gentilmente por el laboratorio de Fitopatología de Universidad de Wageningen, se utilizó como control positivo en la identificación molecular de los aislados cubanos de C.fulvum y en las amplificaciones de los genes de avirulencia (Avr) y proteínas extracelulares (Ecp) del agente fitopatógeno. Como control interno se tomó la mezcla de reacción en la cual se sustituyó el ADN por agua destilada estéril.
Con las bandas obtenidas a partir de las amplificaciones de los genes de avirulencia, de los genes que codifican para las proteínas extracelulares (Ecps) y de los RAPDs de los aislamientos de C. fulvum, se confeccionaron matrices de datos binarios (una para cada análisis). Para la formación de los grupos se realizó un análisis de conglomerados jerárquico por el método de UPGMA (vinculación Inter. grupos) y se empleó la distancia de Nei y Li (14) para generar una matriz de coeficientes de similitud, la cual se utilizó para la construcción de los dendrogramas. La confirmación de la calidad de los grupos detectados se realizó con un análisis discriminante. Se empleó el paquete estadístico SPSS v. 11.5 para Windows (15).
RESULTADOS Y DISCUSIÓN
La comprobación de la identidad desde el punto de vista molecular se observa en la Fig. 1. En esta se aprecia el fragmento de ADN obtenido a partir de la amplificación de la región 28S del gen que codifica para el ARNr de los aislamientos de C. fulvum con el empleo de los cebadores específicos (8). La banda de aproximadamente 622 bp en todos los aislados evaluados, confirma la identidad de los aislamientos.
Las investigaciones que analizan como secuencia blanco la región ITS del ADNr de diferentes organismos han tenido diferentes aplicaciones también en el caso de hongos. En algunos estudios, los ITS han sido empleados para análisis de la diversidad por sí solos como los casos de Rhizoctonia (16), Phytophthora (17), Fusarium (18) y Beauveria (19). Gaskell et al. (20) utilizaron la secuencia de esta región para la discriminación entre los géneros Alternaria, Aspergillus, Cladosporium y Penicillium. En otros estudios, los ITS han sido combinados con otras tecnologías como secuenciación, PCR-RFLP o la PCR en tiempo real, los que apoyan la identificación y el diseño de cebadores género o especie específicos. La naturaleza hiperpolimórfica de la región de ITS2 entre las especies fúngicas, ya sea por longitud o secuencia nucleotídica provee una valiosa información para la determinación de especies. A partir de su empleo se ha logrado la identificación exitosa de muestras que habían fallado en su diagnóstico utilizando los métodos clásicos y especies comunes con perfiles bioquímicos y fenotípicos atípicos, por lo que los autores subrayan la importancia de técnicas moleculares para la identificación fúngica (21, 22).
Para Cladosporium, los resultados obtenidos por Kock (8) mostraron una conservación completa en las secuencias de los diferentes genes que codifican para el ARNr y los espaciadores internos transcriptos (ITS I y ITS II) al estudiar 44 aislamientos de C. fulvum procedentes de diferentes partes de Europa, Canadá, Estados Unidos, América del Sur, Australia, Nueva Zelanda y Japón. También Stergiopoulos et al. (13) observaron la presencia de una sola banda de aproximadamente 622bp, al amplificar la región 28S del gen que codifica para el ARNr con el empleo de estos cebadores específicos, en una colección de 81 aislamientos de C. fulvum, procedentes de diferentes regiones geográficas del mundo colectados en un período de 50 años. Nuestros resultados coinciden con los de los autores mencionados ya que todos los aislados analizados dieron, producto de la amplificación, la banda con la talla esperada, por lo que podemos afirmar que todos pertenecen a la especie C. fulvum.
Los aislamientos que fueron positivos para la amplificación con los cebadores específicos empleados para la detección de los diferentes genes Avr, rindieron un producto único en cada ensayo de PCR, cuya talla en cada caso coincidió con la informada por otros autores (Tabla 2) (13). En la Fig. 2, se muestra el resultado del análisis de las bandas amplificadas para los distintos aislamientos, referentes a los genes de avirulencia (Avrs) a partir de las cuales se realizaron agrupamientos a través de un Dendrograma.
En este caso, al realizar un corte de hasta un 70% se observa la formación de dos grupos. Este agrupamiento mostró un 97,2% de buena clasificación según el análisis de discriminante. De manera general, se encontró una alta variabilidad en la amplificación de los mismos, tanto entre grupos como dentro de un mismo grupo. En el grupo I se observó 16,6% de amplificación para el gen Avr2, mientras que el grupo II agrupó la mayoría de los aislamientos de C. fulvum analizados, con los porcentajes más altos de amplificación de estos factores de avirulencia, un 72% para Avr2 y más de un 80% para los genes de avirulencia Avr4 y Avr4E (Tabla 4).
En este mismo aspecto, se encontró correspondencia entre la amplificación de estos genes y la variabilidad patogénica observada para el hongo fitopatógeno C. fulvum, ya que los aislamientos 6, 20, 06/6, 10, 22, 7 y Quivicán, designados como raza 2, 4 y 2.4, lograron amplificar para la totalidad de los genes Avrs a diferencia del aislado 24 (determinado como raza 2), que no amplificó para Avr4 al igual que los restantes aislados (raza 0) de ese grupo, debido a lo cual posiblemente este aislado se ubique en el grupo I. El resto de los aislamientos (grupo II) determinados como raza 0, amplifican indistintamente para estos genes de avirulencia.
Aunque algunos autores plantean que los genes Avr no son requeridos para la patogenicidad de C. fulvum sobre tomate (23), recientemente, otros autores han encontrado evidencias de algunas de las posibles funciones de estos mismos genes en dicho proceso.
En una interacción incompatible, se demostró que Avr2 interactúa físicamente e inhibe a Rcr3, una Cis proteasa extracelular de tomate, similar a la papaína (que se presume es importante para la defensa basal del hospedante a C. fulvum). La unión de Avr2 a Rcr3 conlleva a un cambio conformacional de la proteasa, que es monitoreado por la proteína Cf-2 (de tomate), resultando en una reacción hipersensible (HR) y resistencia contra aislamientos de C. fulvum que producen los tipos salvajes de Avr2 (7, 24). Se ha demostrado que la función de Avr2 se requiere para la virulencia completa del patógeno y que su silenciamiento reduce la habilidad de este para causar la enfermedad (7).
También se ha hecho patente que la proteína efectora de C. fulvum Avr4 contribuye a la virulencia completa, protegiendo a la hifa del hongo quitinoso de la hidrólisis por las quitinasas de la planta, ya que contiene un dominio de unión funcional a la quitina (7, 24 25, 26).
La función de avirulencia de Avr4E no se conoce aún, pero en presencia de la proteína Cf-4E en tomate, Avr4E desata la respuesta de resistencia mediada por Cf-4E (24).
Los resultados encontrados en nuestro trabajo indican que la patogenicidad depende de un grupo de factores de virulencia que le son indispensables y que en combinación le permite al hongo ser más virulento en este hospedante, coincidiendo con lo expuesto por los autores anteriores.
Por otra parte, se encontró que ninguno de los aislamientos estudiados amplificó para el gen de avirulencia Avr9, lo que sugiere que este se encuentra ausente del genoma de los aislamientos del hongo patógeno estudiado, aún cuando algunos de ellos han resultado ser patogénicos; aspecto que coincide con resultados obtenidos por Rep (23), el cual ha encontrado cepas naturales deficientes de Avr4E y Avr9. También De Wit et al. en el 2009 (24) demostraron que el efector Avr9 contiene un nodo de cisteína con homología estructural, pero hasta el momento no funcional, con inhibidores de carboxypeptidasa y plantearon que la disrupción de Avr9 en C. fulvum no afecta su virulencia en plantas de tomate, por lo que se sugiere que Avr9 no se requiere para una virulencia completa, sino que es mas bien un factor de virulencia redundante. Esto demuestra que la proteína funcional Avr9 no es indispensable para la virulencia completa de C. fulvum, aunque al parecer, sí para vencer la resistencia de los genotipos portadores del gen Cf9.
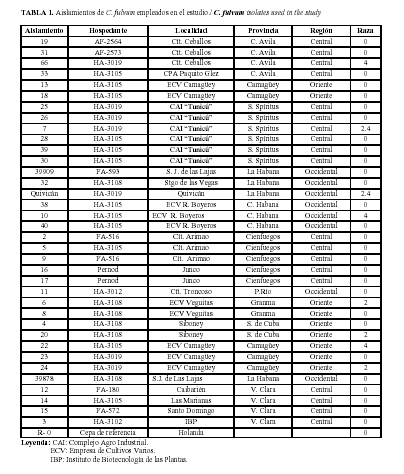
A partir de la amplificación de las proteínas extracelulares (Ecps), también se detectaron productos únicos para cada par de cebadores, con talla idéntica a la informada por otros autores en condiciones iguales a las de nuestro trabajo (Tabla 3) (13), encontrándose variabilidad entre los aislamientos analizados (Fig. 3). Al realizar un corte de hasta un 70% se observó la formación de dos grupos. Este agrupamiento mostró un 100% de buena clasificación según el análisis discriminante.
En el grupo I, representado por más del 50% de los aislamientos, es donde se observan los mayores porcentajes de amplificaciones para la mayoría de las proteínas Ecps, sobresaliendo Ecp2 con un 95,8%. En este mismo grupo el porcentaje más bajo de amplificación se encontró para la proteína extracelular Ecp1. Los aislamientos representados en el grupo I fueron los más virulentos al invadir los tejidos y esporular eficientemente sobre genotipos de tomate evaluados previamente por Bernal et al. (3), a diferencia de aquellos ubicados en el grupo II que solo amplificaron para Ecp1 (100%) y Ecp5 (15,4%) y mostraron una menor virulencia (Tabla 5).
La virulencia de las razas 2, 4, y 2.4 ubicadas en el grupo I están más relacionadas con la presencia de la proteína Ecp2 (100%) que con la Ecp1.
Todos los aislamientos determinados como raza 2, 4 y 2.4 amplificaron para todas las proteínas, excepto el aislamiento 20 (raza 2) para la Ecp1 y el 22 (raza 4) para la Ecp1 y Ecp5.
Las Ecps son proteínas secretadas abundantemente por todas las cepas de C. fulvum durante la infección, y poseen un número importante de residuos de cisteínas que probablemente estén involucrados en la formación de puentes disulfuro intramoleculares. Los Ecp6, Ecp1 y Ecp2 son genes de virulencia y se plantea que su silenciamiento o disrupción compromete la virulencia de C. fulvum en tomate (24, 27). Existen evidencias experimentales que apoyan la hipótesis acerca de que los genes Ecp 1 y Ecp 2 son importantes factores de virulencia de C. fulvum (8, 23, 28), donde la Ecp1 juega un papel principal en la invasión del hongo en el tejido y la Ecp2 en la esporulación.
Tal y como planteamos con anterioridad para los genes Avr, los resultados encontrados en nuestro trabajo para los genes Ecp confirman igualmente que la patogenicidad depende de una combinación de genes presentes en el aislamiento de forma tal que le permitan ser más virulento, aunque algunos de ellos sean determinantes y otros ejercen un efecto aditivo o menor. Al analizar los resultados de los genes Avrs (Fig. 2), como las proteínas Ecps (Fig 3) para los aislamientos determinados como raza 0, se detectan subdivisiones dentro de la raza, aspecto no informado anteriormente y que deberá ser objeto de un estudio posterior más profundo.
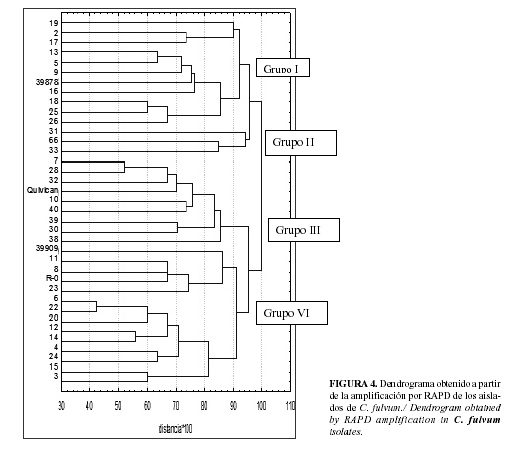
De los cebadores arbitrarios empleados para el ensayo RAPD, los cebadores OPA 02, 04, 09,10 y 13 y OPF 11 y 20 resultaron ser los mejores teniendo en cuenta el número total de bandas amplificadas (de 6-11 bandas por cebador) así como el porcentaje de bandas polimorfitas amplificadas (más del 90% para todos los casos). A través de esta técnica pudo evidenciarse una elevada variabilidad molecular entre los aislamientos analizados (Fig. 4) y en el Dendrograma correspondiente se destacan 4 grupos, tomando como punto de corte el 95% de la distancia.
En el Grupo I se ordenaron 11 aislamientos, todos de la raza 0 y el 72% de ellos originarios de la zona central del país. En el Grupo II, compuestos por tres aislamientos, estuvieron representadas las razas 0 y 4 y solamente aislamientos de la región central. El grupo III estuvo conformado por 9 aislamientos de las regiones occidental y central, en proporciones similares y representando una mayor heterogeneidad de razas (0, 2.4 y 4). Por último, el Grupo IV fue heterogéneo, tanto para el origen de los aislamientos como para las razas, con representación de todas las regiones, aunque la oriental en mayoría (54%) y de las razas 0, 2 y 4.
Los RAPD y los AFLP tienen un gran potencial para la estimación de la variabilidad genómica a través de la medición de la diversidad nucleotídica, la cual es definida como el número promedio de diferencias nucleotídicas por sitio entre dos secuencias de ADN elegidas al azar en una población. Esto se debe a que estos marcadores muestrean gran número de sitios no ligados. Ambas técnicas producen perfiles específicos individuales a partir de un ADN molde por la amplificación de fragmentos anónimos de sitios esparcidos por todo el genoma. La individualidad refleja la diversidad nucleotídica en los sitios complementarios al cebador (RAPD) o en los de restricción y sitios de extensiones del cebador (AFLP). Debido a que los RAPD y los AFLP examinan loci a lo largo del genoma y permiten obtener cantidades enormes de información con esfuerzo mínimo, los estudios con los cebadores múltiples pueden dar una estimación robusta de diversidad genética de la población, relativamente imparcial por la variación genómica local, comparadas con la estimación que pudiera tenerse a partir de los datos de las secuencias (29). Aunque, en general, es mayor la variabilidad interespecífica encontrada que la intraespecífica, y no siempre se encuentra agrupamiento de acuerdo al origen geográfico o el hospedante de procedencia, los análisis por RAPD detectan más variabilidad genética entre aislamientos que otros marcadores (30, 31, 32). Es por estas razones, que a pesar de algunos inconvenientes, continúa empleándose para estudios de variabilidad. Los hongos no escapan al uso de esta técnica alternativa.
En Cladosporium fulvum, se han utilizado para establecer un mapa genético basado en la recombinación mitótica en este hongo imperfecto. El análisis de segregación brindó evidencia molecular de un alto grado de recombinación durante el ciclo parasexual y en el nivel de ploidía de la progenie parasexual. Un estudio molecular con 49 cebadores RAPDs mostró que solo con una excepción, todos los marcadores RAPDs estudiados representaron ADN repetitivo. Esta situación predispone al uso directo de estos marcadores tanto para iniciar camino por los cromosomas hacia genes de interés o para la asignación de grupos de ligamiento a cromosomas separados electroforéticamente. Una simple reamplificación ha sido aplicada y permite la discriminación rápida entre especies de ADN de simple copia o repetitivo, sin necesidad de análisis de hibridación. Además se encontró evidencia para la presencia de especies de ADN de simple hebra en los productos de amplificación y que estas pueden estar presentes además de sus contrapartes de doble cadena. La reamplificación puede también identificar estas especies, evitando así los errores de interpretación de bandas polimórficas (33).
Aun cuando no se encontró una completa correspondencia del agrupamiento obtenido por los datos moleculares del RAPD con la región de procedencia, con la raza de cada aislamiento o con la presencia de genes de Avr y Ecp, en el presente trabajo si fue posible demostrar la variabilidad entre los aislados estudiados. Los resultados obtenidos en este trabajo apoyan el programa de mejoramiento del tomate para la obtención de resistencia a C. fulvum y su distribución en el país, teniendo en cuenta que ofrecen una idea de la diversidad genética del patógeno, teniendo en cuenta diferentes técnicas y especialmente las relacionadas con la detección específica de algunos factores de patogenicidad.
REFERENCIAS
1. Thomma BPHJ, van Esse HP, Crous PW, De Wit PJGM. Cladosporium fulvum (syn. Passalora fulva) a highly specialized plant pathogen as a model for functional studies on plant pathogenic Mycosphaerellaceae. Molecular Plant Pathology. 2005;43:452-485.
2. Thomma BHJ, Van Esse HP, Crous PW, De Wit PJGM. Cladosporium fulvum (syn. Passalora fulva), a highly specialized plant pathogen as a model for functional studies on plant pathogenic Mycosphaerellaceae. Mol Plant Pathol. 2008;6(4):379-393.
3. Bernal A, Martínez B, González E. Incidencia de Cladosporium fulvum Cooke. en híbridos de tomate bajo condiciones de cultivo protegido. Centro Agrícola. 2005;32(2):31-35.
4. Veloukas T, Bardasa GA, Karaoglanidisb GS, Tzavella-Klonaria K. Management of tomato leaf mould caused by Cladosporium fulvum with trifloxystrobin. Crop Protection. 2007;26:845-851.
5. Westerink N, Brandwagt BF, De Wit PJG, Joosten MHAJ. Cladosporium fulvum circumvents the second functional resistance gene homologue at the Cf-4 locus (Hcr9-4E) by secretion of a stable avr4E isoform. Mol. Microbiol. 2004;54:533-545.
6. Bolton MD, Van Esse HP, Vossen JH, De Jonge R, Stergiopoulos I, Stulemeijer IJE, et al. The novel Cladosporium fulvum lysin motif effector Ecp6 is a virulence factor with orthologues in other fungal species. Mol. Microbiol. 2008; 69:119-136.
7. Van Esse HP, Van Klooster JW, Bolton MD, Yadeta KA, van Baarlen P, Boeren S, et al. The Cladosporium fulvum Virulence Protein Avr2 Inhibits Host Proteases Required for Basal Defense. The Plant Cell. 2008;20:1948-1963.
8. De Kock MJD, Iskander HM, Brandwagt BF, Lauge R, De Wit PJGM, Lindhout P. Recognition of Cladosporium fulvum Ecp2 elicitor by non-host Nicotiana spp. Is mediated by a single dominant gene that is not homologous to known Cf genes. Mol Plant Pathol. 2004;5:397-408.
9. Gamborg OL, Millar RA, Ojiva K. Nutrients requirements of suspensión cultura of soybean root cells. Exp Cell Res. 1968;50:148-151.
10.Spanu P. HCf-1 a hydrophobin gene of Cladosporium fulvum, does not affect pathogenecity in tomato. Physiol Mol Plant Pathol. 1998;52:323-334.
11.Peteira B, Perez S, Fraga Y. A modified DNA extraction minipreparation protocol for RAPDs analysis of genetic variability in Alternaria solani. Rev. Prot Veg. 1999;(2):121-124.
12.Sambrook J, Fritsch EF, Maniatis T. Molecular Cloning. 1989. A laboratory Manual, Second Edition. Cold Spring Harbor laboratory Press.
13.Stergiopoulos I, Groenewald M, Staats M, Lindhout P, Crous PW, De Wit PJGM. Mating-type genes and the genetic structure of a world-wide collection of the tomato pathogen Cladosporium fulvum. Fungal Genetics and Biology. 2007;44:415-429.
14.Nei M, Li WH. Mathematical model for studding genetic variation in terms of restriction endonucleases. Proc. Nat. Acad. Sci. USA. 1979;76:5267-5273.
15.SPSS v. 11.5 para Windows (SPSS Inc., 2000).
16.Johanson A, Turner HC, McKay GJ, Brown AE. A PCR-based method to distinguish fungi of the rice sheath-blight complex, Rhizoctonia solani, R. oryzae and R. oryzae-sativae. FEMS Microbiology Letters. 1998;162:289-294.
17.Bilodeau GJ, Lévesque CA, De Cock AWAM, Duchaine C, Brière S, Uribe P, et al. Molecular Detection of Phytophthora ramorum by Real-Time Polymerase Chain Reaction Using TaqMan, SYBR Green, and Molecular Beacons. Phytopathol. 2007;97(5):632-642.
18.Alymanesh MR, Falahatirastegar M, Jafarpour B, Mahdikhanimoghadam E. Genetic diversity in the fungus Fusarium solani f.sp. cucurbitae race 1, the casual agent of root and crown rot of cucurbits in Iran, using molecular markers. Pak. J. Biol. Sci. 2009;12:836-843.
19.Becerra VV, Paredes CM, Rojo MC, France IA. RAPD and ITS Reveal Molecular Variation of Chilean Populations of Beauveria bassiana. Agricultura Técnica (Chile). 2007;67(2):115-125.
20.Gaskell GJ, Carter DA, Britton WJ, Tovey R, Benyon FHL, Lovborg U. Analysis of the internal transcribed spacer regions of ribosomal DNA in common airborne allergenic fungi. Electroforesis Published Online: 14 Apr 2005;18(9):1567-1569.
21.Borman AM, Linton CJ, Miles SJ, Johnson EM. Molecular identification of pathogenic fungi. Journal of Antimicrobial Chemotherapy 2008;61(Supplement 1):i7-i12; doi:10.1093/jac/dkm425.
22.Soeta N, Terashima M, Gotoh M, Mori S, Nishiyama K, Ishioka K, et al. An improved rapid quantitative detection and identification method for a wide range of fungi. J Med Microbiol. 2009;58:1037-1044.
23.Rep M. Small proteins of plant-pathogenic fungi secreted during host colonization. FEMS Microbiol Letters. 2005;253:19-27.
24.De Wit PJGM, Mehrabi R, Van der Burg H and Stergiopoulos I. Fungal effector proteins: past, present and future. Mol. Plant Pathol. 2009;10(6):735-747.
25.Van den Burg HA, Harrison SJ, Joosten MHAJ, Vervoort J, De Wit PJGM. Cladosporium fulvum Avr4 protects fungal cell walls against hydrolysis by plant chitinases accumulating during infection. Mol. Plant Microbe Interact. 2006;19:1420-1430.
26.Van Esse HP, Bolton MD, Stergiopoulos I, De Wit PJGM, Thomma BPHJ. The chitin-binding Cladosporium fulvum effector protein Avr4 is a virulence factor. Mol. PlantMicrobe Interact. 2007;20:1092-1101.
27.Thomma BPHJ, Bolton MD, Clergeot PH, De Wit PJGM. Nitrogen controls in planta expression of Cladosporium fulvum Avr9 but no other effector genes. Mol. Plant Pathol. 2006;7:125-130.
28.Kruijit M. Molecular evolution of Cladosporium fulvum disease resistance genes in wild tomato. Thesis Wageningen University, The Netherlands. 2004; 136pgs.
29.Borowsky RL. Estimating Nucleotide Diversity From Random Amplified Polymorphic DNA and Amplified Fragment Length Polymorphism Data. Molecular Phylogenetics and Evolution. 2001;18(1):143-148.
30.Baires-Varguez L, Cruz-García A, Villa-Tanaka L, Sánchez-García S, Gaitán-Cepeda LA, Sánchez-Vargas LO, et al. Comparison of a randomly amplified polymorphic DNA (RAPD) analysis and ATB ID 32C system for identification of clinical isolates of different Candida species. Rev Iberoam Micol. 2007;24:148-151.
31.Jun-Zhi Q, Zhi-Peng H, Jie-Ru P, Xue-Qin X, Yan-Ping Z, Fang F, et al. RAPD and large subunit nuclear rDNA sequence analyses of the entomogenous fungus Aschersonia. Chinese J. Agricul Biotechnol. 2005;2:85-90.
32.Mitina GV, Mikhailova LA, Mattila TY. RAPD-PCR, UP-PCR and rDNA sequence analyses of the entomopathogenic fungus Verticillium lecanii and its pathogenicity towards insects and phytopathogenic fungi. Arch of Phytopathol and Plant Prot. 2008;41(2):113-128.
33.Arnau J, Housego AP, Oliver RP. The use of RAPD markers in the genetic analysis of the plant pathogenic fungus Cladosporium fulvum. Curr Genet. 1994;25(5):438-44.
(Recibido 28-5-2010; Aceptado 10-6-2010)


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}