










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión impresa ISSN 1010-2752
Rev. Protección Veg. vol.27 no.3 La Habana sep.-dic. 2012
ARTÍCULO ORIGINAL
Biología y conducta alimentaria de Neoseiulus longispinosus (Evans) sobre Tetranychus tumidus Banks
Biology and feeding behavior of Neoseiulus longispinosus (Evans) on Tetranychus tumidus Banks
Yanebis Pérez MadrugaI, D. Alonso-RodríguezII, R. ChicoI, H. RodríguezII
IGrupo Plagas Agrícolas. Dirección de Protección de Plantas. Centro Nacional de Sanidad Agropecuaria (CENSA). Carretera de Jamaica y Autopista Nacional. Apdo 10, San José de las Lajas. Mayabeque. Cuba. Correo electrónico: yanebis@censa.edu.cu.
IIFacultad de Agronomía. Universidad Agraria de La Habana (UNAH). Carretera de Tapaste y Autopista Nacional. San José de las Lajas, Mayabeque. Cuba. CP 32 700.
RESUMEN
Se determinó la duración del desarrollo, mortalidad, cociente sexual, reproducción, sobrevivencia y la conducta alimentaria de Neoseiulus longispinosus (Evans) cuando se alimentó de Tetranychus tumidus Banks. Los estudios se realizaron en condiciones de laboratorio a 24,34±2,90°C de temperatura y 73,54±11,04% de humedad relativa. Se encontró que la duración media del desarrollo fue aproximadamente de siete días. La fase de huevo tuvo la mayor duración, mientras que la de larva fue la menor. El período de preoviposición tuvo una duración media de 2,3 días, mientras que el período de oviposición puede durar hasta 22 días, con un valor medio de 11,5 días y una longevidad de las hembras de hasta 28 días. La fecundidad media fue 21,70±9,11 huevos. El cociente sexual fue de 0,70 favorable a las hembras. No se encontró mortalidad en las fases inmaduras. El número de huevos/hembra/día se incrementó desde el segundo día de oviposición hasta el octavo día. N. longispinosus consumió preferentemente la fase de huevo, aunque la mayor tasa de éxito la alcanzó sobre las fase de larva y deutoninfa. El depredador utilizó más del 50% de su tiempo en los componentes activos de búsqueda y consumo de presas. Los resultados sugieren que este ácaro fitoseido es un biorregulador potencial para T. tumidus en nuestras condiciones.
Palabras clave: Neoseiulus longispinosus, Tetranychus tumidus, duración del desarrollo, conducta alimentaria, ácaro depredador, Phytoseiidae.
ABSTRACT
Development duration, mortality, sex ratio, reproduction, survival and feeding behavior of Neoseiulus longispinosus (Evans) feeding on Tetranychus tumidus Banks were determined. The studies were conducted under laboratory conditions at 24,34±2,90°C temperature and 73,54±11,04% relative humidity. The average duration of development was observed to be approximately seven days. The egg stage was the longest, whereas the larval stage was the shortest. The pre-oviposition period lasted a mean of 2,3 days while the oviposition period can last up to 22 days with an average of 11,5 days and female longevity up to 28 days. The average fertility was 21,70±9,11 eggs. The sex ratio was 0,70 in favor of females. No mortality was found in immature stages. The number of eggs per female/day increased from the second to the eighth day of oviposition. N. longispinosus preferentially consumed eggs, whereas it reached the highest rate of success on the larval and immature stages. The predator spent more than 50% of its time on the active components of consuming and searching. The results suggest that this phytoseiid mite is a potential bioregulator of T. tumidus in our conditions.
Key words: Neoseiulus longispinosus, Tetranychus tumidus, duration of development, feeding behavior, predatory mites, Phytoseiidae.
INTRODUCCIÓN
El ácaro, Tetranychus tumidus Banks (Acari: Tetranychidae), conocido como araña roja, es uno de los tetraníquidos más difundidos en el mundo sobre el follaje del plátano y el banano (Musa spp.) (1). Es una especie polífaga, que en Cuba se informó en más de 50 especies de plantas, entre ellas: Musa spp., Phaseolus vulgaris L., Gossypium sp., Zinnia elegans L., Phytolacea sp., Ipomoea batata (L.) Sam, Albizia lebbek (L.) Benth y Calotropis procera (Oit.) R.Br, por solo citar algunas. Posee un ciclo de desarrollo corto, alta adaptabilidad y plasticidad ecológica y resulta fácil su reproducción en condiciones de laboratorio (2).
En Cuba, en el cultivo del plátano y el banano, T. tumidus produce las mayores afectaciones en condiciones de vivero. En este sistema, las primeras poblaciones aparecen a partir de los 15 ó 20 días después de plantadas las vitroplantas. El ácaro se localiza preferentemente en el envés de las hojas más viejas, de las cuales migra hacia las hojas más jóvenes. En estas condiciones, puede llegar a impedir la comercialización de las posturas. También puede convertirse en el inóculo que llega a las plantaciones de fomento y por lo tanto, producir daños económicos, debido a la poca incidencia de enemigos naturales en esta fase del cultivo (3,4,5).
A partir de estudios realizados en el país, se establecieron las potencialidades de los ácaros depredadores de la familia Phytoseiidae para el manejo de la araña roja en la fase de vivero. Se determinó que con la liberación de Phytoseiulus macropilis (Banks) (Acari: Phytoseiidae), en una relación presa depredador (20:1) se lograba una regulación eficiente de las poblaciones de T. tumidus (3,6). No obstante, esta especie, como depredador especialista, tiende a abandonar las plantas cuando suprime la población de la presa. Esta conducta pudiera generar la necesidad de realizar nuevas liberaciones del biorregulador.
Este inconveniente se ha resuelto en otros agroecosistemas con la liberación combinada de fitoseidos con diferentes preferencias alimentarias (7). Entre las opciones posibles está utilizar un depredador facultativo (8), como Neoseiulus longispinosus (Evans) (Acari: Phytoseiidae).
N. longispinosus es uno de los fitoseidos más abundante y efectivo en Tailandia y comúnmente se encuentra asociado a diversas especies de ácaros fitófagos en cultivos de interés económico (7). Se conoce que su rol en la supresión de las poblaciones del ácaro de dos manchas, Tetranychus urticae Koch (Acari: Tetranychidae) es evidente en fresal en el norte de Tailandia. Por estas potencialidades, se realizaron estudios para garantizar su utilización eficaz en programas de manejo de plagas (10). Peña et al. (11) lo encontraron depredando a Raoiella indica Hirst (Acari: Tenuipalpidae) en Santa Lucía. También fue estudiado en China para el control de Aponychus corpuzae Rimando y Schizotetranychus nanjingensis Ma et Yuan (12,13).
Recientemente se informó la presencia de N. longispinosus sobre hojas de Cocos nucifera L. y Areca catechu L. en asociación con ácaros tetraníquidos en Cuba (14). Posteriormente se detectó contaminando crías de T. tumidus en condiciones de aisladores biológicos del Centro Nacional de Sanidad Agropecuaria (CENSA) (Observación personal). Este hallazgo sugiere que se pudiera valorar la liberación conjunta de un depredador especialista y uno facultativo para manejar las poblaciones de la araña roja y con ello lograr un efecto de control más duradero. Por ello, el objetivo del presente trabajo fue determinar los parámetros biológicos fundamentales y la conducta alimentaria de N. longispinosus sobre T. tumidus, como elementos básicos para establecer sus potencialidades biorreguladoras.
MATERIALES Y MÉTODOS
Cría de T. tumidus y N. longispinosus: Los individuos utilizados en los ensayos provenían de colonias mantenidas en el Laboratorio de Acarología del CENSA (Mayabeque). T. tumidus se crió sobre plantas de frijol obtenidas en macetas en el laboratorio, mientras que N. longispinosus se reprodujo en bandejas de acero inoxidablede de 30x20x5 cm (largo x ancho x altura) utilizando hojas de frijol infestadas con T. tumidus como presa.
Desarrollo, fecundidad y longevidad de N. longispinosus
Se extrajeron 20 hembras de N. longispinosus de la cría establecida y se depositaron en foliolos de frijol con abundante población de T. tumidus, colocados en placas Petri de 11cm de diámetro con algodón humedecido. Estos foliolos se observaron cada 2 horas para extraer los huevos ovipositados por las hembras, con un pincel 00. Los huevos recolectados, se transfirieron de forma individual a un foliolo de frijol infestado con T. tumidus, situado en pequeñas cajas plásticas con cierre hermético.
Los huevos obtenidos se observaron cada 24 horas y se anotaron los cambios de fase y la duración de las mismas hasta la emergencia de los adultos. Con estos datos se calculó la duración media del desarrollo, la desviación estándar y los valores máximos y mínimos absolutos por fase, en días, para ambos sexos y total. Se estimó el cociente sexual a través de la expresión: hembra/(hembra+macho) y se determinó el número de individuos muertos naturalmente para cada fase y total.
Las hembras obtenidas se mantuvieron individualizadas en las unidades experimentales y se les colocó un macho de la cría durante 24 horas para garantizar la primera cópula. Una vez que se confirmó el apareamiento, se observaron diariamente hasta la muerte de las hembras y se registró el número de huevos ovipositados.
Con los datos obtenidos se determinó la duración promedio, la desviación estándar y los valores máximos y mínimos absolutos de los períodos de preoviposición, oviposición, postoviposición y la longevidad de las hembras. Además se determinó la curva de sobrevivencia (Lx) (% de hembras vivas/día), la fecundidad media y la tasa de oviposición diaria (media de huevos/hembra/día).
A partir de una cohorte de 61 huevos, se determinó la fertilidad. Los huevos se observaron diariamente, considerándose no viables aquellos que no eclosionaron en un término de siete días.
Conducta alimentaria
Se extrajeron hembras grávidas de N. longispinosus de la cría establecida en laboratorio y se colocaron en ayuno durante 24 horas, en pequeñas unidades experimentales con cierre hermético. Transcurrido ese período, las hembras fueron colocadas sobre foliolos de frijol infestados con todas las fases de T. tumidus en placas Petri de 11cm de diámetro.
A partir de ese momento, la unidad experimental se colocó bajo un microscopio estereoscópico Zeiss Stemi SV 6 y se observaron todas las actividades que realizó la hembra durante 30 minutos. Se empleó la menor intensidad luminosa que permitiera observar la hembra con nitidez, de manera tal que no se perturbara su comportamiento. Durante la realización del experimento se observaron y midieron las siguientes actividades:
• Número de presas contactadas
• Número de presas consumidas
• Tiempo de manipulación y consumo
• Tiempo de descanso
• Tiempo de limpieza
• Tiempo en actividades de búsqueda
• Tiempo empleado en tomar agua.
Para realizar estas observaciones se siguieron los criterios de Rodríguez y Ramos (15). Este experimento se replicó 14 veces. Con estos datos se determinó el número promedio de presas contactadas y consumidas por la hembra de N. longispinosus para cada fase de la presa y total, así como la tasa de éxito en el ataque, considerando las presas contactadas y las consumidas.
Se calculó, además, la duración promedio (en segundos) de la alimentación de N. longispinosus sobre cada fase de la presa y el promedio general considerando todas las fases, así como la proporción de tiempo que dedica el depredador a cada una de las actividades conductuales descritas anteriormente.
Los experimentos se realizaron en condiciones de laboratorio, con una temperatura de 24,34±2,90°C, 73,54±11,04% de humedad relativa y fotoperíodo natural.
RESULTADOS Y DISCUSIÓN
Desarrollo, fecundidad y longevidad de N. longispinosus
El ácaro N. longispinosus tiene cuatro fases de desarrollo: el huevo, la larva con tres pares de patas, y dos estadios con cuatro pares de patas, la protoninfa y la deutoninfa. A partir del huevo el cambio de fase estuvo determinado por la presencia de la exuvia.
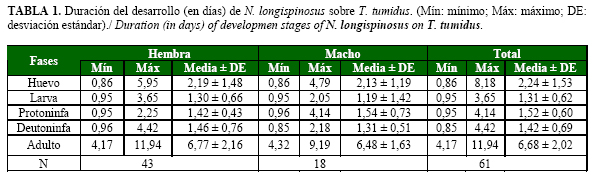
La Tabla 1 muestra la duración por fase y total para ambos sexos. N. longispinosus necesitó poco tiempo para alcanzar la fase de adulto, aproximadamente siete días. La fase de huevo tuvo la mayor duración, representando el 33,53% de la duración total; mientras que la larva fue la menor. De los dos estadios ninfales, la protoninfa demoró más. Este resultado coincide con el informado para esta especie por Ganok (16), quien registró una duración del desarrollo sobre Tetranychus truncatus Ehara de aproximadamente 7 días (T= 30-32°C; HR= 65-70%). Sin embargo, es superior al informado por Lababidi (17), quien notificó una duración de 5 y 4,4 días para las hembras y los machos, respectivamente. Cuando la duración del desarrollo se estudió a temperatura constante (25°C), se encontró que la fase de mayor duración fue la de huevo con una media de 2,2 días, similar a lo observado en este estudio; mientras que la duración fue de 0,6; 1,1 y 0,7 días, para las fases de larva, protoninfa y la deutoninfa, respectivamente, al alimentarse de Tetranychus cinnabarinus (Boisd.).
A partir de la fase de larva no se produjo mortalidad, lo cual evidenció la elevada posibilidad de sobrevivencia de las fases inmaduras de N. longispinosus. Un resultado semejante encontraron Rodríguez y Ramos (15) para Amblyseius largoensis (Muma) a una temperatura de 20,86ºC. La relación sexual fue de 0,70, similar a la informada para esta especie (17).
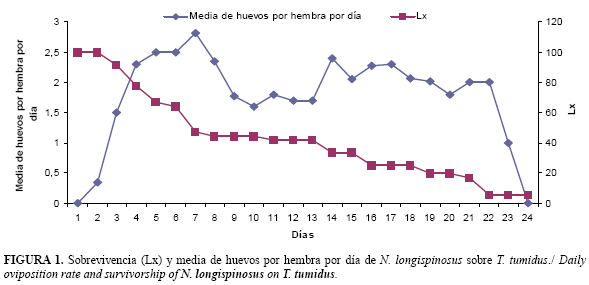
La duración en días de los períodos de preoviposición, oviposición y postoviposición de N. longispinosus se muestran en la Tabla 2. El período de preoviposición fue mayor, mientras que el período de oviposición fue inferior al informado por Rodríguez y Ramos (15) para A. largoensis en laboratorio. Según Lababidi (17), la longevidad de N. longispinosus varió en función de la temperatura, siendo de 33,1; 30,6 y 29,9 días a 25, 30 y 33°C, respectivamente cuando se alimentó de T. cinnabarinus.
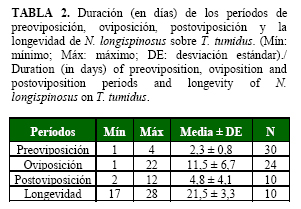
La oviposición comienza, aproximadamente, a los dos días de la emergencia de las hembras. El número de huevos puestos comienza a incrementarse desde el tercer día de la oviposición y se mantiene elevado durante los ocho primeros días, momento a partir del cual decrece gradualmente y la hembra deja de ovipositar a los 24 días después del inicio de la puesta. El pico máximo de puesta se produce entre los tres y siete días (Fig.1). Los huevos alcanzaron un 100% de eclosión.
En este estudio se encontró que aproximadamente mueren un número constante de hembras por unidad de tiempo, independientemente del número de individuos que han sobrevivido (Fig. 1). Esto se corresponde con una curva intermedia entre el tipo I y II, según señaló Vera (18). La sobrevivencia comenzó a disminuir a partir del día tres y lo hizo de forma drástica al principio y después de forma paulatina.
La fecundidad promedio fue de 21,70±9,11 huevos, con un mínimo de 3 y un máximo de 37 huevos, valores inferiores a los encontrados por Lababidi (17), quien informó que las hembras de N. longispinosus produjeron como promedio 48,8 huevos durante 21 días de oviposición. Esto puede deberse a que en este estudio se le colocó solo un macho, lo que es común en estudios con ácaros (19). Sin embargo, se conoce que varios machos por unidad experimental garantizan la posibilidad de múltiples cópulas, lo cual se considera imprescindible para alcanzar la máxima oviposición en muchas especies de ácaros, aspecto que debe ser considerado en futuros ensayos con este depredador.
Los valores del ciclo de desarrollo y los parámetros reproductivos de N. longispinosus sobre T. tumidus obtenidos en este estudio, son en la mayoría de los casos, similares a los informados para otros fitoseidos, que se reconocen como buenos biorreguladores de ácaros tetraníquidos, lo cual avala favorablemente la potencialidad que presenta para controlar a la araña roja.
Conducta alimentaria
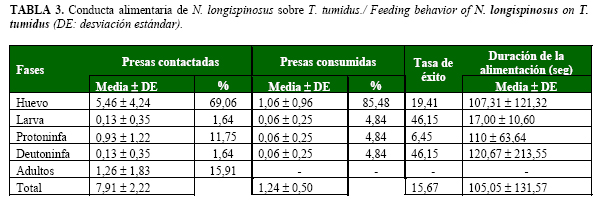
En 30 minutos, N. longispinosus contactó todas las fases de T. tumidus. El mayor número de encuentros se registró para la fase de huevo y adulto y los menores fueron para larvas y deutoninfas. Sin embargo, consumió preferentemente huevos, los cuales representaron aproximadamente el 85% de todas las presas ingeridas. No obstante, la tasa de éxito fue más elevada sobre larvas y deutoninfas, seguida de los huevos. Esto pudo deberse, al reducido número de individuos contactados y consumidos, para estas fases. El depredador no consumió ningún adulto (Tabla 3).
Estos resultados coinciden con lo planteado por Mallik y Rajashekar (20) para esta especie frente a T. urticae, quienes demostraron que este depredador prefiere los huevos y los estadios inmaduros de su presa por encima de los adultos.
Este depredador tiene una adecuada capacidad de búsqueda y una elevada agresividad frente a los huevos y fases inmaduras, lo que influye de forma significativa en la tasa de consumo de la presa.
Los contactos con los huevos y las fases inmaduras condujeron a una mayor proporción de ataques exitosos debido, fundamentalmente, a que son más fáciles de consumir. Los adultos resultan presas más difíciles, ya que tienen el tegumento más esclerosado y una mayor movilidad, en comparación con las restantes fases.
El tiempo que demora la hembra de N. longispinosus en consumir las diferentes fases de la presa tuvo un valor promedio cercano a los dos minutos. Se observó una tendencia al aumento de la duración de la alimentación con el incremento del tamaño de la presa, con excepción de la fase de larva, que presentó la menor duración (Tabla 3). Esto sugiere que el depredador durante la alimentación pudo haber sido perturbado por la presencia de otra presa potencial.
El tiempo que demora un depredador alimentándose de una presa individual depende, entre otros factores, del tamaño de la presa, el nivel de apetito y la excitabilidad del depredador (21).
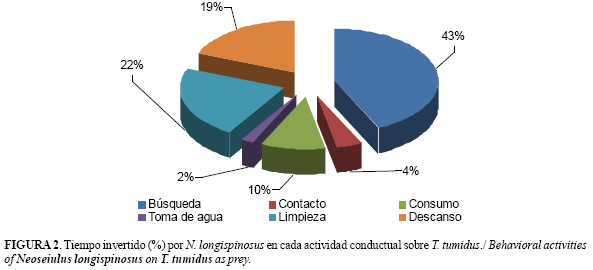
El porcentaje de tiempo invertido en las diferentes actividades conductuales del biorregulador se muestra en la figura 2. N. longispinosus ocupó la mayor parte del tiempo en actividades de búsqueda y limpieza, seguido por las etapas de descanso y consumo. Rodríguez y Ramos (15) en un estudio similar para A. largoensis informaron que la mayor parte del tiempo la dedicó a componentes activos (búsqueda y consumo de presas), pero con un mayor porcentaje en la actividad de búsqueda. Sin embargo, el resultado encontrado en este estudio fue similar al notificado por Rodríguez et al. (22) para A. largoensis frente a R. indica.
Al respecto, Sandness y McMurtry (21) indicaron que, en ocasiones, el tiempo de búsqueda no está directamente relacionado con actividades de alimentación, pues los fitoseidos realizan también esfuerzos para localizar lugares adecuados para la oviposición. Estos autores encontraron que A. largoensis tuvo un tiempo de búsqueda menor y mayores porcentajes del tiempo dedicado al consumo, la limpieza y al descanso, frente a Oligonychus punicae (Hirst).
Estos resultados en su conjunto ponen de manifiesto la marcada habilidad que presentó. N. longispinosus para encontrar, capturar y consumir las fases inmaduras de su presa. Estos elementos son útiles para definir la tasa óptima de liberación y para el diseño de la estrategia futura de control de T. tumidus con este biorregulador.
REFERENCIAS
1. Pérez R, Almaguel L, Cáceres I, Feitó E, de la Torre E. Los depredadores de Tetranychus tumidus Banks en Cuba. 2004. (En línea). Disponible en: http://agris.fao.org/?query=%2BcitationTitle Consultada: 9 de febrero de 2011.
2. Almaguel L. CIDISAV-INISAV. Curso Introductorio a la Acarología Aplicada. Morfología, taxonomía y diagnóstico fitosanitario de ácaros de importancia agrícola. La Habana. 2004.
3. Gil M. Estacionalidad de Tetranychus tumidus Banks (Acari: Tetranychidae) y su control mediante Phytoseiulus macropilis (Banks) (Acari: Phytoseiidae) en viveros de platano. Tesis presentada en opción al título de Ingeniero Agrónomo. Universidad Agraria de la Habana (UNAH), Cuba. 1995.
4. Ramos M. Uso de Phytoseiulus macropilis (Acari: Phytoseiidae) en el control de Tetranychus tumidus (Acari: Tetranychidae) en viveros de plátano. Tesis presentada en opción al grado científico de Doctor en Ciencias Agrícolas. Universidad Agraria de la Habana (UNAH), Cuba. 1995.
5. Almaguel L. INISAV (CID-INISAV Boletín Técnico, 2). Ácaros de importancia económica en Cuba. Ciudad de La Habana: 1996.
6. Ramos M. Control de Tetranychus tumidus mediante Phytoseiulus macropilis en viveros de plátano. Manejo Integrado de Plagas. 2000;58:54-60.
7. Grafton-Cardwell EE, Ouyang Y, Striggow RA. Predaceous mites (Acari: Phytoseiidae) for control of spider mites (Acari: Tetranychidae) in nursery citrus. Biological Control. 1997;14:29-36.
8. McMurtry JA, Croft BA. Life-Styles of phytoseiid mites and their roles in biological control. Ann Rev Entomol. 1997;42:291-321.
9. Kongchuensin M, Charanasri V, Takafuji A. Geographic Distribution of Neoseiulus longispinosus (Evans) and Its Habitat Plants in Thailand. J Acarol Soc Jpn. 2005;14(1):1-11.
10.Kongchuensin M, Charanasri V, Kulpiyawat T, Khantonthong K. Biological control of two-spotted spider mite in strawberries by the predatory mite, Amblyseius longispinosus (Evans) (Acari: Phytoseiidae). En: Halliday RB, Walter DE, Proctor HC, Norton RA, Colloff MJ, eds. Acarology: Proceedings of the 10th International Congress. pp. 513-517, CSIRO Publishing, Melbourne, 2001.
11.Peña JE, Mannion CM, Howard F, Hoy MA. University of Florida IFAS Extension, Raoiella indica (Prostigmata: Tenuipapidae): The Red Palm Mite, A potential invasive pest of palms and bananas and other tropical crops of Florida, 2006. ENY- 837 (En línea). Disponible en: http://edis.ifas.ufl.edu/BODY_IN681. Consultada: 25 jun 2010.
12.Zhang YX, Zhang ZQ, Lin JZ, Liu QY. Predation of Amblyseius longispinosus (Acari: Phytoseiidae) on Aponychus corpuzae (Acari: Tetranychidae). Syst Appl Acarol. 1998;3:53-58.
13.Zhang YX, Zhang ZQ, Ji J, Lin JZ. Predation of Amblyseius longispinosus (Acari: Phytoseiidae) on Schizotetranychus nanjingensis (Acari: Tetranychidae). Syst Appl Acarol. 1998;4:63-68.
14.Hastie E, Benegas A, Rodríguez H. Inventario de ácaros depredadores asociados a fitoácaros en plantas de las familias Arecaceae y Musaceae. Rev Protección Veg. 2010;25(1):17-25.
15.Rodríguez H, Ramos M. Biology and feeding behavior of Amblyseius largoensis (Muma) (Acari: Phytoseiidae) on Polyphagotarsonemus latus (Banks) (Acari: Tarsonemidae). Rev Protección Veg. 2004;19(2):73-79.
16.Ganok U. Preliminary study on the biology of cassava red mite Tetranychus truncatus Ehara (Acarina: Tetranychidae) and the predaceous mite Amblyseius (Amblyseius) longispinosus (Evans) (Acarina: Phytoseiidae). Tesis en opción el grado académico de Masters Sciences. Kasetsart University. Bangkok, Tailandia. 1982.
17.Lababidi MS. Development, longevity, fecundity and sex ratio of the predatory mite Amblyseius longispinosus (Evans) with Tetranychus cinnabarinus (Boisd.) as prey. Anzeiger fuer Schaedlingskunde Pflanzenschutz Umweltschutz. 1989;34:23-28.
18.Vera G. Técnicas demográficas. En: Vera J, ed. Tema III: Temas selectos sobre ecología de poblaciones. 61-99, 1990.
19.Schulten GGM. Mating. En: Helle W, Sabelis MW. Eds. Spider mites: their biology natural enemies and control. (World Crop Pests). 55-66, 1985.
20.Mallik B, Rajashekar K. Amblyseius longispinosus (Acari: Phytoseiidae) an efficient under-exploited predator of the two spotted spider mite in the tropics. En: XI International Congress of Acarology. Program and Abstract Book. Mérida, Yucatán, Mexico. 2002.
21.Sandness JN, McMurtry JA. Prey consumption behaviour of Amblyseius largoensis in relation to hunger. Can Entomol. 1972;104:61-70.
22.Rodríguez H, Montoya A, Flores-Galano G. Conducta alimentaria de Amblyseius largoensis (Muma) sobre Raoiella indica Hirst. Rev Protección Veg. 2010;25(1):26-30.
Recibido: 29-9-2011.
Aceptado: 30-9-2012.

{kind=link}
{kind=link}
{kind=link}
{kind=link}