










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Las infecciones por Staphylococcus aureus resistentes a la meticilina (SARM) constituyen un problema de salud que incrementa los costos en los servicios hospitalarios, debido al aumento de la estadía del enfermo en los hospitales, así como, de las tasas de morbilidad y mortalidad. (1,2,3
El mecanismo de resistencia a los betalactámicos en SARM está condicionado por la existencia del gen cromosómico mecA, que codifica la síntesis de una proteína fijadora de la penicilina (PBP2a), ausente en los cultivos de estafilococos sensibles a la meticilina; además, este gen es capaz de sustituir en sus funciones a otras PBP y tiene poca afinidad por todos los betalactámicos. (4
La preocupación atribuida a SARM no solo radica en su resistencia intrínseca a los betalactámicos, sino a otros grupos de antimicrobianos y a su fácil transmisión, que determina el desarrollo obligatorio de protocolos para garantizar su control. (4,5,6
Dentro de los principales factores de virulencia que contribuyen a la resistencia de S. aureus se incluyen: las toxinas exfoliativas A y B, la leucocidina de Panton Valentine (PVL, por sus siglas en inglés de Panton Valentine Leukocidin), hemolisinas (α, β, γ, δ), el grupo de las toxinas pirógenas superantígenos (PTSAg), que incluye la toxina del choque tóxico, las enterotoxinas estafilocócicas (SE) y las modulinas solubles en fenol, descritas reciente, el factor de aglutinación, las proteínas de unión al fibrinógeno, la fibronectina, la sialoproteína ósea, así como las citotoxinas y proteínas de adhesión extracelular, que le permiten evadir los mecanismos de defensa del hospedero. 7,8
Desde el punto de vista epidemiológico, en regiones de América detectan cifras variables de aislamientos de SARM, las cuales oscilan entre el 45,5 % en pacientes hospitalizados y el 17,7 % en pacientes de la comunidad. El tratamiento de elección para las infecciones causadas por este microorganismo es la vancomicina; sin embargo, en la actualidad, se aíslan cepas de SARM resistentes a este antimicrobiano. (9
Hoy en día esta situación es común en las instituciones hospitalarias. La resistencia de S. aureus a la meticilina es sinónimo de resistencia frente a todos los betalactámicos que incluyen a otras penicilinas, a las cefalosporinas y a los carbapenémicos, además, se asocia a la resistencia múltiple frente a los antibacterianos no relacionados con sus estructuras como las tetraciclinas, los macrólidos, las quinolonas y los aminoglucósidos. (10,11,12,13
Por lo antes expuesto el desarrollo y la obtención de moléculas con actividad antimicrobiana demostrada frente a cepas de SARM, constituyen una prioridad para el sistema de salud cubano.
La peroxiadenosina es una molécula de origen semisintético desarrollada en el Centro de Inmunología y Productos Biológicos (CENIPBI) de Camagüey, obtenida por la peroxidación de nucleósidos de adenina con el peróxido de hidrógeno. Tiene un carácter hidrofílico por la presencia de grupos polares en su azúcar y en la base nitrogenada. Se comporta como una base débil y es estable en soluciones acuosas.
Por desconocerse con anterioridad, la utilidad de la peroxiadenosina in vitro frente a bacterias con estas características, el estudio constituye la primera investigación dirigida a evaluar la actividad antimicrobiana in vitro de moléculas de adenosina modificadas con peróxido de hidrógeno frente a SARM en la provincia Camagüey.
Métodos
Se realizó un estudio experimental preclínico, in vitro, en un modelo biológico (cultivo microbiano), desde enero de 2016 hasta julio de 2020, con el objetivo de evaluar la actividad antimicrobiana de la peroxiadenosina en una cepa salvaje de SARM, perteneciente a la colección de cultivos del Laboratorio de Microbiología del Hospital Amalia Simoni de la provincia Camagüey, aislada de secreciones purulentas en una herida quirúrgica, procedente de un paciente ingresado en la Unidad de Cuidados Intensivos (UCI).
El universo quedó constituido por todas las réplicas (cultivos) de una cepa salvaje de SARM, obtenida a partir de muestras clínicas hospitalarias, conservadas por el método de congelación en glicerol, 14 después de su identificación fenotípica mediante los métodos convencionales, 15) y por las réplicas de la cepa de referencia ATCC 25923, procedente de la American Type Culture Colletion (ATCC, por sus siglas en inglés), empleadas para el control de calidad de los medios de cultivo y como referente para la comparación en las reacciones de las pruebas fisiológicas realizadas a la cepa salvaje, que cumplieron los criterios de inclusión y exclusión establecidos por los autores del trabajo.
Criterios de inclusión:
- Réplicas de la cepa salvaje de SARM, conservada en el Laboratorio de Microbiología del Hospital Amalia Simoni.
- Cultivos biológicos de la cepa de referencia ATCC 25923, conservada en el Laboratorio de Microbiología del Hospital Amalia Simoni.
Citerios de exclusión:
- Réplicas (cultivos) de SARM contaminados con hongos u otras bacterias.
Cálculo de la muestra:
Para el cálculo del tamaño muestral se empleó el programa estadístico Epidat. La muestra quedó conformada por un total de 60 réplicas, 30 correspondieron a la cepa salvaje de SARM, conservada en la colección de cultivos del Laboratorio de Microbiología, después de su identificación y 30 réplicas pertenecieron a la cepa de referencia ATCC 259236. De esta manera se conformaron los dos grupos de estudio.
Identificación y confirmación de la cepa salvaje: se utilizaron los métodos convencionales de diagnóstico microbiológico disponibles: morfología colonial, observación microscópica de la agrupación y morfología del microorganismo, en respuesta a la coloración de Gram, producción de coagulasa y catalasa, así como la fermentación del manitol. 15) Se realizó la prueba de susceptibilidad por el método de difusión con discos o método de Kirby-Bauer, en agar Mueller-Hinton, 16 para definir la cepa como SARM se emplearon discos de cefoxitina (FOX) de 30 µg. Según lo normado por el Instituto de Normas Clínicas y de Laboratorio (CLSI, por sus siglas en inglés Clinical and Laboratory Standard Institute), los diámetros de los halos de inhibición para los discos de cefoxitina (30 µg), se consideraron sensibles cuando la medición de los mismos fue igual o mayor que 22 mm y resistentes cuando fue igual o menor que 21 mm. (17) Todos los discos provenían de la firma comercial Liofilchem. Para el control de la calidad del proceso se utilizó la cepa de referencia Staphylococcus aureus ATCC 25923.
La conservación de las cepas se realizó por el método de cultivos semistock o método de congelación en gicerol, para el cual se realizó una suspensión de la colonia de S. aureus en caldo Brucella, con 15 % de glicerol. Esta suspensión se dispensó en viales de crioconservación de 0,5-1mL y se congelaron a una temperatura entre -10 a -25 °C. El método permite la conservación de bacterias y levaduras en durante años o al menos de 6 a 12 meses. (14
Método para medir la actividad antimicrobiana de la peroxiadenosina: una vez identificadas la cepa salvaje y de referencia, se midió la susceptibilidad microbiana frente a la peroxiadenosina mediante el método de difusión con discos (Kirby-Bauer), en agar Mueller-Hinton. En cada placa se utilizaron controles positivos (C+, disco de doxiciclina), controles negativos (C-, discos impregnados en agua destilada estéril, 2,5 µL) y controles de esterilidad (CE, discos estériles). La peroxiadenosina (2,5 µL) se impregnó en discos de papel de filtro (D1, D2, D3, D4, D5) para estudiar su actividad.
Las categorías de susceptibilidad a la peroxiadenosina se interpretaron, en correspondencia con los criterios establecidos por el CLSI, observados en las pruebas de control de calidad para la doxiciclina (C+) frente a Staphylococcus aureus ATCC 25923. Se consideraron resistentes aquellas cepas cuyo halo de inhibición fue menor o igual que 22 mm y sensibles, las que mostraron un halo de inhibición mayor o igual que 23 mm. (18,19
Procedimiento: la solución de la peroxiadenosina sin diluir se preparó a partir de 5 µL de peróxido de hidrógeno al 30 %, que reaccionó con la adenosina (3 mg por mL) a la temperatura ambiente (18-24 0C) y pH de 7,30-7,40. A continuación se realizaron diluciones dobles seriadas de la peroxiadenosina en agua destilada estéril hasta alcanzar la concentración 1:32. Se usó la peroxiadenosina sin diluir y las diluciones 1:2, 1:4, 1:8, 1:16 y 1:32. Se identificó la formación de peroxiadenosina mediante los métodos colorimétricos y espectrofotométricos. 20
Luego, a partir de un cultivo puro, mediante un asa o aguja para inocular, se tomaron entre cuatro y seis colonias ya identificadas como S. aureus estas se transfirieron a un tubo que contenía entre 2 a 5 mL de caldo Mueller-Hinton; la suspensión obtenida se incubó durante dos a cinco horas a la temperatura entre 35 a 37 °C, hasta alcanzar la fase exponencial de crecimiento. La turbidez se verificó mediante su comparación con el tubo equivalente al patrón 0,5 de la escala McFarland y se ajustó con caldo Mueller-Hinton cuando fue necesario. Este procedimiento fue similar para ambas cepas de S. aureus investigadas.
Una vez obtenidas las réplicas de ambas cepas, estas se sembraron en placas de Petri, conteniendo agar Mueller-Hinton y en cada cultivo se aplicaron los discos impregnados con las diferentes concentraciones de la peroxiadenosina, para medir la actividad antimicrobiana de la misma. Esta actividad se midió en intragrupos (grupos de réplicas tratadas con la misma concentración de peroxiadenosina) e intergrupos (grupos de réplicas tratados con diferentes concentraciones de peroxiadenosina). Los cultivos se incubaron durante 18-24 horas a una temperatura entre 35 a 37 °C y posterior a la incubación, se realizó la lectura. Los resultados se interpretaron, según los criterios recomendados por el CLSI ya mencionados. 18,19
Preparación y distribución de los discos de papel de filtro: los discos impregnados con peroxiadenosina se elaboraron por los autores del trabajo, a través del procedimiento siguiente: en una placa de Petri estéril se distribuyeron los discos de papel de filtro de porosidad #1 y se esterilizaron en autoclave, luego se les añadió 2,5 µL de peroxiadenosina a una concentración de 3 mg/mL. En cada placa los discos se colocaron a una distancia de 1,5 cm del borde de la placa y 2,5 cm entre los discos, para evitar la superposición de los halos de inhibición. En dependencia del tamaño de la placa (100 X 13 mm y 150 mm) se calculó y dispuso el número de discos aplicados.
Se estudiaron las siguientes variables dependientes: la formación de la peroxiadenosina, la preservación de la estructura aromática de la adenosina y el patrón de susceptibilidad de SARM al antimicrobiano objeto de estudio. Las variables independientes incluyeron a la concentración de la peroxiadenosina y la solución de estudio.
Los hallazgos obtenidos se recogieron en una base de datos de Microsoft Excel, se procesaron por medios computarizados, con la ayuda del paquete estadístico SPSS versión 21.00, compatible con el sistema operativo Windows XP. Se realizó un análisis de la varianza (ANOVA), una prueba z de ubicación de la media de una muestra y se plasmaron en valores absolutos mediante distribución de la frecuencia y los porcentajes. Los resultados se muestran en las tablas y el texto, en correspondencia con los objetivos propuestos.
En el desarrollo de la investigación se mantuvo como premisa, respetar los principios bioéticos implícitos en los estudios de este tipo y los resultados se usaron con fines investigativos. Los investigadores velaron por la calidad de los resultados mediante el cumplimiento de los procedimientos normalizados para cada una de las determinaciones.
Resultados
El peróxido de hidrógeno en una solución concentrada de 56,7 mmol/L, reaccionó con las moléculas de adenosina y su concentración se redujo a 11,8 mmol/L, alrededor de la quinta parte de su concentración inicial. Esta observación indicó la síntesis de peroxiadenosina con un rendimiento mayor al 80 %.
La comparación respecto al grado de absorbancia de la luz ultravioleta determinado mediante espectrofotometría entre la adenosina y el producto de la reacción peroxiadenosina, reveló que, la densidad óptica (DO) fue de 1,564 para ambos productos; esta observación indicó que el proceso de peroxidación no afectó la capacidad para la absorbancia de luz a 260 nm.
La concentración sin diluir de la peroxiadenosina, generó un halo de inhibición del crecimiento de SARM promedio de 30 mm, que se correspondió con la mayor medida registrada en este estudio. Mientras que las concentraciones de peroxiadenosina 1:16 y 1:32, mostraron los halos de inhibición más pequeños (6,6 y 6,0 mm), respectivamente. Estos últimos halos se correspondieron con las menores medidas tomadas (Tabla 1).
Tabla 1 Patrón de sensibilidad, según la concentración de peroxiadenosina. Hospital Amalia Simoni. Enero de 2016-Julio de 2020
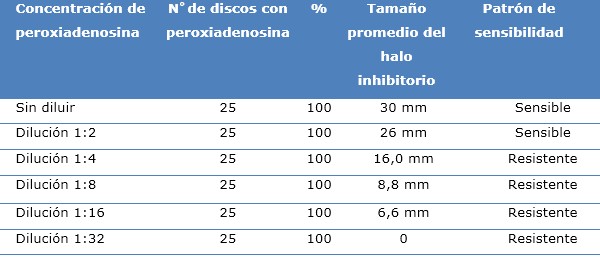
Fuente: base de datos Microsoft Excel del estudio preclínico.
En la Figura 1 se muestran los halos de inhibición generados por los discos de peroxiadenosina sin diluir, cuya concentración fue de 3 mg/mL. El diámetro promedio de estos halos de inhibición con los discos de peroxiadenosina fue de 30 mm, similar al producido por el disco de doxiciclina empleado como control positivo (C+), para medir la actividad antimicrobiana de la peroxiadenosina en ambas cepas (Figura 1).
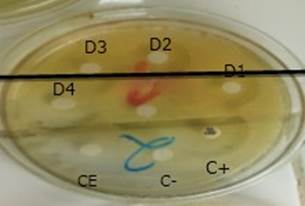
Figura 1 Halos de inhibición generados por los discos de peroxiadenosina sin diluir, en cultivos de SARM. Se observan ocho discos, en una placa Petri de 110 mm, con una separación de 1,5 cm del borde y 2,5 cm entre los discos. Se observa el disco de doxiciclina (C+) y a la izquierda de este los controles negativos y de esterilidad, respectivamente. En la parte superior de la placa y en contra de las manecillas del reloj, se enumeran los cinco discos impregnados en peroxiadenosina.
También se muestran las diferencias entre las medias aritméticas del halo de inhibición generadas por las diferentes concentraciones de la peroxiadenosina en las réplicas de SARM in vitro. Nótese que, en las réplicas intragrupos (grupos de cultivos tratados con la misma concentración) las diferencias observadas en los halos de inhibición generados por las concentraciones de peroxiadenosina fueron significativas; mientras que, en las intergrupos (grupos de cultivos tratados con diferentes concentraciones) la diferencia significativa fue mayor.
El programa estadístico de Fisher presentó una probabilidad de error inferior a 0,05 y un intervalo de confianza de 95 %, lo cual determinó con alta confiabilidad, la existencia de diferencias en los grupos estudiados (Tabla 2).
Tabla 2 Comparaciones múltiples de las medias aritméticas identificadas en los halos de inhibición

Fuente: base de datos Microsoft Excel del estudio preclínico.
Estadística de Fisher: 1078,045 p = 0,000
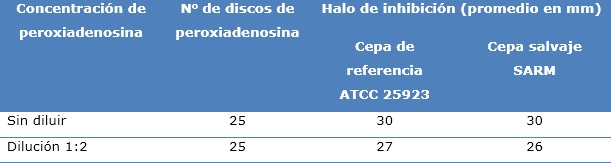
A continuación se muestra una comparación de la actividad antimicrobiana generada en las 30 réplicas correspondientes a la cepa salvaje de SARM y en las 30 réplicas de la cepa de referencia investigada. Se observó que, a la concentración de peroxiadenosina sin diluir, el halo de inhibición generado en ambas cepas fue de 30 mm, mientras que, a la concentración 1:2, el halo de inhibición de la cepa de referencia fue mayor.
Estos apuntes indican que la peroxiadenosina mostró actividad antimicrobiana frente a S. aureus sin tener en cuenta el origen, los cambios genotípicos y fenotípicos de la especie (Tabla 3).
DISCUSIÓN
Respecto a la oxidación de la estructura aromática con el peróxido de hidrógeno, un estudio realizado por Hernández Ramírez MG y Alegría Torres JA, 21) en Guanajuato, México durante el año 2018, al referirse a la determinación de la 8-hidroxideoxiguanosina en el ácido desoxirribonucleico (ADN) mitocondrial, mediante la reacción en cadena de la polimerasa en tiempo real (PCR-RT), se evidencia que, el estrés oxidativo daña al ADN por la oxidación de la base guanina y la subsecuente formación de 8-hidroxideoxiguanosina (8-OHdG), que conserva sus propiedades ópticas observaciones relacionadas con las obtenidas en el estudio.
Además, se conoce que los compuestos fenólicos tienen una estructura aromática, se encuentran en el follaje de muchas plantas y tienen actividad antioxidante, es decir, reaccionan con el peróxido de hidrógeno. Estos apuntes se comprueban en un estudio realizado por Calderón A et al. (22 con un extracto hidroalcohólico de hojas de Terminalia catappa, ensayado in vivo para determinar su acción antioxidante frente a los radicales libres inducidos en el cerebro de ratas. Se determina que el grupo tratado con el extracto hidroalcohólico a la concentración de 0,8 mg/Kg de peso corporal muestra una inhibición significativa de los radicales libres cuando se compara con el subgrupo control positivo. Estos resultados están en correspondencia con los obtenidos en la investigación, en relación con la peroxidación de estructuras aromáticas sin afectar sus propiedades resonantes.
En relación con la absorbancia de la luz, las observaciones obtenidas en el trabajo fueron similares a las señaladas por Cardellá Rosales L y Hernández Fernández R, (23) sobre los nucleótidos y nucleósidos de adenosina; esos autores confirman que, los nucleósidos son capaces de absorber la luz ultravioleta a 260 nm de longitud de onda, observándose como manchas oscuras sobre el papel de acetato. En la bibliografía consultada Baynes JW y Dominiczac MH, (24) reconocen de manera análoga que la causa de este fenómeno se debe a la aromaticidad de la molécula.
La amplificación o replicación de la molécula de ADN, consiste en la capacidad que tiene una molécula madre para producir dos moléculas hijas idénticas en toda su secuencia. Una forma de amplificar el ADN in vitro es a través de la técnica de PCR, donde, la temperatura de hibridación de los partidores juega un rol importante a la hora de amplificar un gen. La técnica de PCR requiere en algún momento determinar la concentración de genes amplificados y para ello se auxilia de la espectrofotometría que le permite cuantificar el ADN mediante el principio de absorbancia aplicado a las bases nitrogenadas a 260 nm de longitud de onda, observación que se correspondió con los hallazgos citados en el estudio. (25
El método de difusión en agar mediante el uso de discos de antimicrobianos o método de Bauer Kirby, detecta la mayoría de los SARM, pero en algunos de estos aislamientos se necesita su confirmación. El método no es confiable para la detección de S. aureus con resistencia intermedia a la vancomicina, ya que no diferencia estas cepas de otras sensibles. El patrón de susceptibilidad es una variable que puede cambiar en dependencia del microorganismo y la concentración del antimicrobiano. (26
En el trabajo, la cepa salvaje de SARM se confirmó mediante el estudio de la susceptibilidad antimicrobiana con disco de cefoxitina, que generó un halo inhibitorio ≤ 21 mm en los medios de cultivo, sin embargo, en presencia de peroxiadenosina la cepa de SARM se mostró sensible, evidenciado por un halo inhibitorio de 30 mm de diámetro. Estos apuntes se correspondieron con los datos ofrecidos por Sáinz-Rodríguez R et al. 27 en el Hospital Regional Universitario de Málaga, con el objetivo de evaluar una prueba rápida para la detección de PBP2a en S. aureus, donde evidencian que, el método de difusión en agar con disco de cefoxitina es muy útil en la detección y confirmación de poblaciones de SARM, con un 100 % de sensibilidad y especificidad, tras el análisis de 107 aislamientos de S. aureus a partir de muestras clínicas y frotis nasales.
Estos datos sugieren que la peroxiadenosina presenta una actividad antimicrobiana superior a la cefoxitina y similar a la doxiciclina (C+) en los cultivos in vitro de SARM. Una causa que podría explicar este comportamiento es la diferencia en el mecanismo de acción de estos antimicrobianos, pero se desconoce el mecanismo responsable en la peroxiadenosina, por lo que habrá que profundizar en este sentido.
Una investigación en el año 2017, dirigida por Aties López L et al. 28 refieren que, en 47 aislamientos de estafilococos procedentes de muestras biológicas investigadas en el Laboratorio de Microbiología del Hospital Provincial Docente Clínico Quirúrgico Saturnino Lora Torres de Santiago de Cuba, desde febrero hasta abril de 2017, con el objetivo de determinar la frecuencia de SARM, aporta los hallazgos siguientes: del total de aislamientos en los que utilizan el disco de cefoxitina, el 85,7 % de S. aureus muestran resistencia (halo de inhibición menor de 14 mm), frente al 75 % de los estafilococos coagulasa negativa.
Estos datos refuerzan las evidencias respecto a la utilidad del método de difusión en agar con discos de cefoxitina para la confirmación de SARM, lo cual se ajustó a las observaciones recogidas en la actual investigación y coincidió con los criterios de Sáinz-Rodríguez R et al. (27 emitidos en este sentido.
Asimismo, un estudio descriptivo de serie de casos, en la terapia pediátrica infantil del Hospital Comandante Pinares, de San Cristóbal, Artemisa, asevera que la susceptibilidad a la meticilina se determina mediante pruebas de difusión con discos de cefoxitina de 30 ug, en el medio de agar Mueller Hinton, incubado a 37 °C durante 24 horas, punto de vista defendido por los autores del experimento, quienes emplearon el mismo método para la confirmación de SARM en los cultivos analizados. Hernández Loriga W et al. (29) refieren que, de una selección de cinco pacientes en quienes aíslan S. aureus por cultivos microbiológicos (hemocultivo y cultivos de secreciones), todos los cultivos muestran halos de inhibición menores de 20 mm, considerado un parámetro de resistencia y confirmación de SARM.
Al comparar el halo inhibitorio promedio generado por los discos de peroxiadenosina sin diluir (halo de 30 mm), con los producidos por los discos de cefoxitina de 30 µg (20 mm) frente al mismo microorganismo y mediante un método similar, se puede inferir que la peroxiadenosina en cultivo in vitro presenta mayor actividad antimicrobiana.
Los hallazgos estadísticos indicaron que, la peroxiadenosina a la concentración sin diluir generó un halo inhibitorio de 30 mm como promedio, lo cual evidenció la alta efectividad de la concentración frente a las cepas de SARM estudiadas, no se observó diferencias significativas entre los halos de inhibición de ambas cepas. Los datos tuvieron una distribución normal, se trabajó con una confiabilidad del 95 % y se aceptó una probabilidad de error de 0,05 durante el análisis de la varianza.
Por las observaciones realizadas en el estudio se puede generalizar que la peroxiadenosina tiene actividad antimicrobiana frente a S. aureus, la cual no parece depender del sitio de aislamiento ni de diferencias genotípicas o fenotípicas, tal como se observó entre una cepa salvaje y otra de referencia.
Conclusiones
La colorimetría y la espectrofotometría constituyen métodos útiles para confirmar la formación y preservación de la estructura aromática de la peroxiadenosina.
El crecimiento in vitro de S. aureus se inhibe en presencia de la peroxiadenosina. Existe una relación lineal ascendente entre las concentraciones de peroxiadenosina y el halo de inhibición generado en el cultivo in vitro de SARM.
La inhibición del crecimiento, por la peroxiadenosina de ambas réplicas de SARM investigadas, sugiere continuar su evaluación como un nuevo producto antibacteriano eficaz contra este microorganismo

