










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936
cultrop v.29 n.4 La Habana oct.-dic. 2008
Estabilidad de la eficiencia de la cepa Glomus mosseae en la respuesta del tomate a condiciones de estrés hídrico fuera de su periodo óptimo
Ms.C. L. R. Fundora. Email: lrfundora@inca.edu.cuI, Aracely MenaI, Ms.C. Yaquelín RodríguezII, Ms.C. P. J. GonzálezIII, Dr.C. P. RodríguezIV, Dianevys González-PeñaV
I Especialistas
II Investigador Agregado
III Investigador Auxiliar del departamento de Biofertilizantes y Nutrición de las Plantas
IV Investigador Agregado
V Reserva Científica del departamento de Fisiología y Bioquímica Vegetal, Instituto Nacional de Ciencias Agrícolas (INCA), Gaveta Postal 1, San José de las Lajas, La Habana, Cuba, CP 32 700.
ABSTRACT
Among the ecological alternatives reducing water stress effects, the arbuscular mycorrhizal fungal inoculation has been put into practice. Deepening into this subject, the work was aimed to test the stability of G. mosseae efficiency on tomato behaviour, growing out of its optimal period under water deficit conditions. Thus, two experiments were carried out under semi-controlled conditions, within half-late and late periods for tomato crop, inoculated and non-inoculated plants being subjected to water stress. Some variables related to mycorrhizal activity, plant growth and its water status were determined. The following indicators were evaluated: root colonization per arbuscular mycorrhizal fungi (% Col), visual density (DV), total dry weight as well as of leaves, stems and roots (g), foliar area (Sf) and water potential (y). Results proved strain efficiency per each indicator evaluated, which were higher in inoculated plants compared to non-inoculated ones, even though they were cultivated out of the optimal period. This experience suggests certain strain-crop specificity and particularly the best tomato development in symbiosis with G. mosseae under the studied conditions.
Key words: fungi, Glomus, tomato, drought stress, plant response
RESUMEN
Entre las alternativas ecológicas para disminuir las afectaciones causadas por el estrés hídrico, se ha puesto en práctica la inoculación con hongos micorrízicos arbusculares. Profundizando en la temática se realizó este trabajo, con el objetivo de comprobar la estabilidad de la eficiencia de G. mosseae en el comportamiento del tomate cultivado fuera de su período óptimo sometido a condiciones de déficit hídrico. Para ello se llevaron a cabo dos experimentos en condiciones semicontroladas, en períodos medio tardío y tardío para el cultivo del tomate, sometiéndose a estrés hídrico las plantas inoculadas y sin inocular. Se determinaron algunas variables relacionadas con la actividad micorrízica, el crecimiento y estado hídrico de las plantas. Los indicadores evaluados fueron la colonización radical por los hongos micorrízicos arbusculares (% Col), densidad visual (DV), masa seca de hojas, tallos, raíz y total (g), superficie foliar (Sf) y el potencial hídrico (y). Los resultados demostraron la efectividad de la cepa para cada uno de los indicadores evaluados, siendo superiores en las plantas inoculadas en relación con las no inoculadas, aun cuando se cultiva fuera de su período óptimo. Esta experiencia sugiere cierta especificidad cepa-cultivo y, en particular, el mejor desarrollo del tomate en simbiosis con G. mosseae en las condiciones estudiadas.
Palabras clave: hongos, Glomus, tomate, estrés de sequía, respuesta de la planta
INTRODUCCIÓN
El tomate (Solanum lycopersicon M.) es considerado uno de los cultivos hortícolas de mayor importancia a escala mundial, su demanda aumenta continuamente y con ella su producción y comercio (1). La producción actual está comprendida en más de 124 millones de TM en un área de alrededor de 4, 518,529 millones de hectáreas. En Cuba, se cultivan 60,000 ha de tomate con una producción que alcanza las 800,000 TM (2).
Las condiciones climáticas que a menudo prevalecen en el Caribe no se corresponden con las exigencias ecológicas del tomate. Por otra parte, las prácticas de manejo agrotécnico no se han desarrollado acorde con la problemática específica (3).
Cuba ha puesto en práctica el uso de biofertilizantes producidos a partir de hongos micorrízicos arbusculares (HMA) (4), los cuales contribuyen de forma más eficiente a su supervivencia y crecimiento, al reducir los estrés asociados con la nutrición, las relaciones con el agua, la estructura del suelo, el pH, las sales, los metales tóxicos y los patógenos (5, 6, 7, 8).
Se ha informado el efecto beneficioso de la simbiosis micorrízica sobre la tolerancia de las plantas ante condiciones de estrés hídrico (9, 10, 11, 12, 13); sin embargo, aún quedan interrogantes que no están esclarecidas.
Los estudios realizados sobre el comportamiento del tomate al ser inoculado con distintas cepas de HMA, provenientes del cepario del Instituto Nacional de Ciencias Agrícolas (INCA), han demostrado un mejor funcionamiento del cultivo en simbiosis con la cepa Glomus mosseae (14, 15). En condiciones de déficit hídrico, esta cepa ha resultado ser la más promisoria, garantizando un mejor aprovechamiento del agua en plantas expuestas a condiciones de estrés (16).
De esta manera, se decide profundizar en la temática analizando la estabilidad de la eficiencia de la cepa G. mosseae, respecto al crecimiento y desarrollo del tomate en un período no óptimo, sometido a condiciones de déficit hídrico.
MATERIALES Y MÉTODOS
Los experimentos se realizaron en condiciones semicontroladas en el área central del INCA, ubicadas bajo un techo de cristal para evitar la influencia de las precipitaciones; el primero en un período medio tardío, que abarcó desde la segunda quincena de diciembre de 2004 y culminó en la primera quincena de febrero de 2005. Y el segundo en el período considerado tardío para el desarrollo del cultivo, que comprendió la primera quincena de marzo de 2005 para finalizarlo en la segunda quincena de abril de ese mismo año.
La especie utilizada fue Solanum lycopersicon M, variedad Amalia, teniendo en cuenta la estabilidad de su potencial productivo al ser cultivada fuera del período óptimo. Las semillas se sembraron directamente en macetas de 0,5 L de capacidad, que contenían una mezcla de suelo Ferralítico Rojo Lixiviado (17) y cachaza en proporción 3:1v/v, y se utilizaron 30 recipientes por tratamiento con una planta por recipiente.
Antes de efectuar la siembra se aplicaron dos gramos de inóculo certificado de la cepa Glomus mosseae, perteneciente al cepario del departamento de Biofertilizantes y Nutrición de las Plantas, con el objetivo de establecer la infección micorrízica. El inóculo contenía 140 esporas promedio por gramo de suelo.
El riego se realizó de forma manual para todos los tratamientos, garantizando la humedad del sustrato acorde con las exigencias del cultivo. Al alcanzar las plantas un estado fisiológico de siete-ocho hojas verdaderas emitidas, se establecieron los tratamientos que se muestran en la Tabla I.
El tratamiento de estrés se realizó a través de la suspensión del suministro hídrico a las plantas por espacio de 10 días, coincidiendo con la aparición visual de síntomas severos de marchitez. Las evaluaciones de los indicadores de infección micorrízica, el crecimiento y las relaciones hídricas se realizaron al finalizar el período de estrés hídrico.
Los indicadores evaluados fueron: la colonización radical por los hongos micorrízicos arbusculares (% C) a través de la técnica de tinción con Azul Trypan y para ello se evaluaron cuatro plantas por tratamiento (18) y la densidad visual (DV) según metodología propuesta (19). La evaluación del crecimiento en masa seca de la raíz y total g.planta-1 se realizó mediante el secado a la estufa a 800C utilizando cinco plantas por tratamiento; la superficie foliar por el método del disco (10) que se calculó mediante la siguiente expresión:
Sf=(Ad x Msf)/(Msd)
donde:
Sf- superficie foliar expresada (cm2)
Ad- área de 40 discos (cm2)
Msf- masa seca foliar de la planta (g)
Msd- masa seca de los discos (g)
Las medidas del potencial hídrico foliar (y) MPa se determinaron con una cámara de presión tipo Sholander según la metodología (20); esta medición se realizó al alba y se hicieron cinco mediciones por tratamiento.
Los tratamientos fueron distribuidos siguiendo un modelo completamente aleatorizado bajo arreglo bifactorial (2x2). El tratamiento estadístico de los datos se realizó mediante un Análisis Factorial, para determinar la interacción entre los factores. A los indicadores que mostraron diferencias entre los tratamientos se les aplicó la Prueba de Comparación de Rangos Múltiples de Duncan, (21), con p£0.05%, utilizando el programa estadístico SPSS 11.5 para Windows. Los resultados del porcentaje de colonización fúngica fueron primeramente transformados por la función 2arcsenvx de acuerdo con Lerch (22).
RESULTADOS Y DISCUSIÓN
El análisis factorial realizado en ambos experimentos, a los indicadores de crecimiento y colonización radical (Tablas II y III), demostró que no existía interacción entres los factores en estudio, por lo que la respuesta de las plantas a la biofertilización con HMA sería la misma tanto para las plantas estresadas como para las que tienen riego sistemático.


Esto permitió la comparación de los tratamientos por separado en función del suministro hídrico. Sin embargo, el potencial hídrico foliar demostró que sí existía interacción entre los factores, por lo que se compararon los cuatros tratamientos entre sí.
El porcentaje de colonización micorrízica en las raíces de las plantas (Figura 1) muestra valores adecuados al cultivo en las plantas inoculadas, en los dos experimentos. La presencia de cepas nativas de HMA en el sustrato fueron capaces de colonizar el sistema radical en un 20 %.
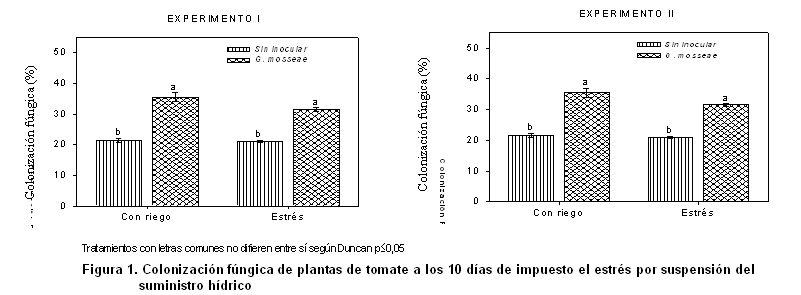
Sin embargo, ante la condición de estrés, se produjo un efecto depresivo en la infección micorrizógena, lo cual pudo estar determinado por la detección del ritmo metabólico provocado por la deficiencia hídrica; mientras que para las plantas sin inocular, estas alcanzaron niveles estables de colonización, lo cual se le atribuye a que las cepas nativas presentes en el sustrato hayan creado un mecanismo de adaptación a las condiciones de estrés presentes en este estudio.
En el experimento I se observa que la densidad visual (Figura 2) no mostró diferencias entre los tratamientos estudiados; este comportamiento pudiera estar dado por el efecto de las cepas nativas, las cuales manifiestan el mismo grado de intensidad en ambas condiciones. En cambio, en el experimento II se encontró un incremento significativo en las raíces inoculadas con la cepa G. mosseae, tanto en las plantas bien abastecidas de agua como en las estresadas. No obstante, los valores de densidad visual de las plantas sin inocular en este experimento fueron inferiores si se compara con los encontrados en el experimento I.
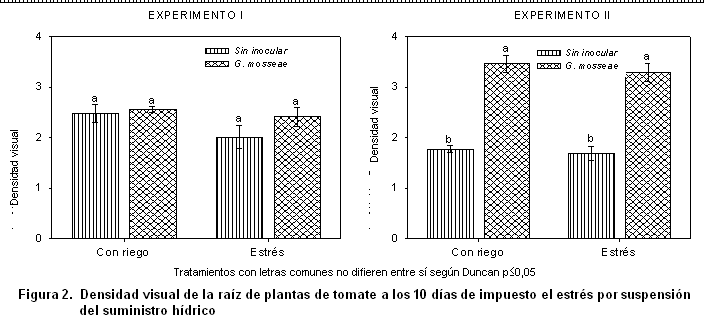
Se deben tener en cuenta las condiciones climáticas específicas del período experimental II (primavera-verano), que al ser no idóneas para el buen desarrollo del tomate, pueden haber alterado la relación genotipo-ambiente del cultivo y ser las responsables del comportamiento de la simbiosis, unido a que la influencia de la falta de agua sobre la infección micorrízica dentro y fuera de la raíz es diferente. Además, es conocido que la simbiosis micorrízica es más aceptada y muestra un mejor funcionamiento cuando las condiciones del cultivo hospedero no son las ideales, tal y como sucede en este caso con el tomate al ser cultivado en el período no óptimo para su desarrollo.
La micorrización también favoreció la acumulación de la biomasa seca total de las plantas (Figura 3), observándose su efecto para condiciones de control como para condiciones de estrés en las plantas inoculadas en ambos períodos.
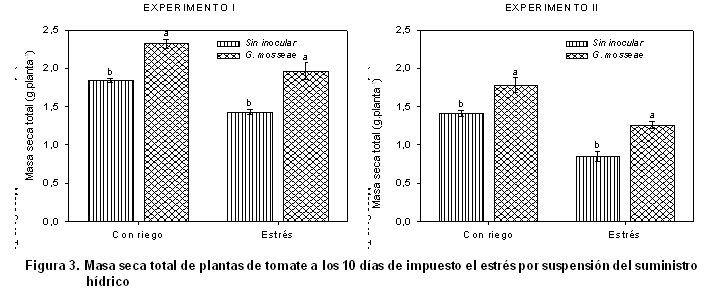
En el experimento 2 se observa cómo hubo una reducción de este indicador para ambas condiciones, debido a que se desarrolló en período no óptimo; aunque la respuesta siempre fue significativamente superior en las plantas inoculadas que en las no inoculadas, demostrándose de esta forma la estabilidad de la cepa Glomus mosseae; estos resultados se corroboran con los obtenidos anteriormente (23).
Al analizar el efecto de la micorrización en la biomasa seca de las raíces (Figura 4), solo se observaron diferencias significativas a favor de las plantas inoculadas, en ambos períodos y condiciones de abastecimiento hídrico.
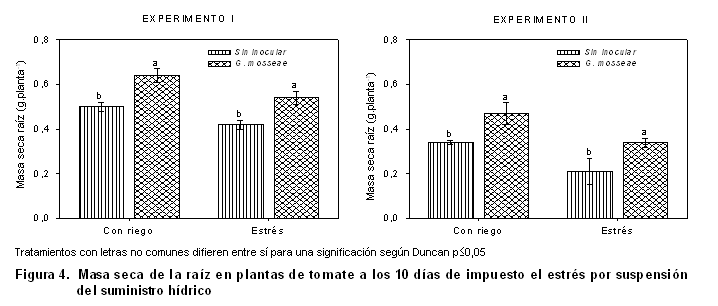
Estos resultados demuestran cómo la micorrización no provoca cambios significativos en el crecimiento de la raíz, pero estimula la acumulación de biomasa seca y el área foliar de las plantas sometidas a ambas condiciones hídricas.
La superficie foliar (Figura 5), por lo general, es uno de los indicadores que más se ajusta o se adapta a los cambios climáticos, sobre todo a aquellos relacionados con la humedad y temperatura del aire, quizás por ser el órgano más expuesto a la interacción con estos factores o por las características propias del tejido foliar. En ambos experimentos se notó cómo los valores fueron relativamente bajos, lo cual es interesante precisar que la mayor reducción en este indicador se detectó en las condiciones de estrés sin inoculación micorrízica del experimento II, quedando asociado este resultado al tratamiento impuesto y al período en que se desarrolló el cultivo, tal y como se ha venido resaltando.
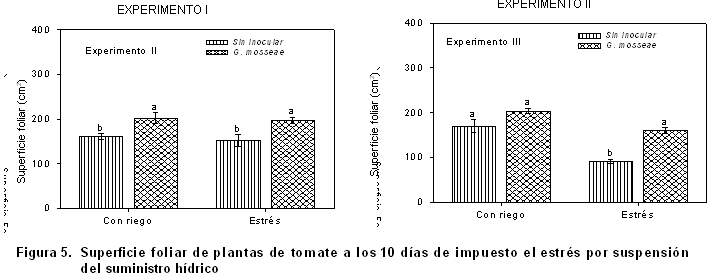
Algunos plantean que, en condiciones de estrés hídrico, las micorrizas juegan un papel fundamental en activar la difusión de iones y agua hacia las raíces en las plantas hospederas (23), mejorando la nutrición nitrogenada y absorción de fósforo, lo que permite a las plantas soportar las condiciones de sequía y recuperarse con mayor facilidad.
Estudiando el efecto de los HMA sobre el crecimiento de plantas de tomate (15), se informó que las variables de crecimiento evaluadas presentaron valores superiores en las plantas inoculadas en relación con el tratamiento sin inocular, siendo la superficie foliar la que mejor reflejó este efecto. También se incrementó la masa seca del tallo de las plantas colonizadas, resultado que está en correspondencia con los obtenidos en esta experiencia. Por otra parte, en un cultivar de Pinus halepensis se obtuvieron ligeros incrementos a favor de las plantas colonizadas en las variables altura y masa seca total (24).
El estado hídrico de las plantas se refleja a través del potencial hídrico (y) medido al alba al final de impuesto el período de estrés (Figura 6), donde las plantas que se abastecieron de agua de forma constante mantuvieron potenciales elevados sin que se apreciaran diferencias por causa de la inoculación micorrízica. En cambio, en las plantas estresadas hídricamente sí se observó el efecto de la inoculación con la cepa G. mosseae, que fue capaz de incrementar el potencial hídrico foliar y, por tanto, reducir las pérdidas de agua en estas plantas respecto a las no inoculadas, por lo que la simbiosis micorrízica continuó revelando su efectividad incluso al desarrollarse el cultivo fuera de su época óptima.
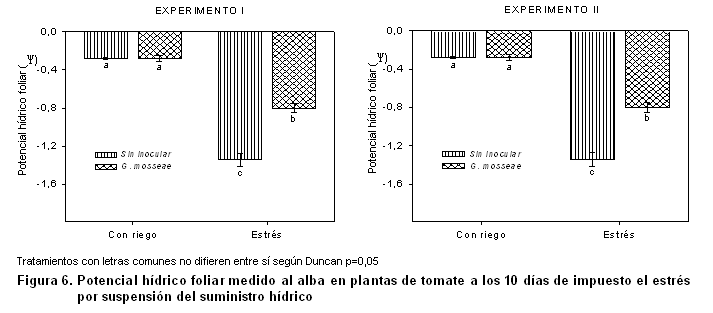
En estudios realizados en plantas de tomate en condiciones de estrés hídrico (11), también se encontró que las plantas inoculadas con la cepa G. mosseae presentaron valores superiores de potencial hídrico foliar y potencial de turgencia que las no inoculadas. Es de señalar que entre los dos tratamientos que estaban bien abastecidos de agua, los valores del potencial hídrico no evidenciaron diferencias significativas, coincidiendo con los resultados observados en este trabajo.
La colonización de las raíces por los HMA puede mejorar las relaciones hídricas y la resistencia a la sequía de las plantas hospederas, siendo el potencial hídrico uno de los indicadores más estudiados (9, 25). Ellos han encontrado resultados a favor de las plantas colonizadas por estos hongos, tal y como se ha corroborado en nuestras condiciones en estos experimentos y en otros previos realizados por el autor.
Es de interés destacar que las plantas de tomate, tanto en condiciones o no de estrés, se vieron favorecidas en su crecimiento y desarrollo por el efecto de la inoculación, ya que los indicadores evaluados mostraron mayores valores en comparación con las plantas no inoculadas. Este comportamiento puede deberse a que las hifas del hongo mejoran indicadores como la conductividad hidráulica de la raíz, lo cual disminuye la resistencia de esta al paso del agua, aspecto que ha sido comprobado (26) en trabajos de semillero de Ulmus americana.
Resultó positiva la inoculación con los HMA para mejorar las relaciones hídricas en condiciones estresantes, pues a pesar de que hubo una buena colonización con HMA nativos, no se encontró un mejor efecto de estos sobre el cultivo en los indicadores evaluados, lo cual denota la importancia que representa continuar trabajando con especies que sean eficientes, así como seguir investigando con otras cepas que puedan proporcionar beneficios similares o superiores a los encontrados en el presente trabajo.
Se plantea que las plantas tienen una serie de respuestas llamadas mecanismos de resistencia a la sequía, por medio de los cuales sobreviven a períodos de estrés hídrico (27); dentro de estos mecanismos están la reducción de la pérdida de agua debido a la resistencia estomática, reducción de la absorción de la radiación por cambios en la orientación de las hojas o reducción de la superpie foliar, los incrementos en la absorción de agua motivados por el desarrollo radical o por incremento en la conductancia de la raíz, y el mantenimiento de la turgencia debido al ajuste osmótico o a un incremento en la estabilidad celular. Se ha encontrado incidencia positiva de los HMA sobre estos mecanismos en plantas estresadas (10, 11, 12, 13).
Este estudio confirmó que los HMA ayudan a las plantas a tolerar más el déficit hídrico y, por tanto, a superar situaciones desfavorables, por sus efectos beneficiosos sobre la relación agua-planta; aunque siempre hay que tener en cuenta la especificidad de las cepas en relación con el tipo de suelo, cultivo y estrés a que se expongan las plantas.
En conclusión, quedó demostrada la estabilidad en la eficiencia simbiótica de la cepa G. mosseae en el crecimiento y desarrollo del tomate en condiciones de estrés hídrico, fundamentalmente cuando se cultiva fuera de su época óptima.
Referencias
1. Gómez, O. y Rodríguez, G. Imparto del cultivar en el sistema protegido de tomate (Lycopersicon esculentum, Mill). Conferencia, La Habana:IIHLD. 2004.
2. FAOSTAT. Última actualización febrero 2005. Consultado 4/03/05 Disponible en: <http://faostat/fao.org/faostat/notel/citacion.htm>. 2005.
3. Gómez, O. /et al./. Mejora genética y manejo del cultivo del tomate para la producción en el Caribe. La Habana. 2000, 159 p.
4. Rivera, R. Bases científico-técnicas para el manejo de los sistemas agrícolas micorrizados eficientemente. En: Los sistemas agrícolas micorrizados eficientemente, una contribución al desarrollo agrícola sostenible en el Caribe. 2003. 233 p.
5. Hutchinson, J. J.; Young, S. D.; Black, C. R. y West, H. M. Determining uptake of radio-labile soil cadmium by arbuscular mycorrhizal hyphae using isotopic dilution in a compartmented-pot system. New Phytologist, 2004, vol. 164, p. 477-484.
6. González-Monterrubio, C. F. Influencia de hongos micorrizógenos arbusculares (HMA) en el desarrollo de plántulas de Opuntia streptacantha Lem. sometidas a sequía, en condiciones de invernadero (Tesis profesional, Facultad de Estudios Superiores, Universidad Nacional Autónoma de México, México, D.F.), 2003. 66 p.
7. Rillig, M. C. y Mummey, D. L. Mycorrhizas and soil structure. New Phytologist, 2006, vol. 171, p. 41-53.
8. Ruiz-Lozano, J. M. Arbuscular mycorrhizal symbiosis and alleviation of osmotic stress. New perspectives for molecular studies. Mycorrhiza, 2003, vol. 13, no. 6, p. 309-317.
9. Barroso, L. Las relaciones hídricas en la albahaca blanca (Ocimum basilicum L) y su efecto en el crecimiento, desarrollo y la producción de aceites esenciales. [Tesis de doctorado]; INCA, 2004.
10. Dell’Amico, J.; Torrecillas, A.; Rodríguez, P.; Morales, D. y Sánchez-Blanco, M. J. Respuesta del tomate (Lycopersicon esculentum) a 36 horas de inundación en dos momentos del fotoperíodo. Cultivos Tropicales, 2002, vol. 3, no. 1, p. 29-34.
11. Marulanda, A.; Azcón, R. y Ruiz-Lozano, J. M. Contribution of six arbuscular mycorrhizal fungal isolates to water uptake by Lactuca sativa plants under drought stress. Physiologia Plantarum, 2003, vol. 119, p. 526-533.
12. Porcel, R.; Barea, J. M. y Ruiz-Lozano, J. M. Antioxidant activities in mycorrhizal soybean plants under drought stress and their possible relationship to the process of nodule senescence. New Phytologist, 2003, vol. 157, p. 135-143.
13. Porcel, R. y Ruiz-Lozano, J. M. Arbuscular mycorrhizal influence on leaf water potential, solute accumulation, and oxidative stress in soybean plants subjected to drought stress. Journal of Experimental Botany, 2004, vol. 55, no. 403, p. 1743-1750.
14. Terry, E. Microorganismos benéficos y productos bioactivos como alternativa para la producción ecológica del tomate (Lycopersicon esculentum, Mill, var. Amalia). [Tesis de doctorado]; INCA, 2004.
15. Rodríguez, Y.; Mazorra, L. M.; Fundora, L. R.; Noval, B. M.; Fernández, F. y Rodríguez, P. Water relations and biochemical changes in non-inoculated and AMF-inoculated tomato plants in response to drought. En: Conferencia Internacional de Micorrizas (ICOM4). (4:2003:Montreal), 2003. p. 731.
16. Fundora, L. R. La biofertilización micorrízica: una alternativa ecológica para cultivar tomate en condiciones de recursos hídricos limitados. [Tesis de Maestría]; UNAH, 2007.
17. Borges, Y. Contribución al estudio de la degradación de los suelos Ferralíticos Rojos Lixiviados por el cambio del uso de la tierra. [Tesis de Diploma]; UNAH. 2004, 67 p.
18. Phillips, J. M. y Hayman, D. E. Improved procedures for clearing roots and staining parasitic and vesicular-arbuscular mycorrhizal fungi for rapid assessment of infection. Trans. Br Mycol. Soc., 1970, vol. 55, p. 158-161.
19. Herrera, R. A. /et al./. Estrategia de funcionamiento de la micorrízas VA en un bosque tropical. Biodiversidad en Iberoamérica. Ecosistemas, evolución y procesos sociales. En: Programa Iberoamericano de Ciencia y Tecnología para el Desarrollo. Subprograma, Diversidad Biológica. (12:1995:Mérida), 1995.
20. Tuner, N. C. Measurement of plant water status by the pressure chamber technique. Irrigation Science, 1988, vol. 9, p. 289-308.
21. Cochran, W. y Cox, G. Diseños experimentales. C. México: Ed. Trellas, 1990, p. 132-135.
22. Lerch, G. La experimentación en las ciencias biológicas y agrícolas. Editorial Científico técnico, 1977, 452 p.
23. Kubikova, E.; Moore, J. L.; Ownley, B. H.; Mullen, M. D. y Augé, R. M. Mycorrhizal impact on osmotic adjustment in Ocimum basilicum during a lethal drying episode. Plant Physiol., 2001, vol. 158, p. 1227-1230.
24. Morte, A.; Díaz, G.; Rodríguez, P.; Alarcón, J. J. y Sánchez-Blanco, M. J. Growth and water relations of mycorrhizal and nonmycorrhizal Pinus halepensis plants in response to drought. Biologia Plantarum, 2001, vol. 44, no. 2, p. 263-267.
25. Augé, R. M. Water relations, drought and vesicular-arbuscular mycorrhizal simbiosis. Mycorrhiza, 2001, vol. 11, p. 3-42.
26. Muhsin, T. M. y Zwiazek, J. J. Ectomycorrhizas increase apoplastic water transport and root hydraulic conductivity in Ulmus Americana seedlings. New Phytol., 2002, vol. 153, p. 153-158.
27. Stone, P. J.; Wilson, D. R.; Reid, J. B. y Gillespie, R. N. Water defficit effects on sweet com. I. Water use, radiation use, efficiency, growth and yield. Australian Journal of Agricultural Research, 2004, vol. 52, no. 1, p. 103-113.
Recibido: 3 de diciembre de 2007
Aceptado: 15 de diciembre de 2008
