










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936
cultrop vol.32 no.3 La Habana jul.-set. 2011
Comportamiento de la variabilidad longitudinal del número de esporas de Glomus hoi-like inoculado en Brachiaria decumbens en dos periodos diferentes.
Behavior of longitudinal variability in the number of spores of Glomus hoi-like Brachiaria decumbens inoculated in two different periods
Bannie Vázquez, Ramón Rivera y Kalyanne Fernández
Instituto Nacional de Ciencias Agrícolas. Carretera de Tapaste Km 31/2, Gaveta Posta No.1, CP: 32700. San José de Las Lajas, La Habana, Cuba.
RESUMEN
Con el objetivo de determinar el comportamiento de la variabilidad longitudinal del número de esporas del hongo micorrízico arbuscular (HMA) Glomus hoi-like en canteros, utilizando Brachiaria decumbens como planta hospedera, se realizó este estudio. Para dar cumplimiento al mismo, se inoculó el cultivo con la cepa Glomus hoi-like (INCAM-4), suministrado por el cepario de HMA del INCA, sobre un sustrato arcilloso. Para evaluar el comportamiento del número de esporas en ambos periodos, se realizaron muestreos a los 60, 90 y 120 días de germinadas las semillas, donde se tomaron tres muestras compuestas en la profundidad de 0-20 cm, en cada cantero. Para determinar la variabilidad espacial del número de esporas en los canteros se tomó una muestra compuesta por tres submuestras cada 1,5 m, a lo largo del cantero. A cada muestra se le determinó el número de esporas y los contenidos de algunos indicadores químicos del sustrato. Se realizó una correlación entre el número de esporas y los indicadores químicos del sustrato. El número de esporas en ambos periodos fue aumentando con el tiempo y presentó una elevada variabilidad a lo largo del cantero. La mayor producción de esporas se presentó en el periodo de junio a octubre. La correlación entre la cantidad de esporas y algunos indicadores químicos revelaron resultados poco significativos y con muy bajos coeficientes de correlación, lo cual demostró que estos indicadores no son los responsables de la variabilidad de la producción de esporas.
Palabras claves: variabilidad, producción de esporas, hongos micorrízicos arbusculares.
ABSTRACT
In order to determine the behavior of longitudinal variability in the number of spores of arbuscular mycorrhizal fungi (AMF) Glomus hoi-like in beds, using Brachiaria decumbens as a trap plant, a study was performed. To comply with it, the crop was inoculated with Glomus hoi-like (INCAM-4), provided by the AMF strain collection of INCA, on a clay substrate. Samplings were conducted at 60, 90 and 120 days after seed germination in which the beds were subdivided into three equal parts and took a sample of five sub-samples in the depth of 0-20 cm, determined. the number of spores. To determine the spatial variability in the number of spores in a sample beds composed of three sub-samples every 1.5 m along the plant, each sample was tested for the number of spores and the contents of some chemical indicators substrate. A correlation between the number of spores and chemical indicators of the substrate were mode. The number of spores in both periods was increasing with time and showed a high variability throughout the bed. The highest production of spores were found in the period from June to October. The correlation between the amount of spores and chemical indicators showed poor results with very low correlation coefficients, which showed that these indicators are not responsible for the variability the spore production.
Keywords: variability, fungal spores production, arbuscular mycorrhizae.
INTRODUCCIÓN
Los hongos micorrízicos arbusculares (HMA) son microorganismos simbiontes pertenecientes al phylum Glomeromycota (1) que establecen una asociación mutualista y benéfica con las plantas. Es la simbiosis más extendida sobre el planeta, no sólo por su distribución, sino porque se establece con más del 90% de las plantas terrestres (2). Su distribución está influenciada por aspectos ecológicos relacionados con el clima y las características del suelo (3).
Los HMA contribuyen de forma eficiente a la supervivencia y crecimiento de las plantas micorrizadas, al reducir los estreses asociados con la nutrición, las relaciones con el agua, la estructura del suelo, el pH, las sales, los metales tóxicos y los patógenos (4). Por los beneficios que le brindan estos hongos a la mayoría de los cultivos y porque conservan el medioambiente, son muy importantes al actuar como biofertilizantes (5).
Entre los propágulos que contienen los biofertilizantes micorrízicos se encuentran las esporas, las hifas y las raíces micorrizadas. Las comunidades de HMA presentan diferencias en la producción de esporas y funciones correlacionadas con el ambiente (6).
Existen grandes variaciones en las estimaciones resultantes de algunos estudios enfocados a determinar el número de esporas de HMA en el suelo de ecosistemas áridos y semiáridos (7). Al respecto, se sugiere que estas diferencias numéricas pueden estar ligadas con factores ambientales y edáficos; así como a factores estacionales de la esporulación de estos hongos en relación con la especie vegetal hospedera.
De ahí la importancia de continuar realizando estudios sobre el ciclo de vida de los HMA a través de la dinámica poblacional de los mismos, principalmente de sus esporas. El objetivo de este estudio fue el de evaluar el comportamiento de la variabilidad longitudinal del número de esporas de estos hongos en dos periodos diferentes.
MATERIALES Y MÉTODOS
Los estudios referidos al presente trabajo se realizaron en el Instituto Nacional de Ciencias Agrícolas (INCA), el cual se encuentra ubicado en el Km 3½ de la carretera a Tapaste, Municipio San José de Las Lajas, Provincia Mayabeque.
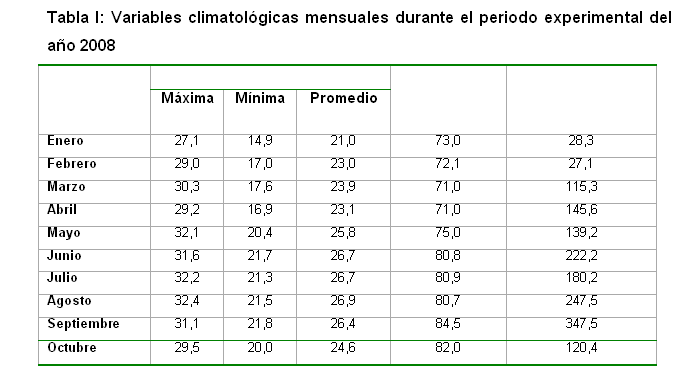
El mismo se ejecutó durante los periodos de enero a mayo y de junio a octubre del año 2008, con el objetivo de evaluar el comportamiento de la variabilidad longitudinal del número de esporas en dos épocas del año. Se recopilaron los valores de temperatura máxima, mínima y promedio, la humedad relativa promedio y las precipitaciones medias mensuales correspondientes al periodo de enero-octubre del 2008 (Tabla I). Tomando como referencia la Estación Meteorológica número 78374 ubicada en el Km 3½ de la carretera a Tapaste, municipio San José de Las Lajas, provincia Mayabeque.
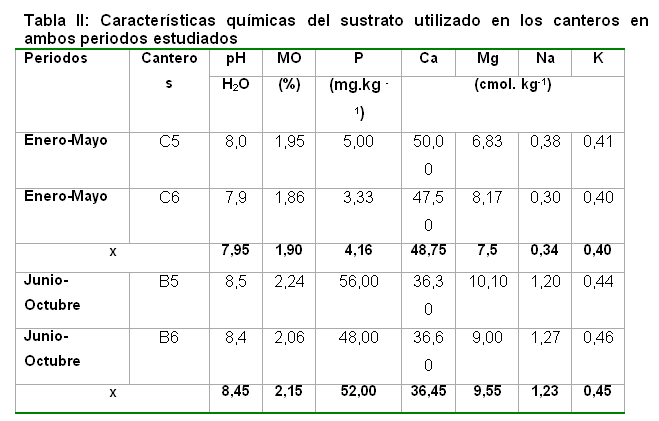
Se utilizaron canteros de polietileno negro con 15,0 m de largo x 1,0 m de ancho y una altura de 0,20 m, identificados como C5 y C6 en el primer periodo de siembra y como B5 y B6 en el segundo. En la base de los canteros se realizaron perforaciones para facilitar el drenaje del agua. Los canteros se llenaron con un sustrato natural arcilloso, cuyas características se presentan en la Tabla II.
El pH en H2O se determinó por potenciometría, utilizando una relación suelo- disolución 1:2,5. El contenido de materia orgánica (% M.O) se determinó siguiendo la metodología descrita por Walkley y Black y el fósforo (P2O5) por Oniani. Los cationes intercambiables se determinaron por una extracción con NH4AC 1 mol.L-1 a pH 7; calcio (Ca) y magnesio (Mg) por Complexometría y potasio (K) por Fotometría de llama (8).
Se empleó un inóculo de HMA que contenía 80 esporas por g-1 de la cepa Glomus hoi-like (INCAM-4), suministrado por el cepario de HMA del INCA.
El sustrato se esterilizó con una solución de formol ( 2,5%) aplicada a razón de 5 L.m-2, el cantero se cubrió cuidadosamente con polietileno negro por 72 horas para que accionara la solución aplicada, posteriormente se destapó dejándolo reposar por 72 horas hasta expulsar los gases.
La siembra se realizó a chorrillo en la primera semana del mes de enero y en la segunda semana del mes de junio, utilizando semillas de Brachiaria decumbens sembradas a una densidad de 13 g.m-2 y distribuidas en 7 surcos por cantero. Las semillas fueron recubiertas con inóculo micorrízico antes de la siembra mediante la tecnología descrita por Fernández y colaboradores (9). El inóculo sólido se aplicó a razón del 10 % del peso de la semilla.
En el momento de la siembra se aplicaron 150 g de la fórmula completa NPK (9 – 13 – 17) en cada cantero y a los 30 días de germinadas las semillas, 75 g de Urea (46 – 0 – 0), ambas aplicaciones se realizaron por cantero. Se aplicó riego manteniendo la humedad del sustrato en un límite cercano a la capacidad de campo.
Para evaluar el comportamiento del número de esporas en ambos periodos, se realizaron muestreos a los 60, 90 y 120 días de germinadas las semillas, donde se tomaron tres muestras compuestas por 5 submuestras en la profundidad de 0-20 cm en cada cantero.
Para la toma de muestras se utilizó una barrena de 6,0 cm de diámetro y 10,0 cm de profundidad. Las esporas se aislaron por el método de Tamizado húmedo y decantado (10, 11) y se contaron con ayuda de una placa de conteo de nemátodos, en un microscopio estéreo modelo Stemi 2000-C (40x).
Con el objetivo de determinar la variabilidad espacial del número de esporas de HMA en los canteros C5, C6, B5 y B6, se procedió a tomar una muestra compuesta por tres submuestras cada 1,5 m, para un total de 10 muestras compuestas por 3 submuestras, a lo largo del cantero. Las muestras compuestas se tomaron en los surcos dos, cuatro y seis y a cada una se le determinó el número de esporas.g-1 de sustrato y los contenidos químicos del mismo, para determinar la posible influencia de los contenidos de químicos sobre la variabilidad del número de esporas en el cantero.
Análisis estadístico
La variabilidad del Número de esporas.g-1 de sustrato a lo largo del cantero, se determinó mediante el establecimiento del porcentaje de variación de cada muestra con el valor promedio, así como el coeficiente de variación obtenido en cada uno de los canteros.
Para evaluar el efecto de los periodos de siembra sobre el número de esporas se compararon las medias con sus intervalos de confianza al 95%, mediante una prueba de Tukey. Se correlacionó, además, la variabilidad existente en la producción de esporas con la variabilidad en los contenidos químicos del sustrato.
Todos los análisis se realizaron utilizando el programa estadístico SPSS 11.5 para Windows.
RESULTADOS Y DISCUSIÓN
En ambos periodos (Figura 1A y B) el número de esporas obtenido mostró un incremento en el tiempo, lo cual se encuentra asociado con el propio desarrollo de la asociación micorrízica y con el completamiento del ciclo de vida del hongo.
En la figura 1A se muestra la dinámica del número de esporas del periodo de crecimiento enero-mayo, este indicador se incrementó lentamente hasta los 60 días con la producción de nuevas esporas. Posteriormente se refleja otro incremento significativo a los 90 días y a los 120 días se alcanzan los mayores valores con 39 esporas.g-1 de sustrato, los cuales mostraron diferencias altamente significativas con el resto de los momentos de muestreo.
En el periodo de junio-octubre (Figura 1B) se encontró, de forma similar, un incremento entre los diferentes momentos de muestreo, pero en este caso se alcanzaron diferencias significativas entre los tres momentos. Los valores alcanzados a los 120 días fueron de 108 esporas.g-1 de sustrato.
Teniendo en cuenta los intervalos de confianza al comparar los dos periodos de muestreo estudiados se encontró que los valores de número de esporas fueron estadísticamente superiores en el periodo junio-octubre con relación a enero-mayo.
Camargo y Esperón (12) también afirman que el número total de esporas de HMA en el suelo tiende a ser mayor durante la estación de lluvias y menor durante la estación de seca. Resultados similares encontraron González y colaboradores (13) en plantaciones de Brachiaria inoculadas con cepas eficientes de HMA.
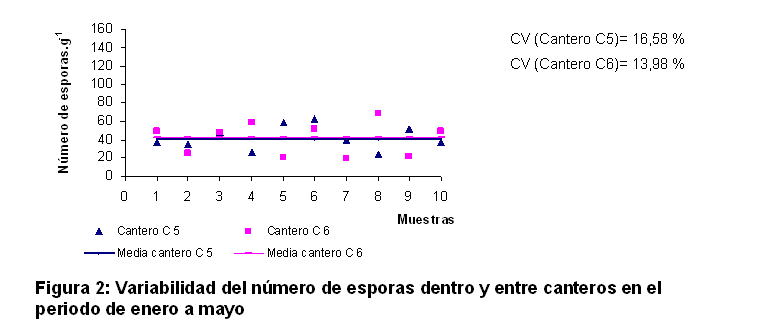
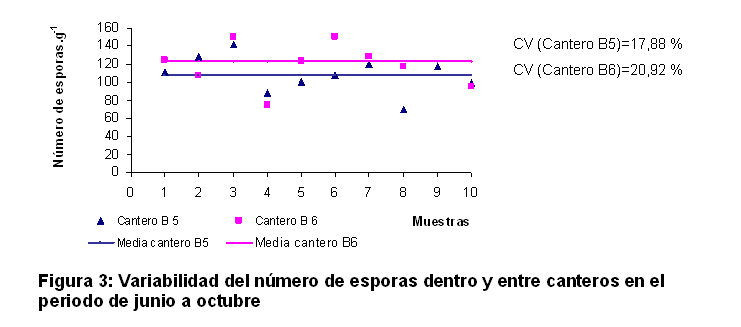
En las figuras 2 y 3 se presenta una estimación de la variabilidad del número de esporas.g-1 en función de la longitud del cantero. Esta estimación responde a la heterogeneidad de este indicador dentro del cantero y la comparación entre las medias de cada uno es una medida de la variación entre ellos.
En ambas figuras se puede observar una elevada variabilidad del número de esporas dentro de los canteros, presentándose un patrón muy similar entre los dos canteros sembrados en cada momento. Se observó que los valores individuales se encontraron en un rango aproximado al ±50% del valor promedio para cada cantero, conducta que se produjo en cada periodo de crecimiento.
En el periodo de enero a mayo se alcanzaron valores de 42 esporas.g-1 en el cantero C5 y 41 esporas.g-1 en el cantero C6, para 1% de variación con relación al valor promedio de ambos. En el periodo de junio a octubre el cantero B5 mostró un valor medio de 108 esporas.g-1 y B6 reveló 123 esporas.g-1, para un 6% de variación con relación al valor promedio de ambos, lo cual indicó una alta reproducibilidad de los resultados en los dos canteros evaluados en cada periodo.
La variabilidad del contenido de esporas de HMA dentro del cantero resultó ser mayor que entre ellos, esto ocurre debido a los patrones de esporulación, los cuales pueden variar a causa de diferentes factores como el movimiento físico y la saturación de agua (14), el pH (15) y la temperatura (16).
La variabilidad encontrada es una característica de la variable, dada por su complejidad biológica.
Las fluctuaciones en el número de esporas de HMA podría relacionarse con factores ambientales, estacionales y del suelo que afectan el proceso de esporulación del hongo, además del ciclo de vida de la planta huésped y de la fisiología propia de la simbiosis (17), todos estos factores pueden influir sobre la variación del número de esporas.
Teniendo en cuenta que los factores del suelo que actúan tanto en la formación como en el funcionamiento y establecimiento de la simbiosis son: disponibilidad de nutrientes, reacción del suelo, concentración de elementos metabólicos, humedad, aireación y textura del suelo, microbiota y fauna, temperatura y sistema de manejo (18). Se obtuvo como resultado en los Análisis de Correlación realizados entre la cantidad de esporas y algunos indicadores químicos revelaron resultados poco significativos (Tabla III) y con muy bajos coeficientes de correlación. La correlación obtenida con el Mg intercambiable se encontró en condiciones de mediana homogenidad de este nutriente en el cantero (Tabla III); ya que no fue el indicador que mayor coeficiente de variación mostró, poniendo en duda cualquier interpretación relacionada con la influencia de la variabilidad de los indicadores químicos sobre la heterogeneidad encontrada en la producción de esporas. Se mostró que en este caso, no parece ser la variación en los indicadores químicos la causa de la variabilidad en el número de esporas dentro del cantero.
Una vez establecida la variabilidad longitudinal dentro de los canteros como una conducta generalizada, al encontrarse en los 4 canteros muestreados (B5, B6, C5 y C6) en diferentes momentos, siguiendo un comportamiento similar en cualquiera de ellos, y al no encontrarse relaciones de dependencia entre ésta y la variabilidad de los indicadores químicos del sustrato, se considera que la variabilidad longitudinal del número de esporas debe ser una consecuencia de la variabilidad espacial de la esporulación como proceso biológico producto de la interacción con el medio a nivel de micrositio (19).
Los resultados de este trabajo, en estas condiciones no coinciden con los encontrados por Ettema y Wardle (20), quienes plantean que la heterogeneidad espacial presente en la composición del suelo puede ser otro factor que influye sobre la diversidad de los HMA. Teniendo en cuenta que sobre la producción de esporas influyen muchos factores, podemos concluir que en este caso las variaciones encontradas en los indicadores químicos del sustrato no fueron los causantes de la variabilidad del número de esporas lo que sugiere realizar otras investigaciones para determinar los factores que mayor influencia presentan sobre el indicador estudiado en nuestras concisiones.
REFERENCIAS
1. Schüßler, A.; Schwarzott, D. and Walker, C. A new fungal phylum,the Glomeromycota plylogeny and evolution. Mycologgical Researc, 2001, vol.105, no 12, p.1413-1421.
2. Wang, B.; Qiu Y. Phylogenetic distribution and evolution of mycorrhizas in land plants. Mycorrhiza, 2006, vol.16, no 5, p. 299-363.
3. Oehl, F.; Sieverding, E.; Ineichen, K.; Mäder, P.; Wiemken, A.; Boller, T. Distinct sporulation dynamics of arbuscular mycorrhizal fungal communities from different agroecosystems in long-term microcosms. Agriculture, Ecosystems and Environment , 2009, vol.134, no 3-4, p. 257-268.
4. Rai, M. K. Current advances in mycorrhization in micropropagation. In vitro Cell. Dev. Biol.-Plant, 2001, vol.37, no 2, p.158 - 167.
5. Smith, S.E y Read, D.J. Arbuscular mycorrhizas. Mycorrhizal Symbiosis. New York: Ed Academic., 2008, 800 p. ISBN-10: 0123705266.
6. Sanjuán, J. Introducción. Biofertilizantes en Iberoamérica: una visión técnica, científica y empresarial. Uruguay: Ed María Luisa Izaguirre-Mayoral, Carlos Labandera, Juan Sanjuan., 2007, 3-4 p.
7. Camargo-Ricalde, S.L y Dhillion, S.S. Endemic Mimosa species can serve as mycorrhizal "resource islands" within semiarid communities of the Tehuacán-Cuicatlán Valley, Mexico. Micorriza, 2003, vol. 13, no 3, p. 129-136.
8. Wolfe,E.; Mummey, D.; Rillig, M.; Klironomos, J . Small-scale spatial heterogeneity of arbuscular mycorrhizal fungal abundance and community composition in a wetland plant community. Mycorrhiza, 2007, vol. 17, no 3, p.175–183.
9. Paneque, V.M. La fertilización de los cultivos. Aspectos teórico-prácticos para su recomendación. (Folleto). La Habana: Ed Instituto Nacional de Ciencias Agrícolas., 2002, 28 p.
10. Fernández, F.; Rivera, R.; Noval, B. /y col./. Metodología de recubrimiento de la semilla con inóculo micorrizógeno. [Patente Cubana No 22641], 1999.
11. Gerdemann, J. W. y Nicholson, T. H. Spore of mycorrhizae endogone species extracted from soil by wet sieving and decanting. Trans Br. Mycol. Soc, 1963, vol. 46, no 2, p. 235 - 244.
12. Herrera, R. A.; Ferrer, R. L.; Furrazola, E. y Orozco, M.O. Estrategia de Funcionamiento de las Micorrizas VA en un Bosque Tropical. Biodiversidad en Iberoamérica: Ecosistemas, Evolución y Procesos sociales. Mérida: Ed Maximina Monasterio, 1995, 30 p.
13. Camargo-Ricalde, S.L. y Esperón-Rodríguez. Efecto de la heterogeneidad espacial y estacional del suelo sobre la abundancia de esporas de hongos micorrizógenos arbusculares en el valle semiárido de Tehuacán-Cuicatlán, México. Revista de Biología Tropical, 2005, vol. 53, no. 3-4. p 339-352.
14. González, P.J.; Plana, R.; Rivera R.; Fernández, F. y Arbola, J. Efectos de la inoculación de hongos micorrízicos arbusculares en pastos del género Brachiaria, cultivados en suelo Pardo Mullido. Revista Cubana de Ciencia Agrícola, 2008, vol. 42, no 1, p. 101.
15. van der Heijden, M.; Streitwolf-Engel, R.; Riedl, R.; Siegrist, S.; Neudecker, A.; Ineichen, K.; Boller, T., Wiemken, A.; Sanders, I.R. The mycorrhizal contribution to plant productivity, plant nutrition and soil structure in experimental grassland. New Phytologist, 2006, vol.172, p. 739-752.
16. IJdo, M., Cranenbrouck, S.; Declerck, S. Methods for large-scale production of AM fungi: past, present, and future. Mycorrhiza, 2011, vol. 21, no 1, p.1-16.
17. IJdo M. Mass production of arbuscular mycorrhizal fungal inoculum. [Thèse présentée en vue de l’obtention du grade de docteur en sciences agronomiques et ingénierie biologique] Université catholique de Louvain, 2010, 208 p.
18. Calderón, A. Estudio de sustratos y micorrizas arbusculares en la adaptación de vitroplantas de banano (Musa sp). [Tesis en opción al grado académico de maestro en ciencias en Nutrición de las plantas y Biofertilizantes] INCA, La Habana, 2004,105 p.
19. Whitcomb, S and Stutz, J. Assessing diversity of arbuscular mycorrhizal fungi in a local community: role of sampling effort and spatial heterogeneity. Mycorrhiza, 2007, vol.17, no 5, p. 429–437.
20. Ettema CH, Wardle DA. Spatial soil ecology. TRENDS in Ecology and Evolution, 2002, vol. 17, no 4, p.177–183.
Recibido 30/11/2010, aceptado 02/06/2011.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}