










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936
cultrop vol.32 no.4 La Habana oct.-dic. 2011
Evaluación de la variabilidad morfoagronómica de una colección cubana de maíz (Zea mays L.)
Evaluation of variability of a Cuban collection morphoagronomic corn (Zea mays L.)
M. Sc. Michel Martínez Cruz1, Dr. Rodobaldo Ortiz Pérez1, Dr. Humberto Rios Labrada1 y M. Sc. Rosa Acosta Roca1.
1 Instituto Nacional de Ciencias Agrícolas (INCA)
RESUMEN
En nuestro país, el maíz se cultiva desde la época de los aborígenes y constituye un alimento básico en la nutrición humana, del ganado y las aves; es el segundo cereal de importancia y tiene alta preferencia de consumo por la población. En Cuba, existen seis razas de maíz con una alta diversidad morfoagronómica, la cual ha sido ampliamente estudiada. El presente trabajo se realizó en una muestra de 106 accesiones, donde se incluyeron accesiones conservadas in situ en fincas de campesinos y conservadas ex situ. El objetivo del estudio fue evaluar el grado de variabilidad morfoagronómica de la colección; utilizando para ello 16 caracteres morfoagronómicos. Los resultados obtenidos permitieron detectar la existencia de variabilidad morfoagronómica en la muestra. Se demostró que la colección de maíz evaluada, presentó variabilidad morfoagronómica debido a la diferencia entre genotipos, a la pluralidad de orígenes, manejo y condiciones ambientales en que se desarrolla este cultivo en Cuba. Además, en la colección estudiada, la variabilidad de los caracteres evaluados es moderada, debido a la selección que realizan los campesinos sobre ellos. Los caracteres más variables fueron la incidencia de Spodoptera frugiperda (Smith), altura a la mazorca superior y peso de cien semillas.
Palabras clave: maíz, variabilidad morfoagronómica.
ABSTRACT
In our country, the corn is cultivated from the time of the aboriginal ones and it constitutes a basic food in the human nutrition, of the livestock and the birds; it is the second cereal of importance and has high consumption preference for the population. In Cuba, six races of corn exist with a high diversity, which has been broadly studied. The present work was carried out in a sample of 106 local varieties, these local varieties were included conserved in situ in peasants' properties and conserved ex situ. The objective of the study was to evaluate the degree of variability of the collection; using for it 16 characters. The obtained results allowed to detect the existence of variability in the sample. It was demonstrated that the evaluated collection of corn presented variability due to the difference among genotypes, to the plurality of origins, handling and environmental conditions in that this cultivation is developed in Cuba. Also, in the studied collection, the variability of the evaluated characters is moderate, due to the selection that the peasants carry out on them. The most variable characters were the incidence of Spodoptera frugiperda (Smith), height to the superior ear and weight of a hundred seeds.
Key Words: corn, morphoagronomic variability.
INTRODUCCIÓN
Entre las plantas cultivadas actualmente, el maíz, por su producción a nivel mundial, ha superado al trigo y al arroz (1); se cultivan aproximadamente 140 millones de hectáreas, siendo Estados Unidos, China y Brasil los principales productores (2).
En nuestro país el maíz se cultiva desde la época de los aborígenes y constituye un alimento básico en la alimentación humana, del ganado y las aves (3). En el 2006, en Cuba se cosecharon 122,160 miles de hectáreas (Mha) de maíz, de ellas, al sector estatal correspondieron 8,919 Mha y al no estatal 113,241 Mha; con una producción de 305,400 miles de toneladas (Mt) de maíz tierno, siendo en el sector estatal de 19,978 Mt y en el no estatal de 285,422 Mt. El rendimiento del maíz fue de 2, 50 toneladas por hectárea (t. ha-1) siendo en el sector estatal de 2,24 t. ha-1 y en el sector no estatal de 2,52 t. ha-1 (4).
Es la planta cultivada más estudiada por el hombre, la de mayor diversidad, no sólo genética, sino también de usos. El maíz ha evolucionado por selección natural, por la selección dirigida por los agricultores-mejoradores durante miles de años y por los mejoradores profesionales en los últimos 150 años (1). De acuerdo con lo planteado por Wilkes en 1988 (5), México es el centro primario de diversidad genética y la Zona Andina el secundario, donde el cultivo del maíz ha tenido una rápida evolución.
Desde el punto de vista biológico los procesos que generan y mantienen la diversidad genética del maíz son dinámicos: mutación, selección, recombinación, etc., y están ligados a las necesidades a largo plazo de las poblaciones humanas en crecimiento y a sus cada vez mayores expectativas de vida (6). La conservación de la diversidad debe estar basada en el mantenimiento de estos procesos, así como, en la preservación de la razas per se. Actualmente se observa una variación continua en la diversidad del maíz, sobre todo en sus caracteres cuantitativos; la mayoría de las poblaciones representan combinaciones de raza (7). En gran medida, la diversidad del maíz, se puede atribuir también a la selección practicada por el hombre desde su domesticación, así como a los numerosos nichos ecológicos y los efectos ambientales que cada condición climática ejerce sobre las poblaciones para determinar la adaptación de éstas (8).
La variabilidad morfoagronómica del maíz ha sido ampliamente estudiada en Cuba (9), no obstante, dado la condición alógama del maíz, es inevitable que se produzcan de manera natural hibridaciones o mezclas, ya que muchos agricultores mantienen en sus sistemas tradicionales más de una accesión, lo que provoca la creación de nueva diversidad con adaptación a condiciones específicas. Dada esta problemática se hacen necesarias las prospecciones y la evaluación periódica de la variabilidad generada.
Dada esta problemática el presente trabajo tuvo como objetivo: Caracterizar la variabilidad morfoagronómica de una colección cubana de maíz.
MATERIALES Y MÉTODOS
Caracterización morfoagronómica.
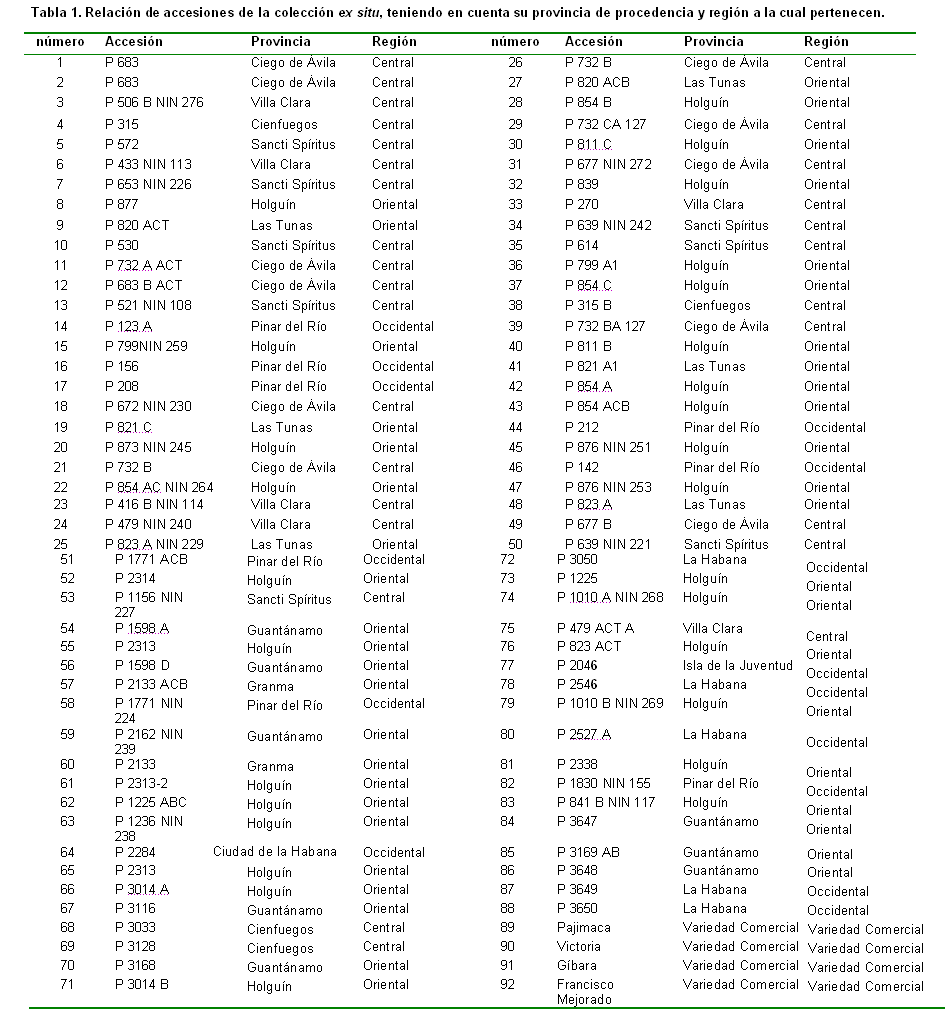
Para el desarrollo del presente trabajo se utilizó una colección de maíz integrada por 106 accesiones (Tabla I): 63 procedentes de la región occidental, de ellas, 19 provenientes del Banco de Germoplasma del Instituto de Investigaciones Fundamentales en Agricultura Tropical (INIFAT), 21 del Banco de Germoplasma del Instituto Nacional de Ciencias Agrícolas (INCA) colectadas por Acosta (2003) en La Palma, Pinar del Río, 21 accesiones colectadas en Catalina de Güines, una de San Antonio de los Baños (variedad FELO) y una de Batabanó (Línea Raúl); 16 accesiones del centro, de ellas, ocho colectadas en Santi Spíritus y ocho en Villa Clara y 27 de la región oriental, de ellas, 17 colectadas en Ventas de Casanova, Santiago de Cuba, seis en la Empresa de Semillas de Santiago de Cuba, tres en Holguín y una en Granma.
Durante los años 2003, 2004 y 2005 se realizaron tres ciclos de evaluación a la colección de maíz en el área central del INCA, ubicada en el municipio San José de las Lajas, provincia La Habana. En el año 2003, la siembra se realizó el 10 de junio; en el 2004, el 5 de febrero y en el 2005, el 11 de noviembre. Fue utilizada la misma área experimental para cada ciclo y el maíz fue parte de un sistema de rotación con frijol o soya. Los datos climáticos de los periodos de siembras de los tres años se muestran en el anexo 2.
Las siembras se realizaron en parcelas de 5,0 m de largo, a razón de seis surcos por parcela, a una distancia de 0,30 m x 0,70 m, con una población de 115 plantas por parcela sobre en un suelo Ferralítico rojo lixiviado (10).
Las atenciones culturales se realizaron según el Instructivo Técnico del cultivo (11), excepto que no se efectuaron aplicaciones de fertilizantes químicos. Se utilizó, en cada siembra, un diseño completamente aleatorizado con tres réplicas.
A las 106 accesiones, se les realizaron evaluaciones de 16 caracteres morfoagronómicos (Tabla II) en cada ciclo de siembra, tomando una muestra de 10 plantas por accesión, según el Manual Gráfico para la Descripción Varietal del Maíz (Zea mays L.) (12). Las evaluaciones se efectuaron, seleccionando, en cada parcela, las plantas del centro, para evitar el efecto de borde y minimizar la influencia de la polinización cruzada.
Análisis de la variabilidad de la colección estudiada.
Se determinaron los parámetros estadísticos Media (X) y coeficiente de variación (CV) de la colección general y de los grupos de accesiones para los caracteres número de granos por hileras, número de hileras, masa de 100 semillas, longitud de la mazorca, diámetro medio de la mazorca, número de granos por mazorca, diámetro del tallo, altura a la mazorca superior, longitud de la planta, incidencia de Spodoptera frugiperda (Smith) y rendimiento por planta. Para determinar estos parámetros se calculó la media de los tres años en cada carácter.
Se realizó un análisis de componentes principales, a los caracteres cuantitativos (DT, AMS, LP, NMP, NGH, NH, LM, DMM, NGM, ISF, M100S y R/P) de la colección general, utilizándose el factor accesión y se construyó un gráfico de dispersión, donde se ubicaron las 106 accesiones estudiadas, en base a las dos primeras funciones canónicas.
Se realizó un análisis discriminante, utilizando como variable discriminante la región de procedencia, basado en los caracteres cuantitativos de la colección general, con el objetivo de definir si existen diferencias entre los grupos de accesiones, pertenecientes a diferentes regiones de procedencia. Posteriormente se elaboró un gráfico de dispersión, donde se ubicaron las 106 accesiones estudiadas, en base a las dos primeras funciones discriminantes.
Todos los análisis estadísticos fueron ejecutados utilizando el paquete estadístico SPSS, Versión 11,5, (13).
RESULTADOS Y DISCUSIÓN
Análisis de la variabilidad de la colección estudiada.
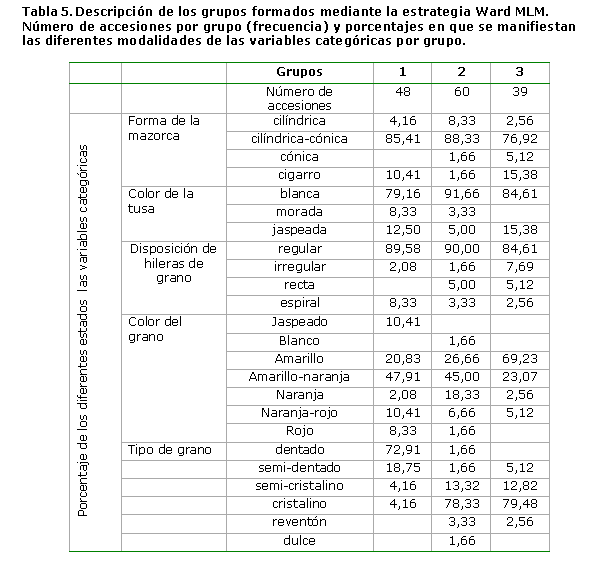
La tabla III presenta los valores de la media y coeficiente de variación para los caracteres número de granos por hileras, número de hileras, masa de 100 semillas, longitud de la mazorca, diámetro medio de la mazorca, número de granos por mazorca, diámetro del tallo, altura a la mazorca superior, longitud de la planta, Incidencia de Spodoptera frugiperda (Smith) y rendimiento por planta, tanto de la colección general como de las tres regiones (occidental, central y oriental).
La incidencia de Spodoptera frugiperda (Smith) fue el carácter de mayor dispersión, en la colección general, con un coeficiente de variación de 27,15 % y en las accesiones de la región occidental (29,17 %); siendo también variable en la región oriental (21,21 %).
El rendimiento por planta mostró una alta dispersión (CV > 25 %) en todas las colecciones, siendo, el carácter de mayor variabilidad, en las accesiones procedentes de las regiones oriental y central, con coeficientes de variación de 28,88 % y 25,64 % respectivamente. La altura a la mazorca superior mostro una alta variabilidad en la región central (20,40 %).
El carácter de menor dispersión, en todas las colecciones fue el número de mazorcas por planta, con coeficientes de variación inferiores a 5 % en todos los casos, debido a que la mayoría de las plantas presentaban una mazorca.
Los caracteres rendimiento por planta, altura a la mazorca superior, masa de 100 semillas, número de granos por mazorcas e incidencia de Spodoptera frugiperda (Smith), mostraron dispersión relativamente alta en todas las colecciones, con porcentajes del CV siempre superiores a 25 %, 16 %, 14 %, 12,5 % y 11 % respectivamente. Los resultados expuestos coinciden con Hatheway (1957) (14), quien informó que los maíces cubanos tienen su mayor variación para los caracteres de la mazorca y que por tanto, constituyen los más importantes para clasificar el germoplasma; sin embargo, la altura a la mazorca superior, es un carácter de la planta, el cual mostró variabilidad en las colecciones evaluadas, por tanto se recomienda evaluar su importancia, en la clasificación del germoplasma cubano.
Ortiz y Sevilla (1997) (15), al evaluar accesiones de maíz en Perú, obtuvieron valores inferiores a los antes citados para los caracteres de la mazorca, en contraste, Ligarreto, Ballén y Huertas (1998), al evaluar 25 accesiones de maíz de la zona andina, encontraron una variabilidad superior para los caracteres número de granos por hileras (22,40 %), número de hileras (14,27 %), masa de 100 semillas (24,62 %), longitud de la mazorca (15,53 %), diámetro medio de la mazorca (12,12 %), diámetro del tallo (13,35 %) y longitud de la planta (12,40 %). Las colecciones evaluadas en Cuba, presentan menor variabilidad que las evaluadas en la región andina; debido a que esta región es definida por Muñoz (2003) (1) como centro secundario de diversificación del maíz y por tanto la variabilidad de los caracteres es alta.
González y col. (2008) (16), al analizar variedades comerciales e híbridos de maíz en el Valle Toluca-Atlacomulco en México, encontraron los coeficientes de variación siguientes: para el número de hileras 4,9 %, para la masa de 100 semillas 10,0 %, para la longitud de la mazorca 4,87 %, para la altura a la mazorca es superior a 7,78 % y para la longitud de la planta 5,25 %.
La dispersión encontrada por estos autores, en los caracteres antes mencionados, fue inferior que en las poblaciones evaluadas en este trabajo, aunque las diferencias no fueron grandes, lo cual se debe a que las variedades evaluadas por dichos autores, son híbridos y variedades comerciales, las cuales, de manera general, presentan mayor homogeneidad.
Espinosa, Mendoza y Castillo (2006) (17), al evaluar poblaciones locales de maíz en México, informaron que la longitud de la mazorca tuvo un CV de 13,7 %, el diámetro medio de la mazorca de 8,7 %, el número de hileras de 8,7 %, el número de granos por hileras de 12,7 % y la masa de 100 semillas de 12,0 %. La dispersión de los caracteres número de granos por hileras y número de hileras fue similar a la encontrada en las poblaciones cubanas; mientras que la variabilidad de la masa de 100 semillas fue inferior, siendo superior en la Longitud de la mazorca y diámetro medio del tallo en las poblaciones locales de México.
Fernández (2009) (9), al evaluar accesiones cubanas de maíz conservadas tanto ex situ como in situ, informó que los caracteres de mayor variabilidad fueron M100S, DT y NGH con coeficientes de variación de 22,48%, 18,44% y 15,57% respectivamente, siendo el carácter menos variable el NH con CV de 8,22%.
En general, la variabilidad de los caracteres, se consideró de moderada a moderadamente alta, lo cual se debió, principalmente, a la interacción genotipo – ambiente, ya a que, las evaluaciones se realizaron en tres años diferentes y en tres momentos de siembra diferentes, conjuntamente, la pluralidad de orígenes de las accesiones que conformaron estas colecciones, provocaron un comportamiento diferenciado de las mismas. Los caracteres menos estables (ISF, AMS y M100S), fueron fuertemente influenciados por el ambiente (9), siendo esta es la causa principal de su variabilidad.
La tabla IV muestra la matriz de funciones canónicas conformada a partir del análisis de componentes principales de los caracteres cuantitativos evaluados en la colección general.
Las dos primeras funciones canónicas resultantes del análisis de componentes principales explicaron el 71,87 % de la variabilidad total, siendo los caracteres R/P, NGM, LM, M100S, NH y NGH los que más aportaron a la conformación de la función I. En la función II los caracteres que más aportaron fueron DT, LP e –ISF.
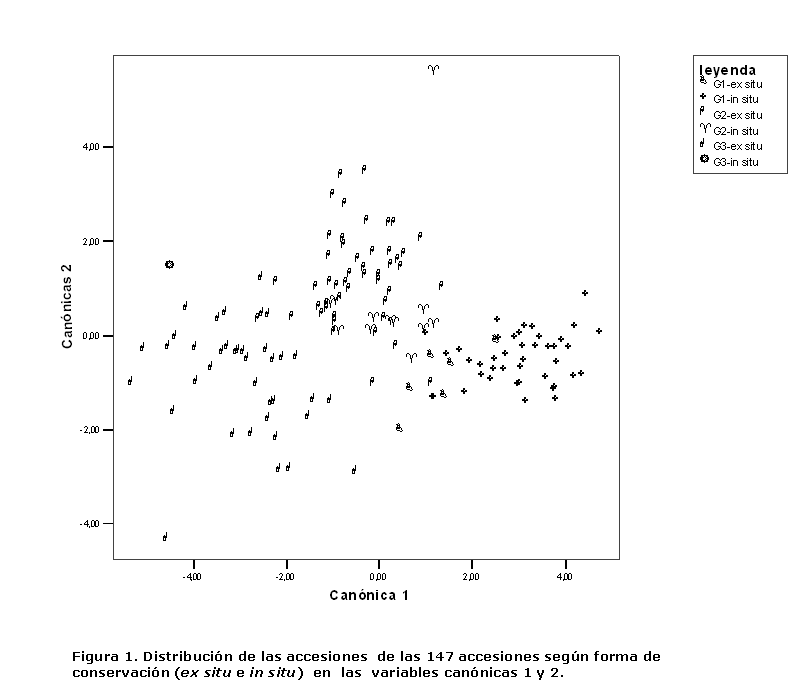
La figura 1 muestra la distribución gráfica de las 106 accesiones evaluadas de acuerdo a las dos primeras funciones canónicas; mostrándose una alta dispersión de las 106 accesiones evaluadas, por tanto, existió una amplia variabilidad, la que se debió principalmente, a la multiplicidad de orígenes de las accesiones que integran la colección de trabajo, donde existen materiales diversos (accesiones criollas, variedades mejoradas, líneas, etc.), introducidos por diferentes vías y obtenidos por programas de mejoramiento destinados a objetivos distintos.
Las hibridaciones o mezclas que manera natural ocurren en el maíz, al constituir éste una especia alógama, constituye otro aspecto que aportó a la variabilidad del cultivo. También influyeron la selección realizada por los agricultores y la gran diversidad de usos y de ambientes en que se siembra, provocando que, poblaciones fenotípicamente parecidas puedan pertenecer ahora a razas distintas, ya que no pueden compartir por falta de adaptación, el mismo ambiente ecológico.
NGH=número de granos por hileras, NH=número de hileras, M100S=masa de 100 semillas, LM=longitud de la mazorca, DMM=diámetro medio de la mazorca, NGM=número de granos por mazorca, DT=diámetro del tallo, LP=longitud de la planta, ISF=incidencia de Spodoptera frugiperda (Smith) y R/P=Rendimiento por planta.
Defacio y col. (2005) (18) encontraron variabilidad en los caracteres diámetro medio de la mazorca, número de granos por hileras y número de hileras al evaluar 60 accesiones de maíz nativas de Argentina.
De igual manera, Grijalva y col. (2003) (19), encontraron una amplia dispersión en los caracteres días a la floración masculina, longitud de la planta, altura de la mazorca superior y masa de 100 semillas, al evaluar seis líneas endogámicas de maíz y sus correspondientes híbridos en Saltillo, México.
Según Alfaro y Segovia (2000) (20), la evaluación de una determinada accesión colectada en un ambiente particular para posteriormente ser evaluada en un ambiente diferente, pudiera afectar el comportamiento de dicha accesión; por lo que la evaluación de la diversidad de las accesiones colectadas en diferentes zonas de Cuba, estuvo basada en los datos obtenidos en el lugar de evaluación (INCA) y no necesariamente representa su comportamiento en el lugar de origen.
Según Grant y col. (1965) (21) y Sevilla (1991) (22), los estudios de las poblaciones locales de maíz son indispensables si se quiere ampliar la base genética del cultivo para el mejoramiento con vista a la obtención de variedades con adaptación a condiciones específicas.
Estos resultados coinciden con Fontana y González (2000) (23), quienes plantearon que la variabilidad genética presente dentro y entre poblaciones de maíz ha sido reconocida como una de las más abundantes del reino vegetal. Mucha de esta variabilidad es de herencia simple, identificada a través del tiempo y que controla caracteres fenotípicos de fácil identificación, tales como, colores, formas y estructuras.
Goodman y Paterniani (1969) (24) al evaluar 111 caracteres de 55 razas y sub-razas de maíz del sureste de América del sur, en Piracicaba, Brasil, señalaron que muchos de los caracteres fueron afectados por el ambiente, siendo particularmente importante la interacción genotipo-ambiente de algunos caracteres relacionados con la arquitectura de la planta.
El trabajo reveló que los caracteres vegetativos tuvieron diferencias entre las razas evaluadas en diferentes ambientes, años y localidades. Algunas característica de la mazorca fueron más discriminativas que las características vegetativas, estas relaciones podrían indicar que, en general, los caracteres reproductivos fueron más estables que los vegetativos.
En este sentido, los mejoradores de maíz han expresado que en esta planta se encuentra toda la variabilidad genética necesaria para mejorar cualquier característica que se desee, pero en realidad, los programas de mejoramiento han hecho un uso limitado de la diversidad de los recursos genéticos del maíz disponibles fuera de los bancos de germoplasma de los institutos de investigación, mostrándose una tendencia general a usar el mismo germoplasma en los trabajos experimentales (25).
La principal fuente de variabilidad genética usada en los trabajos con maíz proviene, en general, de los materiales de que dispone el mejorador, o de un intercambio de germoplasma con otros mejoradores; sin embargo, hay un creciente interés en los mejoradores de maíz, de expandir la búsqueda de genes útiles, que aumenten la variabilidad genética y de este modo incrementar la sostenibilidad de la producción de dicho cultivo. De la misma forma, hay una constante preocupación sobre de la necesidad de conservar los recursos genéticos para su uso en este siglo y en el futuro más lejano.
El fitomejoramiento plantea una paradoja, ya que por un lado, se necesita variabilidad genética para mejorar y mantener alta productividad de los cultivos y por el otro, algunos cultivares mejorados se difunden ampliamente y su extenso uso provoca una reducción de variabilidad genética en las zonas de producción donde tienen éxito. El empleo de pocas variedades uniformes y con estrecha base genética en grandes áreas, ha ocasionado graves problemas en diferentes países y en distintas épocas. Se considera que, en especies cultivadas, el factor de mayor importancia en la pérdida de diversidad es el reemplazo de las variedades tradicionales y razas locales de los agricultores con amplia variabilidad genética, por materiales mejorados de mayor rendimiento pero con gran uniformidad genética, o por otros cultivos más productivos y/o más rentables.
En general, Las condiciones específicas de cultivo en cada finca de los agricultores, así como los diferentes criterios de selección, necesidades e intereses diferentes, contribuyó positivamente a mantener y ampliar la variabilidad útil en el maíz en las zonas estudiadas, por tanto, es importante el uso de esta variabilidad encontrada, tanto en programas de mejoramiento, como directamente en la producción, de forma tal que permita, tanto la conservación de este acervo genético como su mayor distribución y uso.
Distribución de las accesiones, en las diferentes zonas de procedencia, de acuerdo al análisis discriminante.
La tabla V muestra la matriz de funciones discriminantes conformada a partir del análisis de los caracteres cuantitativos evaluados en la colección general que incluye las 106 accesiones estudiadas, utilizando como factor discriminante la región de procedencia de las accesiones.
Las dos primeras funciones discriminantes explicaron el 100 % de la variabilidad, siendo los caracteres: número de granos por mazorcas, número de granos por hileras, longitud de la mazorca, rendimiento por planta, e incidencia de Spodoptera frugiperda (Smith), con aporte positivo todos, los de mayor aporte a la explicación de la varianza en la Función I.
Los caracteres, número de hileras, diámetro del tallo, diámetro medio de la mazorca, masa de 100 semillas y longitud de la planta, con aporte negativo determinaron la conformación de la función II. El análisis dio como resultado diferencias significativas entre las dos funciones discriminantes.
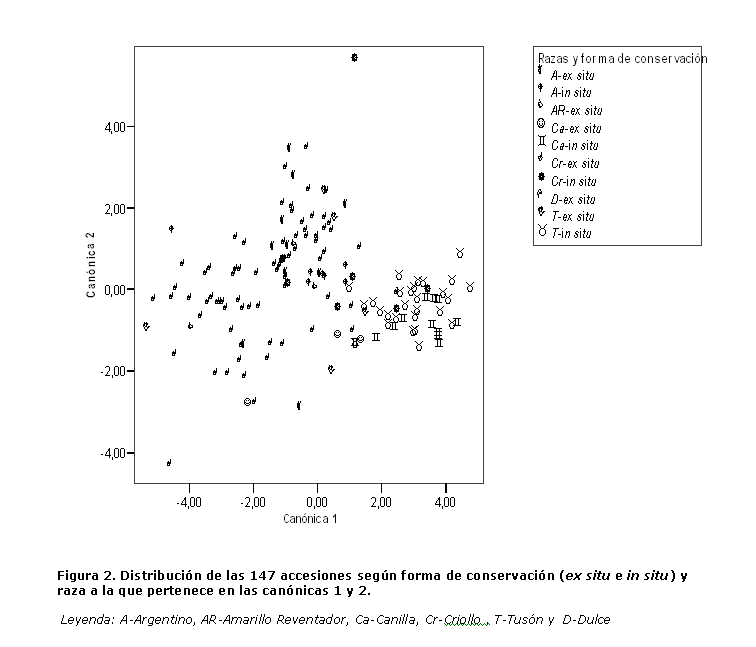
A continuación en la figura 2 se muestra la ubicación gráfica de las 106 accesiones evaluadas en las funciones discriminantes I y II, en base a los caracteres que tuvieron mayor contribución a la formación de las mismas.
NGM=número de granos por mazorca, NGH=número de granos por hileras, LM=longitud de la mazorca, R/P=Rendimiento por planta, ISF=incidencia de Spodoptera frugiperda (Smith), NH=número de hileras, DT=diámetro del tallo, DMM=diámetro medio de la mazorca, M100S=masa de 100 semillas y LP=longitud de la planta.
Las accesiones se agruparon atendiendo a las regiones de procedencia, formándose 3 grupos; un primer grupo (G-I) conformado por 61 de las 63 procedentes de la región occidental, quedando fuera de este, las accesiones 300 (variedad FELO) y 219 (línea Raúl) debido a que en el proceso de obtención de estas variedades fueron utilizadas accesiones de la región central como fuente principal de genes. El G-I se caracterizó por valores bajos de los caracteres en la función I y medios de los caracteres en la función II.
El grupo II (G-II) estuvo integrado por las accesiones de la región central más las accesiones 300, 219 (región occidental) y la 134 proveniente de la región oriental; se caracterizó por valores altos de los caracteres en la función I y bajos de los caracteres en la función II.
El grupo III (G-III) estuvo conformado por 25 de las 26 accesiones de la región oriental; se caracterizó por valores altos de todos los caracteres. Este grupo constituye una importante fuente de genes para el mejoramiento genético, debido a que, tiene las accesiones de mayores rendimientos por planta, aunque presentó altos índices de afectación por Spodoptera frugiperda (Smith), en las condiciones y el ambiente en que fueron evaluadas. En este caso se recomienda evaluar el comportamiento, ante dicha plaga, de estas accesiones en su lugar de origen, donde, según los campesinos, tienen tolerancia a la incidencia de la plaga clave del maíz.
Goodman y Bird (1977) (24), estudiaron la variabilidad entre 219 razas y subrazas de maíz de Latinoamérica, utilizando la taxonomía numérica sobre 20 caracteres morfoagronómicos y datos geográficos incluidos en catálogos de razas de maíz. Cuando consideraron todos los caracteres se formaron 14 grupos; al observar sólo ocho caracteres del grano (CG) y caracteres de mazorca (CM) encontraron que las razas, maíz reventador y el complejo Cuzco, quedaron bien separados de todas las otras razas, estas últimas formaron 19 grupos con poca sobreposición entre ellos.
Álvarez y Lasa (1990) (26), encontraron resultados similares en 36 poblaciones de maíz colectadas al norte de España, evaluando 13 componentes del rendimiento. De esta evaluación se formaron 10 grupos diferentes; los análisis discriminantes revelaron que los caracteres del grano y caracteres de la mazorca constituyeron los de mayor importancia en esa clasificación, donde el 72,4% de la variación fue explicada por las dos primeras funciones canónicas. Llaurado y Moreno (1993) (27), encontraron en maíces del noroeste de España que los caracteres con un mayor poder taxonómico o clasificatorio estaban relacionados con la precocidad y la altura de la planta, seguidos en orden de importancia por el cerrado de la mazorca y el color de los granos.
De las 106 accesiones evaluadas, 103 (97,2 %) se ubicaron en los grupos predefinidos inicialmente por la zona de procedencia demostrándose que las accesiones se agrupan según la zona de procedencia, basado en caracteres específicos que las identifican. Las accesiones provenientes de la región oriental y central, caracterizadas por valores altos de los caracteres de la función I, constituyen una fuente genética importante para ser utilizada en programas de mejoramiento.
Conclusiones
1. La colección de maíz evaluada, presentó variabilidad morfoagronómica; debido a la diferencia entre genotipos, a la pluralidad de orígenes, manejo y condiciones ambientales en que se desarrolla este cultivo en Cuba.
2. En la colección estudiada, la variabilidad de los caracteres evaluados es moderada, debido a la selección que realizan los campesinos sobre ellos. Los caracteres más variables fueron la incidencia de Spodoptera frugiperda (Smith), altura a la mazorca superior y peso de cien semillas.
REFERENCIAS
1. Muñoz, A. (2003). Prehistoria e Historia, Diversidad, Potencial, origen genético y Geográfico.Texcoco, México D.F.: CENTLI-MAÍZ.
2. Taba, S., van Ginkel, M., Hoisington, D. and Wellhausen-Anderson, D. Plant Genetic Resources Center: Operations manual 2004. El Batan, Mexico: CIMMYT, 2004
3. Socorro, M. y Martin, D. Editorial Pueblo y Educación. 318 pp., 1989
4. Oficina Nacional de Estadísticas. (2008. Edición 2009). Anuario Estadístico de Cuba.
5. Wilkes, G. (1988). Teosinte and the Other Wild Relatives of Maize. Mexico.
6. Wilkes, H.G. 1979. Mexico and Central America as a centre for the origin of agriculture and the evolution of maize. Crop Improv. 6(1):1–18.
7. Ramos, A. y E. Hernández X. 1972.Variaciónmorfológica de los maíces de la zona oriental del estado de México y Centro de Puebla, México. En: Xolocotzia. Tomo II. 1987. Universidad Autónoma Chapingo. Subdirección de Centros Regionales. Dirección de Difusión Cultural. Revista de Geografía Agrícola. México.
8. Carballoso, V., Mejía, A., Valderrama, S., Carballo, A., y González, F. (2000). Divergencia en poblaciones de maíz nativas de valles Altos de México. Agrociencia. 34 (2), 167-174.
9. Fernández, L. (2009). Identificación de razas de maíz (zea mays L.) presentes en el germoplasma cubano. La Habana: Instituto de Investigaciones Fundamentales en Agricultura Tropical. INIFAT.
10. Hernández, A., Morell, F., Ascanio, M. O., Borges, Y., Morales, M., y Yong, A. (2006). Cambios globales de los suelos ferralíticos rojos lixiviados (Nitisoles Ródicos Eútricos) de la provincia La Habana. Cultivos Tropicales, 27 (2), 41-5.
11. MINAGRI. (1992). Instructivo Técnico del cultivo del maíz. La Habana: Dirección Nacional de Cultivos Varios.
12. Carballo, C. A., y Benítez, V. A. (2003). Manual Gráfico para la descripción varietal del maíz (Zea mays L.). México, D. F.: Secretaria de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación (SAGARPA)-Servicio Nacional de Inspección y Certificación de Semilla (SNICS)-Colegio de Postgraduados en Ciencias Agrícolas (CP).
13. SPSS Tablas TM 11.5. (2002). ISBN 1-56827-923-X.
14. Hatheway, W. (1957). Races of Maize in Cuba. Washington, D.C.: National Academy Of Siences-National Research Council.
15. Ortiz, R., y Sevilla, R. (1997). Quantitative descriptors for classification and characterization of highland peruvion maize. Plant Genetic Resources Newsletter, 110, 49-52.
16. González, A., Vázquez, L. M., Sahún, J., y Rodríguez, J. E. (2008). Diversidad fenotípica de variedades e híbridos de maíz en el Valle Toluca-Atlacomulco, México. Fitotec. Mex., 31 (1), 67-76.
17. Espinosa, E., Mendoza, M., y Castillo, F. (2006). Diversidad fenotípica entre poblaciones de maíz con diferentes grados de pigmentación. Fitotec. Mex., 29 (Número Especial 2), 19-23.
18. Defacio, R. A., Hourquescos, M. J., Bramardi, S., y Ferrer, M. E. (2005). Estudio de variabilidad en poblaciones nativas de maíz.
19. Grijalva, O., Rincón, F., Gutiérrez del Río, E., Ruiz, N. A., y Bustamante, L. (2003). Componentes genéticos de caracteres agronómicos y de calidad fisiológica de semillas en líneas de maíz. Fitotecnia Mexicana, 26 (001), 1.
20. Alfaro, Y., y Segovia, V. (2000). Maíces del sur de Venezuela clasificados por taxonomía numérica. I. Caracteres de la planta y de la mazorca. Agronomía Tropical, 50 (3), 413-433.
21. Grant, U. J., Hatheway, W. H., Timothy, D. H., Cassalett, C. D., y Roberts, L. M. (1965). Razas de maíz en Venezuela. Instituto Colombiano Agropecuario. Bogotá, Colombia: ABC.
22. Sevilla, P. R. (1991). Diversidad del maíz en la región andina Experiencias en el cultivo del maíz en el área andina.Quito, Ecuador.: IICA-BID-PROCIANDINO.
23. Fontana, N., y González, C. (2000). El maíz en Venezuela. Enfermedades del maíz en Venezuela Caracas. 529: Fundación Polar.
24. Goodman, M. M., y Paterniani, E. (1969). The races of maize. III. Choices of appropiate characters for racial classification. Econ. Bot., 23, 265-273.
25. Seefoó, J. L.; Keibach, N. M. (2010). Ciencia y paciencia campesina. El maíz en Michoacán. El Colegio de Michoacán. Gobierno del estado de Michoacán. Secretaría de Desarrollo Rural.
26. Alvarez, A., y Lasa, J. M. (1990). Populations of maize from Cantabria. I. Morphological evaluation and variability. An. Aula Dei, 20, 1-2.
27. Llaurado, M., y Moreno, J. (1993). Classification of northen Spanish populations of maize by methods of numerical taxonomy. I. Morphological traits. Maydica, 38, 15-21.
Recibido 12/07/2010, aceptado 12/05/2011.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}