










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Medicina Tropical
versión On-line ISSN 1561-3054
Rev Cubana Med Trop v.59 n.3 Ciudad de la Habana sep.-dic. 2007
ARTÍCULO ORIGINAL
Caracterización microbiológica de cepas de Vibrio cholerae no-O1 aisladas en Cuba
Microbiological characterization of non-01 Vibrio cholerae isolated in Cuba
Dra. Laura Bravo FariñasI; Ing. Anabel FernándezII; Dra. María M. RamírezIII; Dra. Alina LlopIV; Dr. Gerardo MartínezV; Lic. Raquel I. HernándezVI; Lic. Luis E. CabreraVII; Lic. Luis MorierVIII; Dr. Jorge FragaIX; Dr. Fidel A. NúñezX; Dr. Adalberto AguilaXI
I Doctora en Ciencias de la Salud. Investigadora Titular. Instituto de Medicina Tropical "Pedro Kourí" (IPK).
II Ingeniera Química. IPK.
IIIEspecialista de II Grado en Microbiología. Investigadora Auxiliar. IPK.
IV Doctora en Ciencias Médicas. Especialista de II Grado en Microbiología. Investigadora Titular. Profesora Titular. IPK.
V Especialista de II Grado en Microbiología. Investigador Auxiliar. IPK.
VI Licenciada en Microbiología. Centro Provincial de Higiene y Epidemiología. Camagüey.
VII Máster en Bacteriología-Micología. Centro Municipal de Güines.
VIII Licenciado en Microbiología. Investigador Auxiliar. IPK.
IX Máster en Parasitología. Investigador Agregado. IPK.
X Doctor en Ciencias Médicas. Especialista de II Grado en Microbiología. Investigador Titular. IPK.
XI Especialista de I Grado en Microbiología. IPK.
RESUMEN
Se estudiaron 422 cepas de Vibrio cholerae no-O1 procedentes de 9 provincias del país, de ellas 9 aisladas de un brote de enfermedad de transmisión hídrica. En la totalidad de las cepas se determinó la susceptibilidad antimicrobiana y la presencia de factores de virulencia. En las 9 cepas procedentes del brote, se realizó además, el estudio de macrorrestricción de ADN mediante la técnica de electroforesis de campo pulsado. Se demostró por primera vez en Cuba y el Caribe, la circulación de cepas de V. cholerae no-O1 atípicas (resistentes al compuesto vibriostático O129 y al trimetoprim-sulfametoxazol). El comportamiento de la susceptibilidad antimicrobiana demostró por primera vez la circulación en Cuba de 2 patrones diferentes de resistencia (ampicilina, trimetoprim/sulfametoxazol, sulfonamida y tetraciclina, trimetoprim/sulfametoxazol, sulfonamida). La frecuencia de cepas resistentes al trimetoprim-sulfametoxazol se mantuvo similar en todo el período de estudio. Sin embargo, se produjo un decrecimiento de la resistencia a la ampicilina y un aumento de la resistencia a la tetraciclina. Los principales factores de virulencia que se encontraron fueron la gelatinasa, la hemolisina, la elastasa y la adherencia a células HEp-2. Sin embargo, las cepas del brote mostraron mayores porcentajes que el resto, para la presencia de la toxina termoestable y la presencia de fimbrias. Los resultados de los estudios molecular y epidemiológico permitieron dar una respuesta acelerada y precisa sobre la etiología del primer brote de enfermedad de transmisión alimentaria.
Palabras clave: Vibrio cholerae no-O1, resistencia antimicrobiana, factores de virulencia.
SUMMARY
The study of 422 non-01 Vibrio cholerae strains from nine provinces, 9 of them isolated from a water-borne disease outbreak, was performed. All the strains exhibited antimicrobial susceptibility and virulence factors. The nine strains from the outbreak were subjected to a DNA macrorestriction study based on the pulsed field electrophoresis technique. For the first time in Cuba and the Caribbean, the circulation of atypical non-01 V. Cholerae strains (resistent to vibriostatic compound 0129 and trimethoprim/sulfamethoxazole). The behavior of antimicrobial susceptibility evinced for the first time the circulation of two different resistence patterns in Cuba (ampicilline, trimethoprim/sulfamethoxazole, sulfonamide and tetracycline, trimethoprim/sulfamethoxazole, sulfonamide). The frequency of trimethoprim/sulfamethoxazole-resistent strains was similar during the whole period of study. However, resistance to ampicilline decreased whereas resistance to tetracycline increased. The main found virulence factors were gelatinase, hemolysine, elastase and adherence to Hep-2 cells. On the other hand, the outbreak strains showed higher percentages than the others due to the presence of heat-liable toxin and fimbriae. The results of the molecular and epidemiological studies allowed giving a speedy and accurate response that explained the etiology of the first food-borne disease outbreak.
Key words: Non-01 Vibrio cholerae, antimicrobial resistence, virulence factors.
INTRODUCCIÓN
Los miembros del género Vibrio son bacilos gramnegativos, anaerobios facultativos y oxidasa positiva, pertenecientes a la familia Vibrionaceae, presentando entre una de sus características fenotípicas la sensibilidad al compuesto vibriostático O/129.1 Este género está integrado por 65 especies, se identificaron 12 patógenas para el hombre, las de mayor importancia clínica son Vibrio cholerae, Vibrio parahaemolyticus y Vibrio vulnificus.1
Vibrio cholerae de acuerdo con la composición del lipopolisacárido somático tiene más de 200 serogrupos, divididos en O1 y no-O1. Entre ellos, solo los serogrupos O1 y O139, son los responsables de epidemias y pandemias de cólera, que han llevado a la muerte a miles de personas en el mundo.1
Vibrio cholerae no-O1 ha sido asociado con casos esporádicos y con brotes de gastroenteritis aguda en diferentes países. La mayoría de las cepas no producen toxina colérica (TC), sino una enterotoxina termoestable (ST), semejante a la producida por Escherichia coli enterotoxigénica (constituye el factor de virulencia principal asociado a la patogenicidad en este microorganismo) y una citotoxina tipo Shiga similar a la producida por Shigella dysenteriae tipo I, relacionada con su invasividad.2 En Cuba la prevalencia de cepas de Vibrio cholerae no-O1 de origen clínico que portan el gen para la toxina termoestable es más alta que la reportada en otras áreas geográficas, excepto en caso de epidemias.3 Otros factores de virulencia presentes en este género, son la producción de enzimas extracelulares (gelatinasa, elastasa, lecitinasa), citolicinas, citotoxinas, hemolisinas, producción de polisacárido capsular, hemaglutininas y toxina termoestable, algunos de los cuales solo son investigados en caso de brotes.4
La forma clínica más común producida por este microorganismo es la gastroenteritis aguda, cuyo espectro clínico es muy parecido al del cólera. Generalmente, la enfermedad es leve y tiene una evolución autolimitada, que no requiere de terapia antimicrobiana; sin embargo, en diarreas profusas, prolongadas con deshidratación severa, moderada y no autolimitable, en las infecciones extraintestinales (septicemia, otitis, entre otras) y en pacientes inmunodeprimidos es necesario el uso de agentes antimicrobianos. Los agentes de elección en los adultos son las tetraciclinas, cloranfenicol y el trimetoprim sulfametoxazol. En niños menores de 9 años debe usarse el trimetoprimsulfametoxazol, y en las embarazadas la ampicilina es el agente antimicrobiano más seguro, se recomienda como agentes alternativos las fluoroquinolonas (ciprofloxacina y ofloxacina).5
Por lo antes expuesto y por la importancia que reviste a los niveles nacional e internacional la vigilancia microbiológica y epidemiológica de Vibrio cholerae no-O1, así como la emergencia y diseminación de cepas resistentes a los agentes antimicrobianos, se propuso realizar la caracterización fenotípica en cepas de Vibrio cholerae no-O1 aisladas, de heces de pacientes con enfermedad diarreica aguda (EDA) en un período de 10 años y así contribuir al mejor conocimiento del potencial patogénico de este microorganismo en Cuba.
MÉTODOS
Se estudió un total de 422 cepas de Vibrio cholerae no-O1, aisladas de heces de pacientes con EDA, procedentes de 9 Laboratorios de Microbiología clínica de los Centros Provinciales de Higiene y Epidemiología (CPHE) del país, de ellas 9 procedían de un brote de transmisión hídrica (acompañado de su encuesta epidemiológica) ocurrido en la provincia de Camagüey en 2001. La totalidad de las cepas fueron remitidas al Laboratorio Nacional de Referencia de Enfermedades Diarreicas Agudas del Instituto de Medicina Tropical "Pedro Kourí"(LNR/EDA/IPK), en el período comprendido de enero de 1996 a diciembre de 2005.
Las 422 cepas mantenidas en medio de conservación de Pasteur fueron inoculadas en caldo cerebro-corazón, e incubadas a 37 °C durante 18 a 24 h en condiciones de aerobiosis. Posteriormente fueron sembradas en placas de agar tiosulfato citrato sales biliares sacarosa (TCBS) e incubadas en las mismas condiciones de temperatura, tiempo y aireación antes mencionadas, al cabo del cual se seleccionaron al menos 3 colonias por placa, que fueron inoculadas en los medios de diferenciación primaria agar hierro y 2 azúcares de Kligler (AHK) y agar hierro lisina (AHL). Se escogieron aquellas colonias que después del crecimiento durante 18 a 24 h en condiciones de aerobiosis a 37 °C, presentaron las características correspondientes al género Vibrio. La confirmación en especie, la serotipificación y el estudio de la sensibilidad al compuesto vibriostático O/129 se realizó según lo descrito en la literatura especializada.1
La susceptibilidad antimicrobiana se llevó a cabo utilizando los agentes antimicrobianos de elección: tetraciclina (TE), cloranfenicol (CL), trimetoprim-sulfametoxazol (SXT), ampicilina (AM) y sulfonamidas (SSS). Para las cepas recibidas a partir de 2001 (197 cepas), se estudiaron además, las drogas alternativas ciprofloxacina y ofloxacina. Se utilizó el método de difusión en agar (Kirby Bauer) en la totalidad de las cepas. Para la lectura e interpretación de los halos de inhibición se utilizaron los protocolos recomendados por el Comité de Estándares de Laboratorio Clínico de 2003 (NCCLS) y las del Instituto de Normas de Laboratorio Clínico (CLSI) de los EE. UU., 2005 (National Committee for Clinical Laboratory Standards. Performance standards for antimicrobial susceptibility testing: eight informatical supplement. M 2-A8. 2003. p. 74) (Clinical and Laboratory Standards Institute. Performance standards for antimicrobial susceptibility testing: eleventh informational supplement. Wayne; 2005. CLSI Document M100-S15).
En la totalidad de las cepas fueron investigadas las enzimas extracelulares DNasa, elastasa, gelatinasa, lecitinasa y ß hemolisina, según lo descrito por Mac Faddin.6 Además, fueron estudiados otros factores de virulencia como la adherencia a células HEp -2, según Carello y otros,7 la presencia de fimbrias siguiendo la metodología descrita por Nishikawa y otros.8 La determinación de la actividad biológica de la toxina termoestable fue investigada mediante la prueba de inoculación intragástrica en ratones lactantes y los valores mayores o iguales a 0,083 (relación cuerpo/intestino) se consideraron positivos.9
En las 9 cepas pertenecientes al brote se llevó a cabo el análisis de macrorrestricción del ADN cromosomal mediante una minielectroforesis en gel de campo pulsado utilizando el sistema Guefast 06.10 El ADN de las cepas de Vibrio cholerae fue preparado con una solución no enzimática que contenía solo agentes químicos.10 Los bloques de agarosa ADN fueron incubados a 37 °C durante 24 h en presencia de 5 U de enzima NotI (Promega). El ADN se visualizó en el minigel mediante una tinción con bromuro de etidio a 0,5 µg/mL durante 30 min y después se observó bajo un transiluminador de luz ultravioleta.10 Los patrones de ADN por la técnica de electroforesis en campo pulsado (ECP) fueron comparados visualmente. La similitud entre los tipos fue evaluada por el programa informático Free Tree-freeware11 que realiza el análisis estadístico de variables múltiples. Los perfiles de los patrones de ADN fueron comparados utilizando el coeficiente de Dice.12
Como marcador de peso molecular se utilizó el fago Lambda (l, 48,5kb- 100kb). Como cepas de referencia en general se utilizaron: Staphylococcus aureus 25923 ATCC; Pseudomona aeruginosa 27853 ATCC; Enterococcus faecalis 29212 ATCC; Escherichia coli 25922 ATCC, y las cepas de V. cholerae 569B; V. cholerae O139; V. cholerae no-O1 (CO32) ST+; E. coli K99 ST-, pertenecientes al LNR/EDA/IPK.
Los análisis estadísticos fueron realizados usando pruebas de proporciones para comparar los porcentajes. La prueba exacta de Fisher fue usada para el análisis de los resultados de las cepas del brote. En todos los casos las diferencias fueron consideradas estadísticamente significativas cuando el valor de p fue menor que 0,05. Todos los análisis fueron desarrollados empleando los paquetes de programas para análisis estadísticos SPSS versión 10.0.1 para Windows13 y EPIINFO, versión 6.04.14
RESULTADOS
Las 422 cepas en estudio resultaron ser bacilos gramnegativos anaerobios facultativos, positivos para las pruebas de oxidasa, lisina y ornitina decarboxilasa y negativos para la arginina dihidrolasa. Todas fueron mótiles, con producción de indol, crecimiento hasta 6 % de NaCl, utilizaron la sacarosa y el manitol, no así el inositol, la arabinosa y la esculina. Todas las cepas resultaron ser Voges-Proskauer (1 % de NaCl) positivo; todas resultaron positivas para la D-galactosa y el citrato de Simmons, y negativas para la hidrólisis de la urea. De las 422 cepas estudiadas, 6 resultaron resistentes al compuesto vibriostático O/129. Ninguna de las cepas en estudio aglutinó con el antisuero polivalente O1 y O139, por lo que quedaron definitivamente identificadas como Vibrio cholerae no-O1.
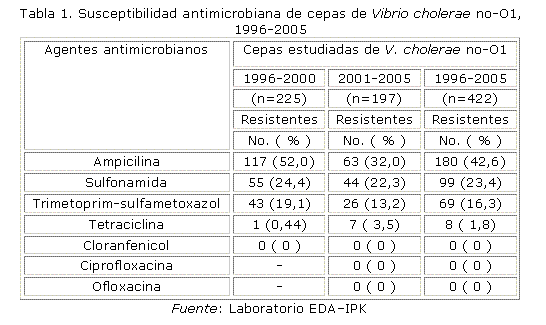
En la tabla 1, se observa la resistencia de las cepas de Vibrio cholerae no-O1 a los diferentes agentes antimicrobianos estudiados, destacándose altos niveles de resistencia a la sulfonamida. Todas las cepas estudiadas en el segundo quinquenio fueron sensibles a los agentes antimicrobianos del grupo de las quinolonas: ciprofloxacina y ofloxacina.
Del presente estudio, se derivaron 2 patrones de resistencia, uno que se mantuvo durante todo el período (AM -, SXT -, SSS) y otro que prevaleció durante el segundo quinquenio (TE -, SXT -, SSS-). Es de destacar que en ambos quinquenios se mantuvo la misma frecuencia de resistencia al trimetoprim-sulfametoxazol y la sulfonamida (p> 0,05), con un decrecimiento significativo de la resistencia a la ampicilina (p< 0,01) y un aumento de la resistencia a la tetraciclina (p< 0,01) en el segundo quinquenio. De las 9 cepas pertenecientes al brote de transmisión hídrica, una resultó sensible tanto a los agentes de elección como a las alternativas, el resto de las cepas resultaron resistentes a la ampicilina. No se observó multirresistencia en ninguna de las cepas.
En las 50 cepas estudiadas en 2001, 6 (12 %) mostraron resistencia al compuesto vibriostático O/129 y al trimetoprim-sulfametoxazol de forma conjunta, por lo que quedaron identificadas como cepas atípicas.
Al investigar los factores de virulencia (tabla 2) se observó como valores significativos la presencia de hemolisina y gelatinasa en 100 % de los aislamientos. Las 9 cepas pertenecientes al brote de transmisión hídrica fueron positivas para 7 de los 8 factores de virulencia investigados, y mostraron mayores porcentajes que el resto de las cepas de casos esporádicos (Prueba de Fisher, p< 0,05).
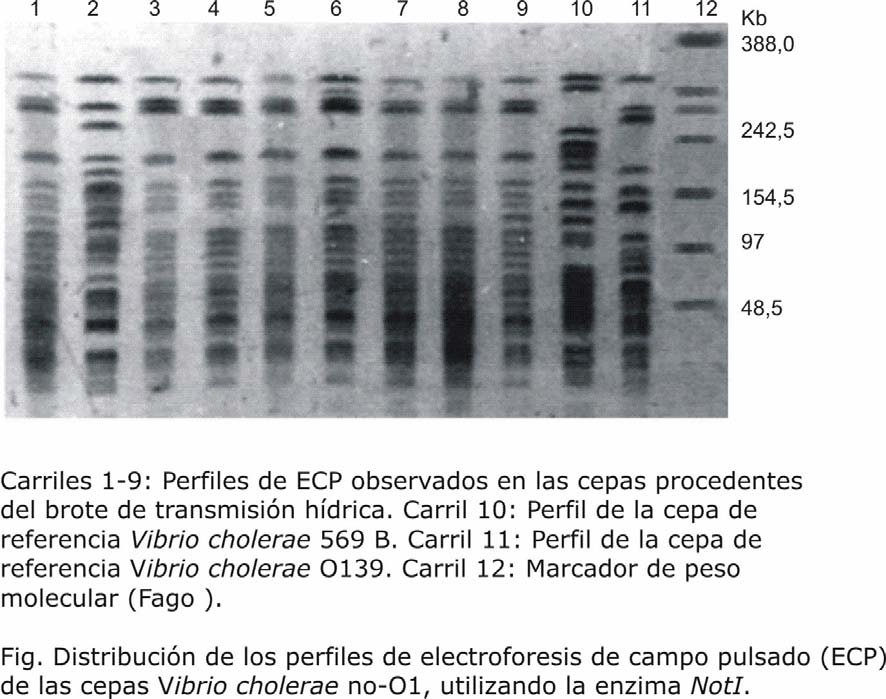
Como resultado de la ECP en las 9 cepas procedentes del brote de transmisión hídrica, utilizando la enzima de restricción NotI se obtuvieron 4 patrones de bandas diferentes, se distinguieron de 26 a 28 bandas por cada aislamiento y se obtuvieron fragmentos con una talla entre 388 kb y 48,5 kb (fig.).
El análisis de agrupamiento permitió identificar un grupo que estuvo conformado por las cepas 1, 3, 4, 5, 6, 7, 8, 9; que son las mismas 8 cepas que fueron resistentes a la ampicilina, y positivas a los factores de virulencia: adherencia a células HEp-2 y toxina termoestable. De estas 8 cepas, la 3, 4, 5, 7, y 8 presentaron los mismos fragmentos de restricción y resultaron indistinguibles (se interpretaron como cepas causantes del brote). Las cepas 1, 6, 9 tenían un nivel de similitud de 98 % en relación con las 5 antes mencionadas, esto indica la estrecha correspondencia y que quizá estuvieran genéticamente muy relacionadas (fig.).
El coeficiente de Dice resultó de 0,85.
DISCUSIÓN
Patógenos reemergentes como Vibrio cholerae O1 y Vibrio cholerae no-O1 están sujetos a medidas de prevención, control, vacunas, estudios clínicos-epidemiológicos, de factores de virulencia y de susceptibilidad a drogas antimicrobianas.15
La caracterización fenotípica y antigénica de las 422 cepas de Vibrio cholerae no-O1 objeto de este estudio resultó similar a la obtenida por Ou y otros, quienes evaluaron 75 episodios diarreicos de 73 pacientes con diarreas invasivas en África. Se identificó con los mismos métodos microbiológicos 90 % de las cepas pertenecientes a este mismo género y especie.16
Numerosos estudios de susceptibilidad antimicrobiana han sido realizados en el género Vibrio, los que han evidenciado su sensibilidad a los aminoglucósidos, cloranfenicol, ácido nalidíxico, sulfonamidas y fluoroquinolonas. La sensibilidad al trimetoprim-sulfametoxazol y la tetraciclina han resultado variables.17
Analizando los resultados de la susceptibilidad antimicrobiana en este estudio, se observó que las cepas presentaron resistencia moderada a la sulfonamida. Similar nivel de resistencia fue encontrado por Iwanaga y otros, quienes al estudiar 50 cepas de Vibrio cholerae no-O1 aisladas de pacientes con EDA en Laos, encontraron entre 33 y 39 % de resistencia a la sulfonamida.18
Los resultados del presente estudio en relación con la ampicilina están acorde con los publicados por Tjaniadi y otros, quienes en una investigación realizada en 8 provincias de Indonesia encontraron 29 % de resistencia a la ampicilina, evidenciándose así, la generalización de la resistencia a este agente antimicrobiano en diferentes áreas geográficas.19 El descenso de la frecuencia de cepas cubanas resistentes a la ampicilina en el período 2001-2005 pudiera explicarse por el cambio en la política de antibiótico en cuanto al tratamiento de otras enfermedades diarreicas agudas (shigelosis), en el cual se sustituyó la ampicilina por ácido nalidíxico.20
Los bajos niveles de resistencia a la tetraciclina y al cloranfenicol encontrados en este trabajo difieren con los de Kondo y otros en Tailandia, quienes encontraron 100 % de resistencia a la tetraciclina en 57 cepas de Vibrio cholerae no-O1 procedentes de una epidemia.21 Estas discrepancias en relación con las cepas cubanas pudieran atribuirse al uso adecuado de la terapia antimicrobiana en el tratamiento de la enfermedad diarreica aguda.
En un brote de cólera ocurrido en Nigeria en 1995 se evaluó la sensibilidad y resistencia de 79 cepas de Vibrio cholerae O1, y se encontró 97,1 % de sensibilidad al trimetoprim sulfametoxazol.22 Similares resultados (95,1 %) se encontraron en cepas de Vibrio cholerae no-O1 aisladas de pacientes con gastroenteritis aguda en Argentina.23
Los bajos niveles de resistencia para ese agente antimicrobiano encontrados en este trabajo, resultaron similares al de los estudios antes mencionados.
Los resultados de la presente investigación muestran que en Cuba existe un bajo grado de resistencia a los agentes antimicrobianos analizados, lo que indica la necesidad de continuar la vigilancia de estos microorganismos por causa de la existencia de cepas resistentes al compuesto O/129 y al trimetoprin- sulfametoxazol, lo que demanda la práctica de estudios moleculares para explicar el origen de esta doble resistencia.24
Con el objetivo de prevenir la propagación de cepas Vibrio cholerae en Cuba, los resultados de la presente investigación aportarían antibiotipos conocidos, lo que contribuiría a un adecuado tratamiento clínico y epidemiológico.25
Son escasos los estudios sobre factores de virulencia realizados en América Latina y en el Caribe en cepas de Vibrio cholerae no-O1 aisladas de pacientes con EDA.26
Cuando se revisa la bibliografía científica internacional en relación con la producción de proteasas, enzimas extracelulares y citotoxinas, entre otras, en el género Vibrio, se cita el artículo escrito por Boiko y otros donde indica que son comunes en este género, implicadas en la aparición de hemorragia, edema y la alteración de los sistemas de defensa en el organismo; esto favorece el desarrollo de los procesos infecciosos. En las cepas de este estudio se evidenció la producción de las enzimas DNasa, gelatinasa, lecitinasa y elastasa, que coincide con lo señalado en las monografías revisadas sobre propiedades y mecanismo de virulencia en las especies del género Vibrio realizadas por el autor antes citado.27
Estudios efectuados por Bravo y otros en 2000 demostraron la presencia de 20 % de toxina termoestable en cepas de Vibrio cholerae no-O1 aisladas de pacientes con EDA,28 esto coincide con los valores obtenidos en la presente investigación.
Numerosos métodos de tipaje han sido utilizados para el estudio y la caracterización de agentes infecciosos causantes de enfermedades de transmisión alimentaria y enfermedades de transmisión digestiva, los cuales han permitido determinar la relación clonal entre los diferentes aislamientos. Para lograr este objetivo el Centro de Prevención y Control de Enfermedades (CDC) de EE. UU. ha evaluado, a través de la Red Internacional de Pulse Net, diferentes protocolos para el estudio y la caracterización molecular de patógenos entéricos con empleo de la técnica de ECP.29
La aplicación de la técnica de minielectroforesis en campo pulsado permitió conocer que 8 de las 9 cepas implicadas en el brote, estaban relacionadas genéticamente y que 5 de ellas conformaban las cepas del primer brote de enfermedad de transmisión hídrica en Cuba por Vibrio cholerae no-O1. Estos resultados fueron avalados por la correspondiente historia epidemiológica que acompañaba las cepas y contribuyó a determinar su relación clonal. Similares resultados fueron obtenidos por Rudra y otros en la India al evaluar 60 casos clínicos en el desarrollo de una epidemia de cólera.30
Kam y otros, mediante la aplicación de la misma técnica pudieron relacionar 220 cepas de Vibrio cholerae no-O1 y O1 procedentes de brotes diarreicos en China. Ellos demostraron que con la utilización de la enzima NotI pudieron obtener patrones heterogéneos de ADN, lo que les permitió definir que muchas cepas no estaban implicadas en estos brotes.31
El hecho de que las 5 cepas de Vibrio cholerae no-O1 procedentes de Camagüey estuviesen genéticamente relacionadas y que además presentaran el mismo patrón de susceptibilidad antimicrobiana reafirma el hecho de que esas 5 cepas fueron las causantes del brote, junto con las 3 cepas que estuvieron genéticamente relacionadas con ellas. Esto reafirma la necesidad de la vigilancia epidemiológica de Vibrio cholerae no-O1 en Cuba. Los resultados de los estudios microbiológico, molecular y epidemiológico, permitieron dar una respuesta acelerada y precisa sobre la etiología del primer brote de enfermedad de transmisión alimentaria, que permitió constar en el país con los primeros patrones de electroforesis de campo pulsado en cepas de Vibrio cholerae no-O1.
REFERENCIAS BIBLIOGRÁFICAS
1. Garrity GM, Bell JA, Lilburm TG. Taxonomic outline of the prokaryotes. En: Bergey ¢s Manual of systematic bacteriology. 2nd ed. Release 4.00; Oct 2003. [Citado el 10 de febrero de 2004]. Disponible en URL: http://dx.doi.org/10.1007/bergeysoutline200310
2. Nair GB, Oku Y, Takeda Y, Ghosh A, Ghosh RK, Chattopadhyay S, et al. Toxin profiles of Vibrio cholerae non-O1 from environmental sources in Calcutta, India. Appl Environ Microbiol. 1988;54:3180-2.
3. Guglielmetti P, Bravo L, Sanchi A, Giovanna L, Rossolini GM. Detection of Vibrio cholerae no-O1 heat-stable enterotoxin gene by polymerase chain reaction. Mol Cell Probes. 1994;8:101-6.
4. Baffone W, Pianetti A, Bruscolini F, Barbieri E, Citterio B. Occurrence and expression of virulence-related properties of Vibrio species isolated from widely consumed seafood products. Int J Food Microbiol. 2000;54:9-18.
5. Bhattacharya SK. An evaluation of current cholera treatment. Expert Opin Pharmacother. 2003;4(2):141-6.
6. Mc Faddin JF. Pruebas bioquímicas para la identificación de bacterias de importancia clínica. Baltimore:William and Wilkins; 2002.
7. Carrello A, Solburn K, Budden J, Chong B. Adhesion of clinical and environment Aeromonas isolates to HEp-2 cells. J Med Microbiol. 1988;26:19-27.
8. Nishikawa Y, Kimiura T, Kishi T. Mannose-resistant adhesion of motile Aeromonas to INT 407 cells and the differences among isolates from humans food and water. Epidemiol Infect. 1991;107:171-9.
9. Takeda Y, Takeda T, Yano T, Yamamoto K, Miwatani T. Purification and partial characterization of heat-stable enterotoxin of enterotoxigenic Escherichia coli. Infect Immunol. 1979;25:978-85.
10. López-Cánovas L, Bravo L, Herrera J, Riverón AM, Javer E, Sánchez A, et al. DNA fingerprinting of Vibrio cholerae and Aeromonas species by pulsed field minigel electrophoresis. Electrophoresis. 2006;27:2857-64.
11. Pavlicék A, Hrdá L, Flegr J. Free tree-freeware program for construction of phylogenetic trees on the basis of distances data and for bootstrap/jackknife analysis of the threes robustness. Application in the RADP analysis of genus Frenkelina. Folia Biol. (Prague) 1999;45:97-9.
12. Tenover FC, Arbeit RD, Goering RV, Mickelsen PA, Murray BE, Persing DH, et al. Interpreting chromosomal DNA restriction patterns produced by pulsed-field gel electrophoresis: criteria for bacterial strain typing. J Clin Microbiol. 1995;33:2233-9.
13. Bryman A. Quantitative Data Analysis with SPSS Release 10 for Windows. London:Routledge; 2001.
14. Dean AG, Dean JA, Coulombier D, Brendel KA, Smith DC, Burton AH, et al. Epi Info Version 6: A Word Processing, Database, and Statistics Program for Epidemiology on Microcomputers. Atlanta, GA:Centers for Disease Control; 1994.
15. Mc Nicholl, Downer MV, Udhayakumar V, Alper CA, Swedlow DL. Host-patogen in emerging and re-emerging infectious diseases. Annu Rev Public Health. 2000;21:15-46.
16. Ou TY, Liu JW, Leu HS. Independent prognostic factors for fatality in patients with invasive Vibrio cholerae no-O1 infectious. J Microbiol Infect. 2003;2:117-22.
17. Guerrant RL, Van Gilder T, Steiner TS, Thielman NM, Slutsker L, Tauxe RV, et al. Practice guidelines for the management of infectious diarrhea. Clin Infect Dis. 2001;32:331-50.
18. Iwanaga M, Toma C, Miyazato T, Insisiengmay S, Nakasorte N, Ehara M. Antibiotic resistance conferred by a class I integron and SXT constin in Vibrio cholerae O1 strains isolated in Laos. Antimicrob Agents Chemother. 2004;48(7):2364-9.
19. Tjaniadi P, Lesmana M, Subekti D, Machpud N, Komalarini S, Santoso W. Antimicrobial resistance of bacterial pathogens associated with diarrheal patients in Indonesia. Am J Trop Med Hyg. 2003;68:666-70.
20. Ramírez M, Valdés N, Llop A, Bravo L, Fernández A. Perfil plasmídico y resistencia antimicrobiana en cepas de Shigella aisladas en Cuba. Rev Cubana Med Trop. 2004;3:105-7.
21. Kondo S, Kongmuang U, Kalnauwakul S, Matsumoto C, Chen CH, Nishibuchi M. Molecular epidemiologic analysis of Vibrio cholerae O1 isolated during the 1997-8 cholera epidemic in Southern Thailand. Epidemiol Infect. 2001;127:7-16.
22. Olukoya DK, Ogunimi AA, Abaelu AM. Plasmid profiles and antimicrobial susceptibility patterns of Vibrio cholerae O1 strain isolated during a recent outbreak in Nigeria. J Diarrhoeal Dis Res. 1995;13(2):118-21.
23. Rivas M, Cacace ML, Ayala LT, Baschkier A, Miliwebsky E, Caffer MI. Cases of gastroenteritis associated to Vibrio cholerae no-O1 in Oran, Salta. Rev Argent Microbiol. 1996;28:163-9.
24. Bravo L, Cabrera R, Cabrera L, Ramírez M, Castanaeda N, Fernández A, et al. Sensibilidad antimicrobiana de cepas de Vibrio cholerae no-O1 aisladas de pacientes en Cuba. Rev Esp Quimioterap. 2004;17:200-1.
25. Karaolis D, Lan R, Reeves P. The sixth and seventh cholera pandemics are due to independent clones separately derived from environmental nontoxigenic Vibrio cholerae non-O1. J Bacteriol. 1995;177:3191-8.
26. Sack D, Sack R, Nair G, Siddique A. Cholera. Lancet 2004;363:223-33.
27. Boiko AV. Pathogenicity factors of various Vibrios and Aeromonas. Zh Mikrobiol Epidemiol Immunobiol. 2000;79(6):104-8.
28. Bravo L, Ramírez M, Maestre J, Llop A, Cabrera R, García B, et al. Vibrio cholerae no-O1 toxigénico. Rev Cubana Med Trop. 2000;52:106-8.
29. Centers for Diseases Control and Prevention. Standarized molecular subtyping of food borne bacterial pathogens by pulsed-field gel electrophoresis: training manual. Atlanta: Centers for Diseases Control and Prevention; 2000.
30. Rudra S, Mahan R, Mathus M, Kathuria K, Tawar V. Cluster of cases of clinical cholera due to Vibrio cholerae O10 in East Delhi. Indian J Med Res. 1996;103:71-3.
31. Kam K, Luey C, Tsang Y, Law C, Chu M, Cheung T, et al. Molecular subtyping of Vibrio cholerae O1 and O139 by pulsed-field gel electrophoresis in Hong-Kong: correlation with epidemiological events from 1994 to 2002. J Clin Microbiol. 2003;41:4502-11.
Recibido: 20 de febrero de 2007.
Aprobado: 21 de mayo de 2007.
Dra. Laura Bravo Fariñas. Instituto de Medicina Tropical "Pedro Kourí". Km 6 1/2, Autopista Novia del Mediodía. Marianao 13. La Lisa. Ciudad de La Habana. Correo electrónico: laura@ipk.sld.cu


{kind=link}
{kind=link}
{kind=link}