










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Investigaciones Biomédicas
versión On-line ISSN 1561-3011
Rev Cubana Invest Bioméd v.28 n.4 Ciudad de la Habana oct.-dic. 2009
ARTÍCULOS ORIGINALES
Participación del óxido nítrico, proteína Fos y el tallo cerebral en la retención de glucosa encefálica durante la hipoxia
Involvement of the nitric oxide, Fos protein and brain stem in the retention of brain glucose during hypoxia
Mónica Lemus VidalI; Sergio Adrián Montero CruzII; José Luis Cadenas FreixasIII; Sonia Luquín de AndaIV; Joaquín García EstradaV; Roces Elena de Álvarez-Buylla EVI
IDoctora en Ciencias Médicas, Profesora Investigadora Asociada C. Centro Universitario de Investigaciones Biomédicas. Universidad de Colima. México.
IIDoctor en Ciencias Fisiológicas, Profesor Investigador Titular C Facultad de Medicina, Universidad de Colima. México.
IIIDoctor en Ciencias Fisiológicas. Profesor Titular. Universidad de Ciencias Médicas de Camagüey. Cuba.
IVDoctora en Ciencias Médicas, Profesora Investigadora Titular C, Universidad de Guadalajara. México.
VDoctor en Ciencias Médicas, Profesor Investigador Titular C. Centro de Investigaciones Biomédicas de Ocidente. Instituto Médico de Seguro Social. México.
VIBióloga. Profesora Investigadora Titular A. Centro Universitario de Investigaciones Biomédicas. Universidad de Colima. México.
RESUMEN
Se ha descrito que el núcleo del tracto solitario (NTS), estructura del tallo cerebral y vía de relevo de las aferencias del los quimiorreceptores del senocuerpo carotídeo (RSCC), participa en el aumento en la retención de glucosa por el cerebro (RGC) ante una hipoxia. Es probable que en esta respuesta participe el óxido nítrico (NO) y la proteína Fos. En este trabajo se analiza el papel del NO en el NTS sobre la modificación de la RGC y la expresión de la proteína inmunorreactiva Fos (Fos-ir) en ratas in vivo. La inyección de un donador del NO como es el nitroprusiato de sodio (NPS) en el NTS, 4 min antes de la estimulación de los RSCC, disminuyó la RGC, pero incrementó la expresión de Fos-ir en un mayor número de neuronas en el NTS con respecto a las ratas control, que sólo recibieron líquido cefalorraquídeo artificial (LCRa) antes de la estimulación RSCC. En contraste, un inhibidor selectivo del NO como el N?-nitro-L-arginina metil éster (L-NAME) en el NTS 4 min antes de la estimulación RSCC con NaCN, aumentó la RGC, pero disminuyó el número de neuronas Fos-ir comparados con el control o con NPS. La detección inmunohistoquímica de la expresión de Fos-ir en las células del tallo cerebral indica que la estimulación RSCC activa vías dependientes de NO en el NTS, para regular la RGC. El estudio de esta población de células en el NTS, será importante para definir su caracterización.
Palabras clave: óxido nítrico, tallo cerebral, glucosa encefálica, hipoxia, proteína Fos.
ABSTRACT
It has been said that the nucleus tractus solitarii (NTS), one structure of the brain stem and path of apherences of chemoreceptors of carotid sinus-body, is involved in the increased glucose retention by the brain in case of hypoxia. It is likely that nitric oxide and Fos protein also take part in this response. This paper analyzes the role of nitric oxide in the NTS on the change of glucose retention by the brain and the expression of inmunoreactive protein Fos (ir-Fos) in rats in vivo. The injection of a NO donor such as sodium nitroprusiate in the NTS four minutes before the stimulation of carotid sinus-body chemoreceptors decreased glucose retention by the brain but increased the expression of ir-Fos in a higher number of neurons in NTS with respect to control group rats which only received artificial cerebrospinal fluid before the stimulation. In contrast, the use of a selective NO inhibitor such as NO-nitro-L-arginine methyl ester (L-NAME) in the NTS four minutes before the stimulation of the chemoreceptors with NaCN, increased the glucose retention by the brain but reduced the number of neurons with ir-Fos expression when compared with the control group or the sodium nitroprusiate injection. The immunohistochemical detection of ir-Fos expression in the brain stem cells indicated that stimulation of carotid sinus-body chemoreceptors activated NO-dependent paths in the NTS to regulate glucose retention by the brain. The study of this cell population in the NTS will be important to define its characterization.
Key words: nitric oxide, brain ítem, brain glucosa, hypoxia, Fos protein.
INTRODUCCIÓN
Los estudios de Álvarez-Buylla y Roces de Álvarez-Bulla1, Montero y col.2 demuestran que los receptores del senocuerpo carotídeo (RSCC) participan en la homeostasis de la glucosa. Dichos receptores son sensibles tanto a los cambios de la presión parcial de oxígeno (pO2) como a las variaciones en las concentraciones de glucosa en la sangre que los irriga, e inducen cambios reflejos en el sistema nervioso central (SNC) en respuesta a las concentraciones de oxígeno y/o glucosa.3-5 Las vías aferentes de los quimiorreceptores carotídeos llegan al núcleo del tracto solitario (NTS) a través del nervio del seno carotídeo (NSC).6 y participa en la regulación de las funciones respiratoria, cardiovascular y en la homeostasis energética del propio SNC. 7 Se desconocen los mecanismos centrales que median los efectos descritos sobre los niveles de glucosa central y periféricos. Se señala la participación del óxido nítrico (NO) que interviene en el aprendizaje y la regulación de la homeostasis autonómica a distintos niveles del neuroeje, incluyendo el NTS.8 El NO incrementa los niveles de GMPc9 para regular el flujo sanguíneo cerebral10, funciones neuroendocrinas11, funciones cardiovasculares12, la memoria13 y las respuestas fisiológicas ante un estado de hipoxia-isquemia cerebral.14. El NO induce una depleción de ATP celular en el SNC modulando las vías metabólicas durante la glucólisis.15 En experimentos in vitro se demuestra que el NO producido por la NOS neuronal funciona como un neuromodulador inhibitorio en la quimiorrecepción hipóxica.16 Después de estimular los RSCC con NaCN se observa una elevación de la RGC, pero la administración central de NO inhibe este efecto.2 La activación de los barorreceptores carotídeos induce la formación de NO en el NTS, de la proteína Fos y la expresión del proto-oncogén c-fos, a través de la fosforilación de la proteína de unión al elemento de respuesta del AMPc (CREB, por sus siglas en inglés) con la participación de la guanilato ciclasa soluble/GMPc/proteincinasa G1.17,9 El motivo principal de este estudio fue analizar in vivo las vías centrales que participan en el reflejo hiperglucemiante con retención de glucosa por el cerebro inducido por la estimulación de los RSCC, determinando la transmisión nitroxidérgica en la expresión de la proteína Fos en el NTS. Analizar el papel del NO en el NTS sobre la modificación de la RGC y la expresión de la proteína inmunorreactiva Fos (Fos-ir) post-estimulación quimiorreceptora con NaCN en ratas in vivo.
MÉTODO
Animales y procedimientos quirúrgicos: los experimentos se realizaron en ratas Wistar macho de 280-300 g de peso, con 4 meses de edad mantenidas en condiciones de luz-oscuridad 12h/12h, a temperatura ambiente y en ayuno de 12h antes del experimento, anestesiadas con pentobarbital sódico (Anestesal, Pfizer, 3 mg/100 g por vía intraperitoneal-ip), manteniendo el nivel de anestesia constante durante todo el experimento por infusión ip de una solución de pentobarbital sódico (0.063 mg/min). Para la ventilación artificial se utilizó un respirador para pequeñas especies (Stoelting-Ugo Basile) conectado a una cánula endotraqueal (intubación por vía bucal), con frecuencia de 54 respiraciones/min y presión positiva hasta evitar los movimientos respiratorios espontáneos de la rata.4 Se realizó incisión por línea media en cara ventral del cuello. Se cateterizó la vena yugular externa derecha, con ayuda de ganchillos de vidrio, con un tubo de silastic (Dow Corning 602-155) hasta el seno venoso yugular.18 La arteria femoral se disecó y cateterizó hasta la aorta abdominal con un tubo de polietileno (PE-10 Clay Adams). Todas las cateterizaciones se realizaron con tubos heparinizados sin interrumpir la circulación normal en los vasos.18 Al final de cada experimento se verificó la posición correcta de los catéteres. Las ratas se dividieron al azar en 3 grupos: a) LCRa en NTS + NaCN en SC (n = 10); b) nitroprusiato de sodio (NPS) en NTS + NaCN en SC (n = 10);c) N-nitro-L-arginina metil éster (L-NAME) en NTS + NaCN en SC (n = 10);
Estimulación de los RSCC: para estimular los RSCC con NaCN, el seno carotídeo izquierdo se aisló temporalmente de la circulación general (preparación de seno carotídeo aislado).1 Tanto la carótida externa (por encima de la arteria lingual) como la carótida interna izquierda (cerca del foramen yugular) se ocluyeron temporalmente (20 s) durante las inyecciones de NaCN para evitar su paso a la circulación cefálica y/o circulación general. La estimulación quimiorreceptora se efectuó inyectando lentamente NaCN (5 µg/100 g en 0.25 ml de sol. sal.) a través de una aguja del No 27 en la arteria carótida común izquierda, con objeto de evitar la estimulación barorreceptora.19
Obtención de muestras de sangre para análisis de glucosa: se obtuvieron 0.12 ml de sangre arterial (arteria femoral) y 0.12 ml de sangre venosa procedente del cerebro (seno yugular). La colección de sangre se realizó a los tiempos siguientes: t=-10 y -5 min (antes de la aplicación de las drogas en NTS y de la estimulación de los RSCC), y t=5, 10, 20, 30, 40 y 50 min (después de la estimulación de los RSCC); la inyección de drogas en NTS se hizo al t=-4 min (4 min antes de la estimulación de los RSCC).20,21 En cada experimento la extracción total de sangre fue de 3.84 mL (16% del volumen total). Para compensar la pérdida de sangre, las ratas recibieron después de cada muestra de sangre una inyección de 0.2 ml de NaCl al 0.9%. Las muestras de sangre se centrifugaron (centrífuga refrigerada, Beckman T J-6) a 3000 rpm durante 5 min para hacer la determinación de glucosa en plasma por el método de glucosa-oxidasa (analizador Beckman).
Microinyecciones en NTS: después de exponer la superficie dorsal del cráneo de la rata, se fijó la cabeza en un esterotáxico (Stoelting) y se hizo una craneotomía occipital. En el NTS izquierdo, se insertó una aguja de 200 µm de diámetro conectada a una microjeringa (Hamilton, de 0.1 a 0.5 µl) para las inyecciones de las drogas en un volumen de 100 nl durante 20-30 s22. Las coordenadas para abordar el NTS (P=12.7 mm, L=1.45 mm, V=7.7 mm) fueron descritas por Paxinos y Watson.23 Para marcar el punto de inyección, al final de cada experimento, se inyectó azul de metileno (1%, mismo volumen) utilizando la misma microjeringa. La rata se sacrificó por decapitación con guillotina24, removiendo el cerebro que se congeló en ultracongelador (Revco) para después realizar los cortes en críomicrotomo (Leica), teñir con violeta de cresilo y verificar histológicamente el lugar de la inyección.25
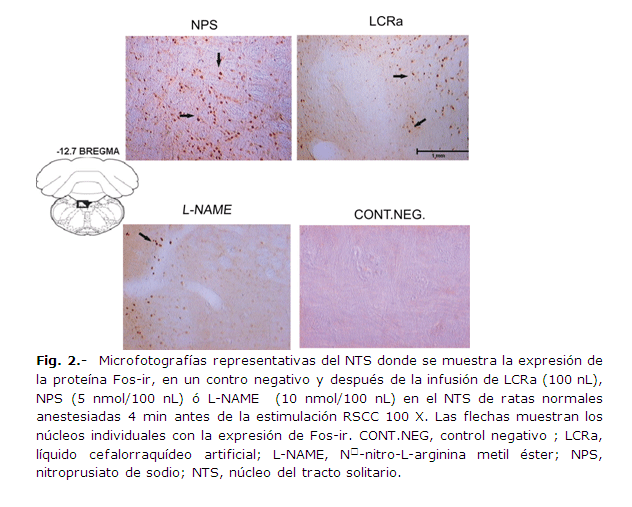
Inmunohistoquímica para la detección de la proteína Fos en el NTS: cinco min después de concluir cada experimento, los animales se perfundieron por vía intracardiaca con paraformaldehído al 4%, se extrajo el cerebro y se post-fijó por inmersión, en el mismo fijador durante toda la noche. Se hicieron cortes transversales seriados de 50 µm de espesor de la médula oblongata para identificar el NTS (vibratomo Leica VT 1000E), se procesaron para visualizar Fos-ir por inmunohistoquímica.26 Como control negativo, se incubó un corte histológico semejante a los anteriores en una solución que contenía únicamente anticuerpo secundario anticonejo biotinilado. Las secciones de tejido se montaron en un portaobjetos con gelatina y sulfato de potasio crómico (CKS) para secarse al aire o con glicerol. Los mismos cortes se utilizaron para localizar la punta de la micropipeta empleada en las microinyecciones, facilitada por la tinción con azul de Evans. Las células Fos-ir positivas se contaron manualmente unilateralmente en cada sección, por dos observadores, utilizando un objetivo de cámara lúcida en un microscopio Zeiss equipado con una cámara digital (VGA), y registrando el promedio de células Fos-positivas (Fos-ir) dentro de los confines del NTS27, la diferencia en el conteo realizado por los dos observadores no fue estadísticamente significativa (P>0.05, t de Student).
Estadística: los resultados obtenidos son medias aritméticas ± error estándar. La comparación estadística entre grupos se realizó por la prueba de ANOVA, fijando la significancia a P<0.05.
RESULTADOS
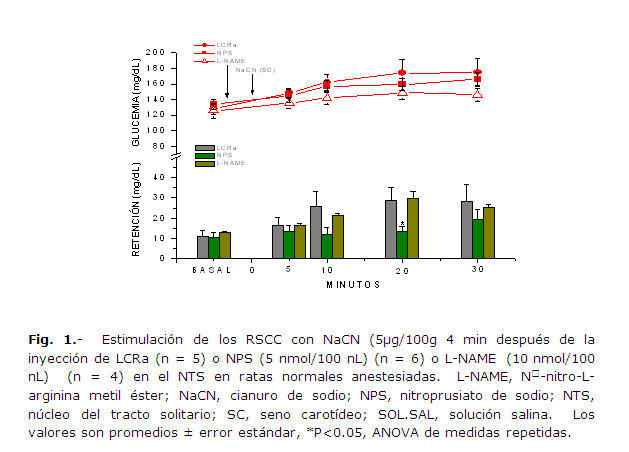
En la fig. 1 se presentan, para su comparación, los resultados obtenidos en relación con la glucemia arterial y la retención de glucosa cerebral en las ratas normales anestesiadas, después de la infusión de LCRa, NPS ó L-NAME con estimulación de los RSCC. La estimulación RSCC 4 min después de la infusión de NPS provocó un descenso significativo en la retención de glucosa cerebral al t = 20 min; cuando este resultado se comparó con el resultado obtenido con la inyección de LCRa antes de la estimulación RSCC, la RGC bajó de 28.6 ± 6.7 mg /dL hasta 13.28 ± 2.7 mg/dL (P = 0.029, ANOVA de medidas repetidas, 2 grupos). Por otro lado, la infusión de L-NAME en el NTS antes de la estimulación de los RSCC no produjo cambios significativos en la RGC en ninguno de los tiempos analizados (P = 0.94, ANOVA de medidas repetidas, 2 grupos). Tampoco se observaron cambios significativos en la glucemia arterial al comparar las curvas obtenidas después de la estimulación de los RSCC precedida por las inyecciones de los distintos fármacos en el NTS.
Cuantificación de la expresión de la proteína Fos-ir en el NTS
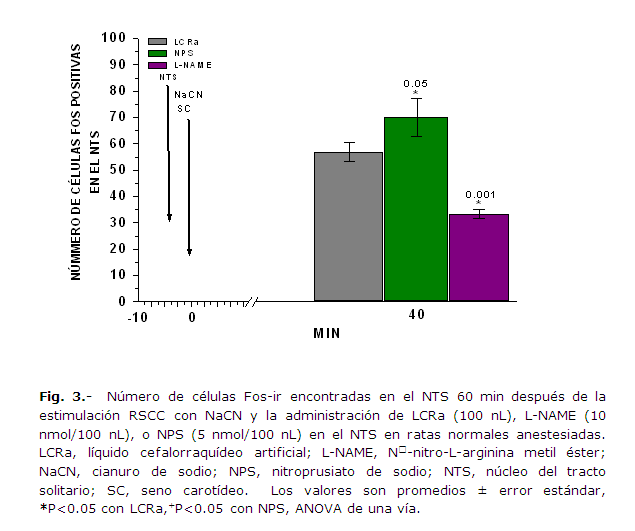
La infusión de NPS en el NTS, antes de la estimulación anóxica de los RSCC, indujo un incremento significativo en la expresión de Fos-ir en las células del NTS, en comparación con las ratas del grupo control que sólo recibieron LCRa (P = 0.050, ANOVA de una vía para muestras independientes); a los 40 min (tiempo transcurrido entre la estimulación RSCC y el inicio de la perfusión con la solución de paraformaldehido) de la estimulación; por el contrario, en las mismas condiciones, la infusión de L-NAME, disminuyó de manera significativa la expresión de la proteína Fos-ir en comparación con la expresión obtenida en las ratas control (P = 0.000001 ANOVA, de una vía para muestras independientes) y con la obtenida en las ratas que recibieron el NPS en el NTS (P =0.00000000003 ANOVA, de una vía para muestras independientes) (Figs. 2 y 3). La presencia de Fos-ir en las neuronas del NTS constituye un correlato neuronal de la participación de este núcleo en los reflejos glucémicos iniciados por la estimulación anóxica de los quimiorreceptores carotídeos.
DISCUSIÓN
Las inyecciones de LCRa en el NTS, 4 min antes de la estimulación de los RSCC con NaCN en el seno carotídeo, aumentaron significativamente la concentración de glucosa en la sangre arterial así como la retención de glucosa por el cerebro. Álvarez-Buylla y col.1,4 plantean la existencia de vías aferentes y efectoras conectadas a elementos centrales para regular los niveles de glucosa en el SNC y en la sangre. El NaCN en el SC produce una anoxia histotóxica que estimula la glucogenolisis hepática.28 El NPS en el NTS 4 min antes de la estimulación de los RSCC, produjo un aumento significativo en la expresión de las células Fos positivas, en la concentración de glucosa en la sangre arterial, pero la retención de la glucosa cerebral no mostró variaciones significativas. En las neuronas del NTS es posible detectar Fos-ir después de la activación baro o quimiorreceptora.29 La microinyección unilateral de L-arginina (donador de NO) en el NTS produce un efecto depresor sobre los reflejos cardiovasculares.30 La oclusión bilateral de la arteria carótida común induce la liberación de NO en el NTS con elevación de la presión arterial sistémica.31 El L-NAME en el NTS 4 min antes de la estimulación de los RSCC disminuyó significativamente la expresión de la proteína Fos-ir e inhibió el reflejo hiperglucemiante sin inhibir el aumento en la retención de glucosa encefálica observado en las ratas control. El L-NAME inyectado en el NTS atenúa significativamente los efectos cardiovasculares del glutamato32 y de la adenosina30; reduce la hiperglucemia inducida por 2-deoxiglucosa33; y también atenúa los reflejos baro y cardiopulmonares.34 La expresión de la proteína Fos-ir en el NTS después de la estimulación de los quimiorreceptores del CC con NaCN, constituye un indicador de la actividad de las neuronas nitroxidérgicas.35 Estos resultados indican que el NO incrementa la actividad de las neuronas nitroxidérgicas en el NTS, centro de relevo de los estímulos aferentes de los RSCC, transmitidos a otros centros superiores del cerebro, ante una estimulación anóxica que modifica la retención de glucosa cerebral.
Agradecimientos
CONACyT y Fondo Ramón Álvarez-Buylla de Aldana.
REFERENCIAS BIBLIOGRÁFICAS
1. Álvarez-Buylla R, y Roces de Álvarez-Buylla E. Hypoglycemic conditioned reflex in rats: preliminary study of its mechanism. Journal of Comparative Physiology and Psychology. 1975;88: 155-160.
2. Montero S, Cadenas JL, Lemus M, Álvarez- Buylla E, y Álvarez- Buylla R. Nitric oxide in brain glucose retention after carotid body receptors stimulation with cyanide in rats. Advances in Experimental Medicine and Biology. 2006;580: 293-300.
3. Eyzaguirre C, Zapata P. Perspectives in carotid body research. Journal of Applied of Physiology. 1984;57: 931-957.
4. Álvarez-Buylla R, Álvarez-Buylla E. Carotid sinus receptors participate in glucose homeostasis. Respiration Physiology. 1988;72: 347-360.
5. López-Barneo J. Oxygen and glucose sensing by carotid body glomus cells. Current Opinion on Neurobiology. 2003;13:493-9.
6. Housley GD, Sinclair JD. Localization by kainic acid lesions of neurons transmitting the carotid chemoreceptor stimulus for respiration in rat. Journal of Physiology 1988;406:99-114.
7. Mifflin SW. Convergent carotid sinus nerve and superior laryngeal nerve afferent inputs to neurons in the NTS. American Journal of Physiology, 1996;271, R870-R880.
8. Wang S, Paton JF, y Kasparov S. Differential sensitivity of excitatory and inhibitory synaptic transmission to modulation by nitric oxide in rat nucleus tractus solitarii. Experimental Physiology. 2007;92:371-382.
9. Chan SH, Chang KF, Ou CC, Chan JY. Nitric oxide regulates c-fos expression in nucleus tractus solitarii induced by baroreceptor activation via cGMP-dependent protein kinase and cAMP response element-binding protein phosphorylation. Molecular Pharmacology. 2004;65:319-25.
10. Bolanos JP, Almeida A. Roles of nitric oxide in brain hypoxia-ischemia. Biochemical and Biophysical Acta. 1999; 1411:415-36.
11. Lewis SJ, Ohta H, Machado B, Bates JN, Talman WT. Microinjection of S-nitrosocysteine into the nucleus tractus solitarii decreases arterial pressure and heart rate via activation of soluble guanylate cyclase. European Journal of Pharmacology. 1991;202:135-136.
12. Pechánová O, Jendeková L, Vranková S. Effect of chronic apocynin treatment on nitric oxide and reactive oxygen species production in borderline and spontaneous hypertension. Pharmacol Rep. 2009;61:116-22.
13. Sunico CR, Portillo F, González-Forero D, Moreno-López B. Nitric oxide-directed synaptic remodeling in the adult mammal CNS. Journal of Neuroscience, 2005;25:1448-1458.
14. Marks KA, Mallard CE, Roberts I, Williams CE, Gluckman PD, Edwards AD. Nitric oxide synthase inhibition attenuates delayed vasodilation and increases injury after cerebral ischemia in fetal sheep. Pediatric Research. 1996;40:185-191.
15. Almeida A, Cidad P, Delgado-Esteban M, Fernández E, García-Nogales P, Bolaños JP. Inhibition of mitochondrial respiration by nitric oxide: its role in glucose metabolism and neuroprotection. J Neurosci Res. 2005;79:166-71.
16. Prabhakar NR. Neurotransmitters in the carotid body. Adv Exp Med Biol. 1994;360:57-69.
17. Chan RK, Sawchenko PE.Organization and transmitter specificity of medullary neurons activated by sustained hypertension: implications for understanding baroreceptor reflex circuitry. J Neurosci. 1998;18:371-87.
18. Alvarez-Buylla R, Bencosme SA. Reflex hypoglycemia initiated by insulin. Acta Physiol Lat Am. 1981:31:1-11.
19. ALVAREZ BUYLLA R. Dissociation of chemo- and baroreceptor activities in cat: oscillographic study. Arch Inst Cardiol Mex. 1954;24:26-37.
20. Montero SA, Yarkov A, Alvarez-Buylla R. Carotid chemoreceptors participation in brain glucose regulation. Role of arginine-vasopressin. Adv Exp Med Biol. 2000;475:749-60.
21. Yarkov A, Montero S, Lemus M, Roces de Alvarez-Buylla E, Alvarez-Buylla R. Arginine-vasopressin in nucleus of the tractus solitarius induces hyperglycemia and brain glucose retention. Brain Res. 2001;902:212-22.
22. Lemus M, Montero S, Cadenas JL, Lara JJ, Tejeda-Chávez HR, Alvarez-Buylla R, de Alvarez-Buylla ER. GabaB receptors activation in the NTS blocks the glycemic responses induced by carotid body receptor stimulation. Auton Neurosci. 2008;141:73-82.
23. Paxinos G, Watson C. The Rat Brain in Stereotaxic Coordinates. New York: Academic Press 1986.
24 Waynforth HB, Flecknell PA. Experimental and surgical technique in the rat. Acad.Press. London. 1992.
25. Iarkov AV, Kovalev GI, Gal'chenko AA. Changes in the electrical activity of the rat brain during the central administration of different doses of GABA agonists and antagonists. Eksp Klin Farmakol. 1994;57:6-11.
26. Kobelt P, Tebbe JJ, Tjandra I, Bae HG, Ruter J, Klapp BF, Wiedenmann B, Monnikes H. Two immunocytochemical protocols for immunofluorescent detection of c-Fos positive neurons in the rat brain. Brain Research. Brain Research Protocols. 2004;13:45-52.
27. Brunton PJ, Meddle SL, Ma S, Ochedalski T, Douglas AJ, Russell JA. Endogenous opioids and attenuated hypothalamic-pituitary-adrenal axis responses to immune challenge in pregnant rats. The Journal of Neuroscience. 2005;25:5117-5126.
28. Zinker BA, Namdaran K, Wilson R, Lacy DB, Wasserman DH. Acute adaptation of carbohydrate metabolism to decreased arterial PO2. American Journal of Physiology: Endocrinology Metabolism. 1994;266:921-9.
29. Shih CD, Chan SH, Chan JY. Participation of Fos protein at the nucleus tractus solitarius in inhibitory modulation of baroreceptor reflex response in the rat. Brain Research. 1996;738:39-47.
30. Lo WC, Jan CR, Wu SN, Tseng CJ. Cardiovascular effects of nitric oxide and adenosine in the nucleus tractus solitarii of rats. Hypertension. 1998;32:1034-1038.
31. Wu WC, Chai CY. Nitric oxide release in the nucleus tractus solitarius during and after bilateral common carotid artery occlusion. Clinical and Experimental Pharmacology and Physiology. 2004; 31: 152-158.
32. Lo WC, Lin HC, Ger LP, Tung CS, Tseng CJ. Cardiovascular effects of nitric oxide and N-methyl-D-aspartate receptors in the nucleus tractus solitarii of rats. Hypertension. 1997;30:1499-503.
33. Sugimoto Y, Yamada J, Yoshikawa T, Horisaka K. Inhibitory effects of nitric oxide synthase inhibitor, N(G)-nitro-L-arginine methyl ester (L-NAME), on 2-deoxy-D-glucose-induced hyperglycemia in rats. Biological and Pharmacological Bulletin. 1997;20:1307-1309.
34. Dias AC, Vitela M, Colombari E, Mifflin SW. Nitric oxide modulation of glutamatergic baroreflex, and cardiopulmonary transmission in the nucleus of the solitary tract. American Journal of Physiology: Heart Circulatory and Physiology. 2005;288:H256-H262.
35. Talman WT, Dragon DN, Ohta H, Lin LH. Nitroxidergic influences on cardiovascular control by NTS a link with glutamate. Annals of the New York Academy of Sciences. 2001;940:169-78.
Recibido: 28 de septiembre de 2009.
Aprobado: 17 de octubre de 2009.
Dra. Mónica Lemus Vidal. Universidad de Colima. México. E mail: mlv@ucol.mx


{kind=link}
{kind=link}
{kind=link}