










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Desde 1933, los ácidos biliares (AB) han sido considerados agentes carcinogénicos, calificados como precursores del proceso secuencial inflamación-cáncer en diversos segmentos del tracto digestivo e interrelacionados con factores medioambientales que intervienen en el desarrollo del cáncer.1 Este hecho fue descrito en los años noventa del siglo XX como modelo secuencial inflamatorio-adenoma-cáncer, del cáncer colorectal (CCR) en humanos, por Fearon y Vogelstein (Fig. 1). En él se expresa una vez más la interacción de los factores medioambientales clásicos, genéticos, epigenéticos y biomoleculares, como determinantes de la progresión de la secuencia de manera lenta, escalonada y con fallas, que conlleva a una pérdida secuencial de la función de los genes supresores de tumores, la activación de oncogenes, la pérdida de cromosomas, retardo de la apoptosis, hasta la aparición del tumor como su expresión clínica.2,3
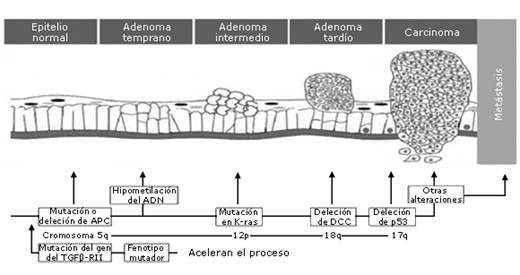
Este modelo, también aplicado a otros órganos del sistema digestivo, fundamentan la correlación entre las diferentes etapas de la carcinogénesis, que transitan desde lesiones intermedias (planas, semiplanas, inflamatorias, úlceras, pólipos inflamatorios, hiperplásicos, adenomatosos, serratos, etc.) hasta el cáncer como lesión más avanzada, según el segmento afectado, las características morfológicas, moleculares, y clínicas.4,5
El objetivo principal de este artículo es describir la asociación entre los niveles elevados de AB en la luz intestinal y la secuencia inflamación-cáncer, expresados como lesiones inflamatorias, premalignas y malignas del tracto digestivo. Asociación fisiopatológica que le permitirá a cirujanos generales, gastroenterólogos, oncólogos, clínicos y fisiólogos, contextualizar la génesis del cáncer en los diferentes segmentos del sistema digestivo y su interacción con los factores de riesgo medioambientales clásicos, genéticos, epigenéticos y biomoleculares que justificaría como criterio de riesgo al cáncer la persistencia de niveles suprafisiológicos de AB en la luz intestinal.
El aporte más reciente de los estudios asociados a la secuencia inflamación-cáncer digestivo nos permitió describir las evidencias biomoleculares del efecto dañino de los AB en la luz intestinal, como constituyente de las bases futuras de la medicina preventiva para el cáncer digestivo.6,7
En este artículo se brindan los fundamentos teóricos actuales de la actividad inflamatorio-carcinogénica de los AB, presentes en pacientes consumidores de dietas ricas en grasas y/o en situaciones clínicas. Entre estas últimas destacan: colelitiasis, hepatopatías crónicas, colecistectomizados, cirugía bariátrica, síndrome de intestino corto, con énfasis en la relación con la biología molecular y la génesis de las lesiones en los diferentes órganos del sistema digestivo.8,9,10,11,12,13
El método empleado para la búsqueda de la información consistió en una revisión sistemática, crítica y actualizada de las evidencias reportadas de alto impacto en las bases de datos internacionales como PubMed, Medline, SciELO, Lilacs, y Elservier, así como, los estudios de observación, caso control, metaanálisis, etc., relacionados con este tema, desde 2015 hasta 2020.
Dicha búsqueda, realizada tanto en idioma español como inglés, utilizó como criterios de selección un índice de materia médica, palabras clave: secuencia inflamación-carcinogénesis y cáncer digestivo; y las abreviaturas AB y ABs, en combinación con los mecanismos biomoleculares del cáncer actualmente propuesto. También procedió una búsqueda manual de referencias citadas por artículos originales y de revisión actuales publicados por el autor principal del presente estudio.
El contenido de los resúmenes o manuscritos en texto completo, seleccionados de la literatura consultada, fueron revisados de manera individual e independiente por el resto de los autores y todos confirmaron que cumplían con los criterios para ser elegidos e incluidos, sin mediar alguna discrepancia ni requerir ajustes posteriores.
Ácidos biliares
Los ácidos biliares actúan como detergente fisiológico y regulador de la homeostasis epitelial intestinal. Por su naturaleza anfipática son solubles en agua y en pH ácidos y resistentes a la precipitación por iones de calcio, propiedades que justifican su permanencia en la luz intestinal y caracterizan su actividad biológica, consistente en: formar micelas, emulsionar las grasas y vitaminas liposolubles y controlar la microbiota intestinal, la proliferación celular, la apoptosis, así como, el metabolismo general y energético en el organismo.14,15
Por tanto, la pérdida de su actividad biológica favorece su exposición mantenida y prolongada sobre los tejidos, donde genera una serie de daños celulares: a nivel de sus membranas y organelos (mitocondria, núcleo, nucléolos, etc.), la producción de RLO y RLN, las mutaciones de genes (oncogenes y gen supresor de tumores), la pérdida de cromosomas, alteraciones en proliferación celular y el retardo de la apoptosis, causante del acúmulo de alteraciones moleculares que implican la inestabilidad genómica, expresados en el modelo de secuencia inflamación-carcinogénesis (Fig. 1).16
En este sentido, los AB, en especial los secundarios (ABs), actúan en condiciones no fisiológicas como agentes precursores o inflamatorio-carcinógenos a nivel de esófago, estómago, intestino delgado, hígado, tracto biliar, páncreas y colon/recto en humanos, y generan todos los daños descritos a nivel celular.17,18
En la práctica clínica, la predisposición genética del individuo al cáncer se desconoce, a no ser que exista un antecedente familiar de primer grado reconocido. Por lo tanto, se infiere que el control de la homeostasis de los AB impide la aparición de lesiones, con independencia de la existencia de una predisposición genética y ha comenzado a ser valorado su control como terapia prometedora en la prevención de lesiones premalignas y malignas del tracto digestivo.19,20
Los AB son metabolizados por la microbiota del colon, donde los primarios (ácido cólico y quenodesoxicólico) son convertidos en ABs (ácidos desoxicólico y litocólico). El desoxicólico, absorbido en el colon, pasa a la circulación enterohepática, se conjuga nuevamente en el hígado y es secretado en la bilis; mientras que el litocólico usa el mismo trayecto, pero es poco reabsorbido por ser insoluble. Por lo tanto, la mezcla de ambos grupos de AB, desde que son reabsorbidos hasta que regresan al hígado, está compuesta de 30-40 % del ácido cólico y quenodesoxicólico; un 20-30 %, de desoxicólico; y menos del 5 %, de litocólico.21,22
Evidencias afirman que en los consumidores de dietas ricas en grasa, de carne roja, fumadores, pacientes con litiasis vesicular y aquellos sometidos a cirugía gastrointestinal (bariátrica, Whipple, síndrome de intestino corto, colecistectomizados, etc.), se estimula el flujo biliar persistente, asociado con cambios estructurales y funcionales no modificables del sistema hepato-vesículo-bilio-intestinal. Dichos cambios acrecientan la concentración fisiológica de los AB en la luz intestinal, hecho que involucra los mecanismos moleculares comunes en la carcinogénesis de los órganos gastrointestinales expuestos a niveles altos de AB y, justifica la inclusión clínica de estos grupos como riesgo al cáncer digestivo.23,24,25,26
Mecanismos celulares y biomoleculares desencadenados por niveles altos de ácidos biliares en la luz intestinal
Los mecanismos propuestos actualmente para explicar el aumento del riesgo de cáncer gastrointestinal y de las lesiones que lo preceden, son las alteraciones del flujo de la bilis (su incremento y persistencia en la luz intestinal) y las del metabolismo de control de los niveles hormonales que actúan como citoprotectores.27) Por tanto, las alteraciones en la síntesis y la transformación o el transporte de los AB, implican trastornos metabolómicos a nivel celular expresados en la secuencia inflamación-carcinogénesis y son considerados, en el concepto novedoso, como biomarcadores de agentes mutagénicos y de lesiones de premalignidad en la carcinogénesis gastrointestinal.28,29
El 90 % del cáncer digestivo es precedido por lesiones premalignas, resultante de un proceso por etapas, lento y de varios años de duración (5-10 años), que involucra factores medioambientales, genéticos y epigenéticos interrelacionados con mecanismos biomoleculares, esto se concibe como la nueva concepción paradigmática, que insta a la comunidad científica a ampliar y desarrollar acciones en su prevención.16
A finales de la década de los noventa del siglo XX, se evidenció que los AB a niveles suprafisiológicos son capaces de promover daños celulares. Su unión y activación con los receptores nucleares, tales como: la proteína quinasa C y el factor de crecimiento epidérmico (FCE), incrementan la permeabilidad celular y paracelular, permitiendo su paso al citoplasma, donde interactúan con los organelos (específicamente, la mitocondria), y desencadenan un evento secuencial, caracterizado por una respuesta inflamatoria aguda con activación del sistema inmunológico (predominio de neutrófilos), que libera los mediadores químicos de la inflamación (citoquinas, RLO, RLN, eucosanoides, neuropéptidos, entre otros), que participan en el control de la tolerancia inmune y en la reparación tisular.
Este evento, en su fase inicial, macroscópicamente se manifiesta en una mucosa eritematosa y edematosa que recupera su aspecto normal al eliminarse el agente agresor.30,31 Si dicha elevación es mantenida, como sucede en los grupos mencionados ut supra, la respuesta inflamatoria es exagerada, persistente y descontrolada; se caracteriza por un infiltrado de linfocitos y plasmocitos que liberan mayor cantidad de mediadores químicos de la fase aguda. Los mediadores químicos son responsables de amplificar y perpetuar la respuesta inflamatoria generadora de daños diversos, desde un mínimo grado hasta los que expresan la evolución anómala de la célula más allá del punto nebuloso de “no retorno”, que culmina en “lesión irreversible y muerte celular”, en el proceso reconocido como secuencia inflamación-cáncer. Pues, sobre todo en individuos con predisposición genética desconocida, esta secuencia conlleva a un incremento de la proliferación celular con alta probabilidad de mutaciones del ADN, desactivación del gen supresor de tumores (proteína p53 mutada) y activación de oncogenes. Precisamente, estos últimos son favorecedores del crecimiento celular y de la displasia en diferentes grados, traducidos morfológicamente por inflamación-adenoma-displasia, que en el 90 % evolucionan al cáncer.32
Desde de los primeros años del siglo XXI se han incrementado los estudios in vitro, con diferentes líneas celulares de cáncer digestivo, cuyos resultados evidencian que los AB dentro del citoplasma celular activan rutas de señalización de la β-catenina, lo que induce el incremento de la fosforilación de tirosinas que disocia la interacción existente entre la β-catenina y la E-caderina y conlleva a la pérdidas en la adhesión, estabilización y translocación de la β-catenina de las células epiteliales.33,34 A este mismo nivel los AB interactúan con organelos como la mitocondria, el núcleo, el nucléolo y el retículo endoplásmico, donde estimulan la producción de RLO y RLN y promueven la salida de calcio, a través de la activación de enzimas como la nicotinamida adenin dinucleótido H oxidasa (NADHO), la fosfolipasa A2 y C; de los receptores FCE y el ligando de Fas, que es una citoquina capaz de activar el programa de la apoptosis en células que expresan el receptor Fas.35,36,37
Estudios realizados en líneas celulares del CCR estimuladas por los ácidos desoxicólico y litocólico, evidencian que la activación de la Fas, la rotura proteolítica de la procaspasa-8 y la activación de la caspasa-8, compromete la cadena respiratoria de la mitocondria con el incremento de los niveles de RLO y RLN, que causan daño al ADN y, por tanto, incrementan y favorecen la mutación de los genes supresores de tumores como el gen p53, que origina una proteína p53 mutada, con pérdida de su función como guardián de la proliferación celular hasta su destrucción y muerte celular.38,39
Otro mecanismo propuesto, por cuanto incrementa la inflamación o retarda la apoptosis de la célula epitelial, es la capacidad de los AB de activar el factor NF-kB.40,41) Los AB también inducen en las células epiteliales inestabilidad genómica por diversos mecanismos (incluyendo la alteración de la mitosis, que da lugar a la aneuploidia), defectos en los controles del conjunto del huso (spindle), daño oxidativo al ADN, que constituye un riesgo aumentado de cáncer digestivo, paro del ciclo celular en G1 y/o G2 junto a un alineamiento inadecuado de los cromosomas en la lámina de metafase y divisiones multipolares, y la muerte celular, tanto por su efecto detergente no específico como, sus interacciones mediadas por los receptores, a nivel de la membrana celular.42
Los niveles elevados de ABs también activan el proceso apoptósico intrínseco, que implica la estimulación del estrés oxidativo en la mitocondria, generación de RLO, liberación de citocromo C y activación de las caspasas citolíticas. Esto ha hecho surgir la teoría de que las células epiteliales del tracto intestinal adquieren resistencia a la apoptosis después de una exposición crónica de niveles altos de AB.43,44
Las células con resistencia apoptósica, inducidas por los efectos genotóxicos de los AB y otros agentes carcinogénicos componentes del lumen intestinal, adquieren también modificaciones genéticas adicionales bajo su acción, que incrementan su agresividad, capacidad proliferativa e invasiva.45) Y si estas se incorporan al proceso de renovación celular del epitelio, la inflamación inicial evoluciona al cáncer.
En Cuba, desde 1992, se desarrolla una línea de investigación en el Instituto de Gastroenterología acerca de la acción tóxica de los AB en el tracto digestivo, que reporta una asociación significativa entre niveles altos de AB totales, tanto en jugo gástrico como en heces y, presencia de lesiones premalignas y malignas en la mucosa gástrica y del colon en pacientes con gastritis alcalina, sometidos a cirugía gástrica, colecistectomizados, resecados de íleon y con litiasis vesicular. Lo anterior hace recomendable la continuidad y profundización de esta investigación en el país, con el fin de justificar la inclusión de estos como grupos de riesgo al cáncer digestivo.39,46,47,48,49,50
Conclusiones
Las evidencias expuestas contextualizan que los niveles altos de AB en cualquier segmento digestivo en interacción o sinergia con los factores medioambientales, genéticos y epigenéticos desencadenan la secuencia inflamación-cáncer según los criterios siguientes, representados en la figura 2:
Originan destrucción focal del epitelio intestinal, que estimula los mecanismos de reparación, seguidos por las reacciones inflamatorias y de hiperproliferación de células indiferenciadas; procesos que causan una transición celular a un estado precanceroso.
Con la activación de las enzimas intracelulares (NADHO, FLP A2) y el daño mitocondrial, inducen la producción de RLO y RLN que, a su vez, originan daños en el ADN, interrumpen sus vías de reparación y ocasiona inestabilidad genómica.
Al inducir mutaciones de la proteína p53 y estimular la expresión del inhibidor ligando X de la proteína de apoptosis (ILXPA), favorecedor de una rápida mutación irreversible, aumenta la resistencia apoptósica de las células epiteliales y aparecen las células cancerosas.
Por último, estimulan la activación del receptor colinérgico muscarínico (RCM3) y de la molécula de señalización Wnt/β-catenina que conduce a la translocación nuclear de la β-catenina, originador del complejo denominado factor de transcripción, integrado por el factor de células T/factor potenciador linfoide (β-catenina-FCT/FPL), que actúa como un coactivador para expresar un gen que regula la madurez celular denominado c-Myc, y, al incrementarse, favorece el cambio a célula cancerígena desde su fase inicial.
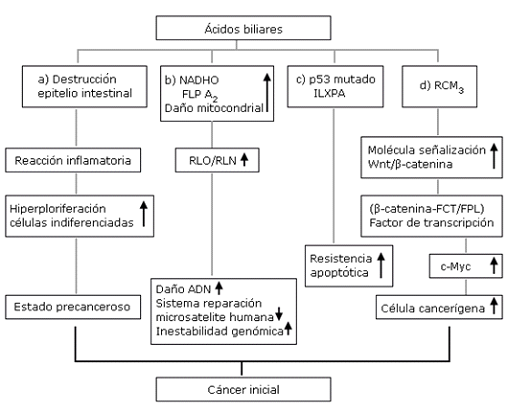 Oxidasa. Fosfolipasa A2 | RLO/RLN: radicales libres de oxígeno/radicales libre de nitrógeno | ILXPA: inhibidor ligado a X de la proteína de apoptosis | RMC3: receptor colinérgico muscarínico 3, β-catenina-FCT/FPL (factor de células T / factor potenciador linfoide | c-Myc: gen que regula la madurez celular.
Oxidasa. Fosfolipasa A2 | RLO/RLN: radicales libres de oxígeno/radicales libre de nitrógeno | ILXPA: inhibidor ligado a X de la proteína de apoptosis | RMC3: receptor colinérgico muscarínico 3, β-catenina-FCT/FPL (factor de células T / factor potenciador linfoide | c-Myc: gen que regula la madurez celular.Fig. 2- Mecanismo de carcinogénesis desencadenado por niveles altos de ácidos biliares en la luz intestinal.

