










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCuban Journal of Agricultural Science
versión On-line ISSN 2079-3480
Cuban J. Agric. Sci. vol.53 no.4 Mayabeque oct.-dic. 2019 Epub 05-Dic-2019
CIENCIA DE LOS PASTOS
Composición mineral de Cenchrus purpureus vc. Cuba CT-115 después del pastoreo utilizado como banco de biomasa
1Instituto de Ciencia Animal, Aptdo. 24 San José de la Lajas, Mayabeque, Cuba
El objetivo del presente trabajo fue estudiar la composición de algunos nutrientes minerales de Cenchrus purpureus vc. Cuba CT-115 después del pastoreo cuando se utiliza como banco de biomasa. Para ello se utilizó un diseño de muestreo completamente aleatorizado con 15 repeticiones. Los tratamientos consistieron en la edad de rebrote o reposo del pasto a partir del momento de salida de los animales del potrero (tiempo cero) y de acuerdo con la tecnología de banco de biomasa. Los resultados indicaron que los contenidos de cenizas, calcio, fósforo y magnesio tuvieron un comportamiento variable, mientras que el nitrógeno se redujo con la edad de rebrote para todos los ciclos de pastoreo de la tecnología. Los contenidos de cenizas oscilaron entre 3.89 % y 15.41% (P< 0.01), mientras que el calcio y el fósforo estuvieron entre 0.30 y 0.83 % (P< 0.05) y entre 0.17 y 0.43% (P< 0.05), respectivamente. Todos los indicadores estudiados tuvieron valores superiores en hojas que en tallos tanto para los hijos basales como para el residuo con su nuevo crecimiento. Los contenidos de fósforo y magnesio encontrados tanto en hojas como en tallos para algunas edades de rebrote fueron inferiores a los requeridos para el adecuado crecimiento y desarrollo de la gramínea, por lo que podría ser necesaria la utilización de fertilización de mantenimiento. Se recomienda usar estos resultados para diseñar otras opciones de manejo.
Palabras-clave: Cuba CT-115; cenizas; nitrógeno; calcio y fósforo
INTRODUCCIÓN
El Cenchrus purpureus vc. Cuba CT-115 se utiliza ampliamente en Cuba por sus características favorables de crecimiento, entre las que se encuentran: menor resistencia al corte, mayor cantidad de hojas, menor altura que el King grass y menor distancia entrenudos en la medida que avanza la edad. Por tal motivo, ofrece mejores posibilidades para su cosecha como banco de biomasa, incluyendo el pastoreo (Crespo y Martínez 2016).
La composición mineral del pasto tiene gran importancia desde el punto de vista fisiológico ya que, es clave para establecer sistemas eficientes de nutrición y manejo de la planta (Bloom y Smith 2014 y Rogóz y Tabak 2017). Además constituye una fuente importante para el suministro básico de los elementos minerales, que son necesarios para la alimentación animal (French 2017 y Villalobos y Sánchez 2018).
Teniendo en cuenta estos aspectos el objetivo del trabajo fue estudiar la composición de algunos nutrientes minerales del C. purpureus vc. Cuba CT-115 cuando se utiliza como banco de biomasa.
MATERIALES Y MÉTODOS
Ubicación, clima y suelo. La investigación se inició con la determinación de la homogeneidad del área y la forma de realizar el muestreo que permitiera obtener resultados confiables, precisos, repetibles y representativos (Fortes et al. 2007). El estudio se realizó en la vaquería B del Instituto de Ciencia Animal (ICA), San José de las Lajas, Mayabeque, ubicada entre los 22º 53 LN y los 82º 02 LO, a 80 msnm (Anon 1989); en un cuartón previamente establecido de Cenchrus purpureus vc. Cuba CT-115.
El suelo se clasificó como Pardo grisáceo mullido según Hernández et al. (2015). Antes de comenzar la etapa experimental se tomaron 10 muestras compuestas de suelo en las diagonales del largo del cuartón, entre 0 y 15 cm de profundidad, con una barrena helicoidal. Se secaron al aire, se homogeneizaron en un mortero y se pasaron por un tamiz con malla de 0,5 mm. Se envasaron en frascos de cristal con cierre hermético y se almacenaron a temperatura ambiente hasta su análisis. Se determinó la materia orgánica (Walkley y Black, citado por Jackson 1970), el nitrógeno (AOAC 2016), el fósforo (Oniani 1964), el calcio y el magnesio (Maslova, citado por Paneque 1965).
La composición química del suelo aparece en la tabla 1, los datos que se presentan son las medias de las muestras compuestas que se tomaron. El pH fue ligeramente ácido y los contenidos de N y K fueron relativamente bajos (Fortes 2013).
Table 1 Chemical composition of soil of the experimental area
| % | mg.100g -1 | pH | ||||
|---|---|---|---|---|---|---|
| N | OM | Ca | Mg | P | K | |
| 0.19 | 3.20 | 2.53 | 0.26 | 2.15 | 5.44 | 5.87 |
En la figura 1 aparece el acumulado de precipitaciones y la temperatura promedio mensual, mínima y máxima durante el año de experimentación, datos que fueron tomados de la Estación Meteorológica del ICA.

Figure 1 Performance of the accumulate of precipitations and temperatures during the experimental period
Tratamientos y diseño. Los tratamientos consistieron en las edades de rebrote o reposo del pasto a partir del momento de salida de los animales del potrero (tiempo cero) como sigue: 0, 15, 30, 45, 60, 75 y 90 días de rebrote del ciclo uno de pastoreo; 0, 15, 30, 45 y 60 días de rebrote del ciclo dos; 0, 15, 30 y 45 días de rebrote del ciclo tres de pastoreo y 0, 15, 30, 45, 60, 75, 90 y 105 días de rebrote del ciclo cuatro de pastoreo (tabla 2); y de acuerdo con la tecnología de banco de biomasa (Martínez y Herrera 2006). Para la toma de muestras se realizó un estudio de homogeneidad del área seleccionada y se determinó el tamaño de muestras adecuado para el estudio morfofisiológico de Cenchrus purpureus vc. Cuba CT-115 en pastoreo. Los resultados mostraron que el área resultó ser homogénea, y se determinó que 15 muestras (macollas como unidades experimentales) son suficientes para indicar con precisión el comportamiento de los indicadores en la población estudiada (Fortes et al. 2007).
Table 2 Distribution of rest cycles during the experiment
| Rest cycles | Date | Duration, days |
|---|---|---|
| Cycle 1 | December 2006- March 2007 | 90 |
| Cycle 2 | March-May 2007 | 60 |
| Cycle 3 | May-July 2007 | 45 |
| Cycle 4 | August- November 2007 | 105 |
Procedimiento. Se utilizó un cuartón de 0.75 ha integrado a la tecnología de bancos de biomasa (Martínez y Herrera 2006). Se seleccionaron 15 muestras (macollas como unidades experimentales) a las edades de rebrote antes mencionadas para cada ciclo de pastoreo. Las macollas se separaron siempre en residuo (rechazo con su nuevo crecimiento) y rebrote (hijos basales). Luego se fraccionaron en hoja, tallo y material muerto (MM) del residuo y hoja y tallo del rebrote.
Para el análisis químico las muestras fueron secadas a 60 °C en estufa de circulación de aire marca Memmert y molidas en un molino de martillo (Culatte typs MFC) con tamiz de 1 mm de diámetro, se envasaron en frascos de cristal herméticamente cerrados y se almacenaron a temperatura ambiente hasta su procesamiento.
Se determinaron los contenidos de cenizas, nitrógeno (N), calcio (Ca), fósforo (P) y magnesio (Mg) según la metodología descrita por la AOAC (2016).
Análisis estadístico. Se efectuó análisis de varianza según diseño completamente aleatorizado y se empleó la dócima de Duncan (1955) para la comparación de las medias en los casos necesarios, todos los análisis se realizaron por duplicado por tratamiento. Para el procesamiento de los datos se utilizó el paquete estadístico IBM-SPSS, Versión 22 (2013).
RESULTADOS Y DISCUSIÓN
En la tabla 3 aparecen los contenidos de cenizas para todos los ciclos de pastoreo durante el año de evaluación. Se observaron diferencias (P<0.01) entre edades de rebrote tanto para las hojas como los tallos de los hijos basales y del residuo; excepto en las hojas de los hijos basales del ciclo tres de pastoreo. Las cenizas de las hojas generalmente fueron superiores a los tallos, y los valores oscilaron entre 3.89 % y 15.41%. Herrera et al. (2002) al estudiar este indicador en el CT-115 en condiciones de pastoreo encontraron mayores contenidos de cenizas en las hojas que los tallos y variaron entre 8-9% y desde 4.5 hasta 7%, respectivamente. Por su parte, Valenciaga et al. (2001) encontraron valores inferiores en las hojas de 4.01% y superiores en el tallo de 7.04%, lo cual podría deberse a las diferentes condiciones de manejo utilizadas.
Por otra parte, Valenciaga et al. (2009) encontraron valores de cenizas en C. purpureus vc. Cuba CT-115 que se incrementaron con la edad, hasta 12.64% para 140 días de rebrote. Sin embargo, en este trabajo los contenidos fueron superiores a los reportados por estos autores y el comportamiento con la edad fue variable. Por su parte, Correa et al. (2016) obtuvieron en pasto kikuyo mayores contenidos de cenizas a los 45 días que a los 80 días de rebrote; mientras que Kozloski et al. (2005) en muestras de C. purpureus vc. Mott cosechado entre 30 y 90 días de rebrote no encontraron grandes diferencias en este indicador.
Table 3 Contents of ash (%) in Cenchrus purpureus cv. Cuba CT-115
| Cycle 1 (December-March) | ||||
|---|---|---|---|---|
| Age, Days | Tillers | Residue | ||
| Ash leaf | Ash stem | Ash leaf | Ash stem | |
| 0 | - | - | 12.97ab | 4.02ab |
| 15 | 9.57ª | 5.10ª | 12.34ª | 3.89ª |
| 30 | 8.94ab | 8.62bc | 12.29ª | 4.94ab |
| 45 | 8.76ab | 8.16bc | 12.37ª | 4.45ab |
| 60 | 8.44b | 8.94c | 12.50ª | 4.89ab |
| 75 | 8.56b | 8.30c | 14.21b | 5.25b |
| 90 | 8.52b | 6.95ab | 12.77ª | 4.69ab |
| SE±/Sign. | 0.23** | 0.28*** | 0.35*** | 0.21*** |
| Cycle 2 (March-May) | ||||
| 0 | - | - | 11.33ª | 6.35ª |
| 15 | - | - | 11.83ab | 6.32ª |
| 30 | 10.52ª | 5.02ª | 12.49bc | 5.43b |
| 45 | 10.25ab | 9.34b | 12.78c | 5.50b |
| 60 | 8.96b | 10.34b | 11.96abc | 5.67ab |
| SE±/Sign. | 0.14*** | 0.20*** | 0.18** | 0.17** |
| Cycle 3 (May-July) | ||||
| 0 | - | - | - | 4.47ª |
| 15 | 12.05 | 5.37ª | 13.77ª | 5.00ab |
| 30 | 12.33 | 6.17ª | 14.03ab | 6.11bc |
| 45 | 11.89 | 8.27b | 15.12bc | 6.25c |
| 60 | 11.25 | 7.65b | 15.35c | 6.53c |
| SE±/Sign. | 0.34 | 0.30** | 0.23** | 0.25** |
| Cycle 4 (August-November) | ||||
| 0 | - | - | 15.41ª | 5.48abc |
| 15 | 11.52ab | 5.85ab | 14.27ab | 6.71ª |
| 30 | 12.08b | 8.13bc | 13.85ab | 6.16ab |
| 45 | 11.53ab | 9.25c | 13.13b | 6.64ª |
| 60 | 12.30b | 7.07abc | 14.67ab | 4.58bc |
| 75 | 10.17ª | 4.34ª | 13.91ab | 4.11c |
| 90 | 11.15ab | 5.65ab | 12.91b | 4.91abc |
| 105 | 11.16ab | 5.32ab | 13.50ab | 4.65bc |
| EE±/Sign. | 0.25*** | 0.43*** | 0.32*** | 0.26*** |
abcValues with different letters per column differ at P <0.05 (Duncan 1955)
***P <0.001 ** P <0.01
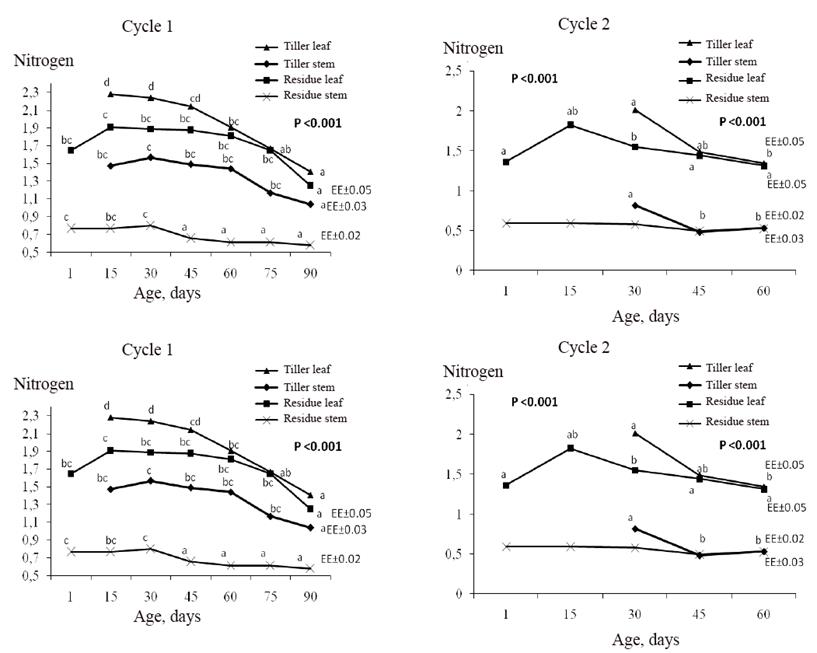
En la figura 2 aparecen los contenidos de nitrógeno para todos los ciclos de pastoreo estudiados. El nitrógeno tuvo mayor valor en hojas que en tallos tanto en hijos basales como en el residuo. El contenido de nitrógeno tuvo una tendencia de reducción en la planta con la edad.
Esto ocurre porque este nutriente es de mayor importancia en las partes más jóvenes. Su absorción es cada vez menor cuando fisiológicamente la planta envejece o llega a su estado de madurez. Gándara et al. (2017) también encontró reducciones del nitrógeno y la proteína con la edad del pasto. En este sentido, Pérez et al. (2004) afirman que el contenido de nitrógeno en las plantas disminuye durante el crecimiento, y presenta una alta correlación con la acumulación de materia seca, más que otros indicadores y para cualquier estadio de crecimiento y edad de rebrote, estos autores encontraron reducciones de los contenidos de N en hojas de Brachiaria híbrido en la medida que avanzó la edad de la planta.
En la tabla 4 aparecen los contenidos de calcio en los cuatro ciclos de pastoreo de la tecnología. En todos los casos se observaron diferencias (P<0.05) con la edad de rebrote, tanto en las hojas como en tallos, excepto en la hoja del residuo del ciclo uno de pastoreo, los valores oscilaron entre 0.30 y 0.83 % de la MS. A pesar de las diferencias significativas encontradas entre edades, la variación entre ellas estuvo en un rango estrecho. En este sentido, Valenciaga et al. (2009) no encontraron diferencias entre edades de rebrote para este indicador en el Cuba CT-115.Los contenidos de calcio siempre fueron superiores en hojas a 0.30 % de la MS, nivel establecido por Cunha y Mc Dowell (2012) como crítico para la alimentación animal. Resultados similares fueron reportados por Clavero et al. (1994).
Los valores de los contenidos de calcio encontrados en hoja siempre fueron superiores a los tallos. Resultados similares a estos fueron reportados por Herrera et al. (2008) para todas las variedades de Cenchrus estudiadas incluida la de este estudio. Dicho comportamiento resulta contradictorio si se tiene en cuenta que el calcio desempeña un importante papel como elemento cementante en la pared celular y es precisamente en las hojas, donde se encuentra el menor contenido de pared celular.
Table 4 Calcium content (%) in leaves and stems of Cenchrus purpureus cv. Cuba CT-115
| Cycle 1 (December-March) | ||||
|---|---|---|---|---|
| Age, days | Tillers | Residue | ||
| Ca Leaf | Ca Stem | Ca Leaf | Ca Stem | |
| 0 | - | - | 0.79 | 0.50ª |
| 15 | 0.65ab | 0.47ª | 0.82 | 0.54ab |
| 30 | 0.69bc | 0.57b | 0.82 | 0.62c |
| 45 | 0.70c | 0.54b | 0.83 | 0.58bc |
| 60 | 0.64ª | 0.58b | 0.82 | 0.60bc |
| 75 | 0.65ab | 0.53b | 0.79 | 0.64c |
| 90 | 0.64a | 0.57b | 0.82 | 0.65c |
| SE±/Sign. | 0.01*** | 0.01*** | 0.01 | 0.01*** |
| Cycle 2 (March-May) | ||||
| 0 | - | - | 0.41ª | 0.34ª |
| 15 | - | - | 0.42ª | 0.40c |
| 30 | 0.50ª | 0.32ab | 0.43ª | 0.36ab |
| 45 | 0.63ab | 0.29ª | 0.47b | 0.34ª |
| 60 | 0.62b | 0.38b | 0.47b | 0.371bc |
| SE±/Sign. | 0.01*** | 0.01*** | 0.004*** | 0.01** |
| Cycle 3 (May-July) | ||||
| 0 | - | - | - | 0.37ab |
| 15 | 0.63ª | 0.45ª | 0.48ª | 0.39b |
| 30 | 0.63ª | 0.30b | 0.46ª | 0.34ab |
| 45 | 0.57b | 0.31b | 0.35b | 0.31ª |
| 60 | 0.49c | 0.35b | 0.40b | 0.36ab |
| SE±/Sign. | 0.01*** | 0.01** | 0.01** | 0.01*** |
| Cycle 4 (August-November) | ||||
| 0 | - | - | 0.60ab | 0.38ª |
| 15 | 0.52ª | 0.35ab | 0.62b | 0.36ab |
| 30 | 0.54ª | 0.37b | 0.62b | 0.37ª |
| 45 | 0.55ab | 0.37b | 0.63b | 0.36ab |
| 60 | 0.51ª | 0.35b | 0.58ª | 0.30bc |
| 75 | 0.61b | 0.31ª | 0.63b | 0.30c |
| 90 | 0.54ª | 0.32ab | 0.63b | 0.33abc |
| 105 | 0.52a | 0.33ab | 0.62b | 0.31bc |
| SE±/Sign. | 0.01*** | 0.01** | 0.01* | 0.01** |
abc Values with different letters per column differ at P <0.05 (Duncan 1955)
***P <0.001 **P <0.01 * P <0.05
Para todos los ciclos de pastoreo los contenidos de fósforo (tabla 5) mostraron diferencias (P<0.05) de este indicador con la edad de rebrote. En las hojas, los contenidos de fósforo mostraron una reducción (P <0.01) con la edad de la planta, excepto para el ciclo uno de pastoreo, los valores oscilaron entre 0.17 y 0.43% de la MS. En el caso de los tallos, no hubo una tendencia fija a incrementarse o reducirse con la edad sino que en todos los casos los tenores fueron variables y oscilaron entre 0.15 y 0.26% de la MS. En la literatura se reportan reducciones de este elemento con la edad de la planta (Santiago et al. 2016), sin embargo en este trabajo el comportamiento fue variable.
Table 5 Phosphorus content (%) in leaves and stems of Cenchrus purpureus cv. Cuba CT-115
| Cycle 1 (December-March) | ||||
|---|---|---|---|---|
| Age, days | Tillers | Residue | ||
| P Leaf | P Stem | P Leaf | P Stem | |
| 0 | - | - | 0.24abc | 0.19ª |
| 15 | 0.24ab | 0.19ª | 0.26c | 0.23ab |
| 30 | 0.21ª | 0.19ª | 0.21ª | 0.21ab |
| 45 | 0.21ª | 0.21ab | 0.23ab | 0.21ab |
| 60 | 0.26b | 0.25c | 0.25bcd | 0.24b |
| 75 | 0.23ª | 0.25bc | 0.24abc | 0.22ab |
| 90 | 0.23ab | 0.24bc | 0.25bc | 0.21ab |
| SE±/Sign. | 0.01*** | 0.01*** | 0.01*** | 0.01*** |
| Cycle 2 (March-May) | ||||
| 0 | - | - | 0.33ª | 0.29ª |
| 15 | - | - | 0.28c | 0.17b |
| 30 | 0.35ª | 0.25ª | 0.29c | 0.24ab |
| 45 | 0.27b | 0.26ª | 0.32ab | 0.23ab |
| 60 | 0.25b | 0.22b | 0.30bc | 0.24b |
| SE±/Sign. | 0.01*** | 0.01* | 0.01** | 0.01** |
| Cycle 3 (May-July) | ||||
| 0 | - | - | - | 0.19ab |
| 15 | 0.32ª | 0.20ª | 0.43ª | 0.19ab |
| 30 | 0.33ª | 0.22ª | 0.34ab | 0.21b |
| 45 | 0.27b | 0.20ª | 0.26bc | 0.18ª |
| 60 | 0.24b | 0.24b | 0.24c | 0.20ab |
| SE±/Sign. | 0.01** | 0.01** | 0.01*** | 0.01** |
| Cycle 4 (August-November) | ||||
| 0 | - | - | 0.23ab | 0.20ab |
| 15 | 0.24ª | 0.21ª | 0.25ª | 0.15ª |
| 30 | 0.23ab | 0.18ab | 0.23ab | 0.18ab |
| 45 | 0.23ab | 0.16b | 0.21abc | 0.19ab |
| 60 | 0.23ab | 0.20ª | 0.22abc | 0.21b |
| 75 | 0.20b | 0.16b | 0.19bc | 0.17ab |
| 90 | 0.21ab | 0.19b | 0.17c | 0.18ab |
| 105 | 0.20b | 0.19ab | 0.19bc | 0.20ab |
| SE±/Sign. | 0.01** | 0.01** | 0.01*** | 0.01*** |
abc Values with different letters per column differ at P <0.05 (Duncan 1955)
***P <0.001 **P <0.01 * P <0.05
Las hojas presentaron mayores valores de fósforo que los tallos de forma general. Este comportamiento resulta lógico si tenemos en cuenta las múltiples funciones que desarrolla este elemento en el metabolismo vegetal, ya que forma parte de una amplia gama de moléculas como azúcares fosfatos, ácidos nucleicos, coenzimas, entre otras, además de que controla diferentes procesos metabólicos que ocurren fundamentalmente en las hojas de la planta (Gardner et al. 2017 y Malhotra et al. 2018).
Los requerimientos de fósforo para el óptimo crecimiento de las plantas están en el rango de 0,3 a 0,5% de la materia seca para el periodo de crecimiento vegetativo según Vistoso et al. (2017). Sin embargo, Epstein y Bloom (2005), así como Domínguez et al. (2012) consideran que un contenido superior a 0,2% de la masa seca constituye un nivel tisular adecuado de fósforo en las plantas. Teniendo esto en consideración para algunas edades de rebrote el fósforo estuvo por debajo de los niveles adecuados para el metabolismo por lo que se podría sugerir el uso de fertilización fosfórica al suelo para suplir esta deficiencia en la planta. Para la alimentación animal se considera como nivel crítico valores inferiores a 0,25% de la MS (Cunha y Mc Dowell 2012), en algunos ciclos de pastoreo se encontraron valores inferiores a este, por lo que es posible que se necesite algún tipo de suplementación en los animales para cubrir este déficit.
Los contenidos de magnesio siempre fueron superiores en las hojas que en los tallos (tabla 6). Los cloroplastos de las plantas son ricos en Mg pues la principal función de este elemento, es su rol como átomo central de la molécula de clorofila presente fundamentalmente en las hojas de las plantas (Basantes 2016 y Chen et al. 2018). Los valores estuvieron entre 0.25 y 0.52 % para las hojas y 0.16 y 0.30 % para los tallos. Según Torres (1999) contenidos de 0.2 % de magnesio en la MS se consideran concentraciones aceptables de este elemento para el buen funcionamiento fisiológico de la planta. Por tal motivo, las concentraciones encontradas en el trabajo en algunos casos están por debajo de las necesarias por la planta, aunque la diferencia entre los requerimientos y los contenidos de Mg encontrados es muy baja.
Por su parte, Cunha y Mc Dowell (2012) consideran como nivel crítico para la alimentación animal 0,20% de la MS, por lo que también para algunas edades de la planta los resultados fueron inferiores a este valor.
Table 6 Magnesium content (%) in leaves and stems of C. purpureus cv. Cuba CT-115
| Cycle 1 (December-March) | ||||
|---|---|---|---|---|
| Age, days | Tillers | Residue | ||
| Mg Leaf | Mg Stem | Mg Leaf | Mg Stem | |
| 0 | - | - | 0.40ª | 0.24ª |
| 15 | 0.33bc | 0.30b | 0.47b | 0.22ª |
| 30 | 0.35c | 0.36c | 0.52b | 0.26ab |
| 45 | 0.37c | 0.36c | 0.48b | 0.30b |
| 60 | 0.26ª | 0.29ab | 0.47b | 0.25ª |
| 75 | 0.27ab | 0.25ab | 0.47b | 0.25ª |
| 90 | 0.25a | 0.25a | 0.38a | 0.26ab |
| SE±/Sign. | 0.01*** | 0.01*** | 0.01*** | 0.01*** |
| Cycle 2 (March-May) | ||||
| 0 | - | - | 0.28ab | 0.26bc |
| 15 | - | - | 0.29abc | 0.28c |
| 30 | 0.32ª | 0.28ª | 0.28ª | 0.28c |
| 45 | 0.42ab | 0.22b | 0.30bc | 0.20ª |
| 60 | 0.49b | 0.23b | 0.31c | 0.22ab |
| SE±/Sign. | 0.04* | 0.01** | 0.01* | 0.01** |
| Cycle 3 (May-July) | ||||
| 0 | - | - | - | 0.24ª |
| 15 | 0.50ª | 0.27ab | 0.38ª | 0.25ab |
| 30 | 0.48ª | 0.29b | 0.39ª | 0.26ab |
| 45 | 0.29b | 0.29b | 0.29b | 0.26b |
| 60 | 0.30b | 0.25a | 0.30b | 0.24b |
| SE±/Sign. | 0.01*** | 0.01** | 0.01*** | 0.01* |
| Cycle 4 (August-November) | ||||
| 0 | - | - | 0.29a | 0.23abc |
| 15 | 0.30 a | 0.19ab | 0.36ab | 0.24bc |
| 30 | 0.33 ab | 0.23ab | 0.36ab | 0.22abc |
| 45 | 0.34 ab | 0.25b | 0.37b | 0.25c |
| 60 | 0.32 ab | 0.23ab | 0.34ab | 0.21abc |
| 75 | 0.34 ab | 0.25b | 0.36ab | 0.22abc |
| 90 | 0.35 ab | 0.16a | 0.37b | 0.17ab |
| 105 | 0.37 b | 0.16a | 0.38b | 0.16a |
| SE±/Sign. | 0.01*** | 0.01*** | 0.01*** | 0.02** |
abc Values with different letters per column differ at P <0.05 (Duncan 1955)
***P <0.001 **P <0.01 *P <0.05
Se concluye que los contenidos de cenizas, calcio, fósforo y magnesio tuvieron un comportamiento variable con la edad de rebrote para todos los ciclos de pastoreo de la tecnología de banco de biomasa. El contenido de nitrógeno se redujo con la edad de rebrote en todas las fracciones de la planta estudiadas. Los indicadores estudiados tuvieron valores superiores en hojas que en tallos tanto para los hijos basales como para el residuo con su nuevo crecimiento. Los contenidos de fósforo y magnesio encontrados tanto en hojas como en tallos para algunas edades de rebrote fueron inferiores a los requeridas para el adecuado crecimiento y desarrollo de la gramínea, por lo que podría ser necesaria la utilización de fertilización de mantenimiento. Se recomienda usar estos resultados para diseñar otras opciones de manejo.
REFERENCES
Anon. 1989. Atlas Nacional de Cuba. (Ed). Instituto de Geografía de la Academia de Ciencias de Cuba e Instituto Cubano de Geodesia y Cartografía. Impreso por el Instituto Geográfico Nacional de España. [ Links ]
AOAC. 2016. Official methods of analysis of AOAC International. 20th ed., Rockville, MD: AOAC International, ISBN: 978-0-935584-87-5, Available: Available: http://www.directtextbook.com/isbn/9780935584875 , [Consulted: October 13, 2017]. [ Links ]
Basantes, E.R. 2016. Silvicultura y fisiología vegetal aplicada. David Andrade Aguirre (Ed.). Comisión Editorial de la Universidad de las Fuerzas Armadas ESPE. Primera edición. ISBN: 978-9978-301-36-4. 439 p. [ Links ]
Bloom, A.J. & Smith, S. 2014. Mineral Nutrition. In: Plant Physiology and Development. Sixth Edition. Lincoln Taiz, Eduardo Zeiger, Ian Max Moller and Angus Murphy (Eds). Sinauer Associates Inc., Sunderland, MA. ISBN: 978-1-60535-255-8. 761p. [ Links ]
Chen, Z. C., Peng, W. T., Li, J., & Liao, H. 2018. Functional dissection and transport mechanism of magnesium in plants. Seminars in Cell & Developmental Biology, 74(1): 142-152. doi:10.1016/j.semcdb.2017.08.005 [ Links ]
Clavero, T., Ferrer, O. & Pérez, J. 1994. Contenido mineral del pasto elefante enano (Cenchrus purpureus cv Mott.) bajo diferentes condiciones de defoliación. Rev. Fac. Agron. (LUZ). 11: 355-364. ISSN 0378-7818. [ Links ]
Correa, H.J., Jaimes, L.J., Avellaneda, J.H., Pabón, M.L. & Carulla, J.E. 2016. Efecto de la edad de rebrote del pasto kikuyo (Pennisetum clandestinum) sobre la producción, la calidad de la leche y el balance de nitrógeno en vacas Holstein. Livestock Research for Rural Development 28 (3). ISSN: 0121-3784. [ Links ]
Crespo, G & Martínez, R.O. 2016. Study of the chemical soil fertility in the biomass bank technology of Cenchrus purpureus Schum cv. CUBA CT-115 with different exploitation years. Cuban Journal of Agricultural Science. 50th Anniversary. 50(2): 497. [ Links ]
Cunha, T. J. and McDowell, L. R. 2012. Nutrition of Ruminants in warm climates. Academic Press, New York. 443p. [ Links ]
Domínguez, T.G., R.G. Ramírez, A.E. Estrada, L.M. Scott, H. González, & M.D.S. Alvarado. 2012. Importancia nutrimental en plantas forrajeras del matorral espinoso tamaulipeco. Ciencia UANL 15(59):77-93. ISSN 2007-1175. [ Links ]
Duncan, D.B. 1955. Multiple range and multiple F test. Biometrics 11(1): 1-42 [ Links ]
Epstein, E. & Bloom, A.J., 2005. Mineral Nutrition of plants: principles and perspectives, 2nd ed. Sinauer Associates, Inc., Publishers, Sunderland. ISBN 0878931724. [ Links ]
Fortes, D. 2013. Comportamiento de algunos indicadores morfofisiológicos y de calidad de Cenchrus purpureus vc. Cuba CT-115 utilizado como banco de biomasa. PhD Thesis. UNAH, Mayabeque, Cuba. 100 p. [ Links ]
Fortes, D., Herrera, R.S., Torres, V., García, M., Cruz, A.M., Romero, A., Noda, A. & González, S. 2007. Determination of a sampling method for the morphophysiological study of grazing Cenchrus purpureus cv. Cuba CT-115. Cuban Journal of Agricultural Science , 41(4): 359-364. [ Links ]
French, K.E. 2017. Species composition determines forage quality and medicinal value of high diversity grasslands in lowland England. Agriculture, Ecosystems and Environment. 241: 193-204. http://dx.doi.org/10.1016/j.agee.2017.03.012. [ Links ]
Gándara, L., Borrajo, C.I., Fernández, J.A. & Pereira, M. 2017. Efecto de la fertilización nitrogenada y la edad del rebrote sobre el valor nutritivo de Brachiaria brizantha cv. "Marandú". Revista de la Facultad de Ciencias Agrarias, 49(1): 69. ISSN: 0370-4661. [ Links ]
Gardner, F. P., Pearce, R. B. & Mitchell, R. L. 2017. Physiology of crop plants. F. P. Gardner, R. B. and R. L., Mitchell.(Eds) Second Edition. 327 pp. ISBN: 9788172336622. [ Links ]
Hernández, J. A.; Pérez, J. J. M.; Bosch, I. D. & Castro, S. N. 2015. Clasificación de los suelos de Cuba. Ed. Ediciones INCA, Mayabeque, Cuba. ISBN 978-959-7023-77-7. 93p. [ Links ]
Herrera, R.S., Martínez, R.O., Tuero, R., García, M. & Cruz, A.M. 2002. Movement of substances during grazing and regrowth of the clone CUBA CT-115 (Cenchrus purpureus sp). Cuban Journal of Agricultural Science, 36 (4): 403-407. [ Links ]
Herrera, R.S., Fortes, D., García, M., Cruz, A.M. & Romero, A. 2008. Study of the mineral composition in varieties of Cenchrus purpureus. Cuban Journal of Agricultural Science, 42(4): 393-398. [ Links ]
IBM-SPSS. 2013. Software for Window, IBM® SPSS Statistics, version 22.0, INC., Chicago, IL USA. [ Links ]
Kozloski, G. V., Perottoni, J. & Sanchez, L. M. B. 2005. Influence of regrowth age on the nutritive value of dwarf elephant grass hay (Cenchrus purpureus Schum. cv. Mott) consumed by lambs. Animal Feed Science and Technology 119(1-2):1-11. https://doi.org/10.1016/j.anifeedsci.2019.03.006. [ Links ]
Malhotra, H., Vandana Sharma, S., & Pandey, R. 2018. Phosphorus Nutrition: Plant Growth in Response to Deficiency and Excess. In: Plant Nutrients and Abiotic Stress Tolerance. Hasanuzzaman, M., Fujita, M., Oku, H., Nahar, K. y Hawrylak-Nowak, B. (Eds). Springer, Singapore. p. 171-190. doi:10.1007/978-981-10-9044-8_7. ISBN: 978-981-10-9043-1. [ Links ]
Martínez, R.O. & Herrera, R.S. 2006. Empleo del Cuba CT-115 para solucionar el déficit de alimentos durante la seca. En: Producción y manejo de los recursos forrajeros tropicales. Eds. M.E. Velasco, A. Hernández, R.A. Perezgrovas y B. Sánchez. Universidad Autónoma de Chiapas, p 75-97. [ Links ]
Oniani, O. G. 1964. Determinación del fósforo y potasio del suelo en una misma solución de suelo Krasnozen y Podsólicos en Georgia. Agrojima 6:25. [ Links ]
Paneque, V. 1965. Manual de prácticas de suelo. Universidad de la Habana. p. 25. [ Links ]
Pérez, J.A., García, E., Enríquez, J.F., Quero, A.R., Pérez, J. & Hernández, A. 2004. Análisis de crecimiento, área foliar específica y concentración de nitrógeno en hojas de pasto “mulato” (Brachiaria híbrido, cv.). Téc Pecuaria Méx. 42: 447. [ Links ]
Rogóż, A. & Tabak, M. 2017. Content of macroelements in pasture sward and their effect on the fodder value. In: VIII International Scientific Conference. Toxic Substances in the Environment. Edited by: Tomáš Lošák, Monika Tabak, Dawid Tabak, Jacek Antonkiewicz. Krakow, Poland, 14-15 September 2017. ISBN: 978-83-948965-0-8. [ Links ]
Santiago, I., Lara, A., Miranda, L.A., Huerta, M., Krishnamurthy, L. & Muñoz-González, J.C. 2016. Chemical and mineral composition of leucaena associated with star grass during the rainy season. Revista Mexicana de Ciencias Agrícolas Pub. Esp. Núm. 16 p. 3173-3183. DOI: https://doi.org/10.29312/remexca.v0i16.400. [ Links ]
Torres, A. 1999. Consideraciones sobre la fisiología de la nutrición mineral en las plantas superiores. Universidad Agraria de La Habana, Facultad de Agronomía. [ Links ]
Valenciaga, D., Chongo, B. & La O, O. 2001. Characterization of Pennisetum CUBA CT-115 clone. Chemical composition and rumen DM degradability. Cuban Journal of Agricultural Science, Volume 35(4): 325-330. [ Links ]
Valenciaga, D., Chongo, B., Herrera, R.S., Torres, V., Oramas, A., Cairo, J.G. and Herrera, M. 2009. Effect of regrowth age on the chemical composition of Cenchrus purpureus cv. CUBA CT-115. Cuban Journal of Agricultural Science , 43(1): 71-76. [ Links ]
Villalobos, L. & Sánchez, J. M. 2018. Contenido macro y micromineral del pasto Ryegrass (Lolium spp.) en la zona alta de Cartago, Costa Rica. Nutrición Animal Tropical. 12(2):1-19.ISSN:2215-3527 / DOI: https://doi.org/10.15517/nat.v12i2.34927 [ Links ]
Vistoso G., Erika, Sandaña, P. & Iraira, S. 2017. Fertilización fosfatada de praderas en suelos Trumaos de la Región de Los Lagos. Erika Vistoso, Patricio Sandaña y Sergio Iraira (Eds.) Colección de Libros INIA Nº 37. Instituto de Investigaciones Agropecuarias, Centro Regional de Investigación Remehue, Osorno, Chile. ISSN: 0717-4713.124 p. [ Links ]
Recibido: 21 de Enero de 2019; Aprobado: 04 de Febrero de 2019

