










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Las razas o ecotipos criollos existentes en el continente americano suelen ser muy diversos y aunque todos son denominados “criollos”, presuntamente tienen el mismo origen genético a partir de la península ibérica. Posteriormente se fueron desarrollando bajo diferentes condiciones de manejo o sistemas de producción y condiciones climáticas.
Estas razas que son de origen Bos taurus, y el resultado de la evolución y adaptación (Guintard, 2016) desde hace más de 500 años, ha demostrado que su origen está relacionado con las razas españolas Rubia Gallega, Asturiana y Andaluza, aunque en Cuba se ha identificado también alguna influencia de animales Cebú (Lirón et al., 2011; Uffo et al., 2012). Estas razas se han considerado como localmente adaptadas y se consideran rústicas. Todo esto que se puede considerar válido debe ser examinado a la luz de las condiciones actuales y en el caso particular del Criollo cubano en la localidad donde se encuentra el rebaño más importante, Suárez Tronco et al., (2021) demostraron un empeoramiento de las condiciones climáticas a través del tiempo, así como de los indicadores de las pruebas de comportamiento, estando ambos aspectos relacionados. Además, Suárez Tronco et al. (2022 a y b) concluyeron que aun cuando se reconoce el mérito adaptativo de la misma al clima del país su comportamiento pre destete está afectado por el estrés calórico y se demostró la existencia de una importante variabilidad genética en la forma de respuesta a esta importante característica.
Por lo planteado anteriormente nos trazamos como objetivos en este trabajo valorar el peso final en prueba de comportamiento bajo diferentes condiciones, la estimación de los parámetros genéticos y la posible existencia de interacción genotipo ambiente.
MATERIALES Y MÉTODOS
Las condiciones de tenencia de los animales fueron descritas por Suárez Tronco et al., (2021), así como los rasgos estudiados. Se utilizó la información referente a 1 532 pedigrís, de 2 772 animales machos y hembras diferentes, hijos de 72 sementales y 1 193 madres que nacieron entre los años 1981 y 2016 de la Empresa Genética y Cría “Manuel Fajardo” y se utilizó el peso a 18 meses de edad que es el peso final de la prueba.
Primero la información se dividió en dos conjuntos de datos, aquellos que se encontraban por encima de la media para el carácter estudiado (peso final en prueba de comportamiento) y los que estaban por abajo, que se denominaron peso alto (PA) y peso bajo (PB) y se consideraron como dos caracteres distintos para contrastar solamente los valores extremos. Un procedimiento similar ya fue utilizado por Rodríguez y Guerra (2013) en ganado Cebú, pero utilizando la ganancia y Suárez Tronco et al. (2020) en ganado Santa Gertrudis. Se depuró la base de datos eliminando aquellos sementales con menos de 5 hijos y grupos contemporáneos menores de tres. La Tabla 1 muestra una descripción de cada subconjunto de datos con la información fundamental.
Tabla 1 Composición genética para los dos ambientes
| Ambiente | n | No. GC | No. padres | No. madres |
|---|---|---|---|---|
| Alto | 1 386 | 80 | 73 | 658 |
| Bajo | 1 386 | 80 | 64 | 665 |
n= número de animales; GC Grupo Contemporáneo; No. Padres y No. madres
Posteriormente la propia base de datos se subdividió en 3 partes: animales evaluados hasta 1990, entre 1991 a 2000 y posterior al 2000, para considerar el comportamiento de los animales a través del tiempo donde ha existido cambios en el manejo, sistema de producción, etc. (Tabla 2).
Tabla 2 Composición genética para los 3 periodos de tiempo
| Ambiente | n | No. GC | No. padres | No. madres |
|---|---|---|---|---|
| Hasta 1990 | 323 | 22 | 72 | 262 |
| 1991-2000 | 371 | 29 | 72 | 277 |
| >2001 | 752 | 49 | 72 | 574 |
n= número de animales; GC Grupo Contemporáneo; No. Padres y No. madres
En el primer caso se utilizó un modelo animal bicarácter y para los periodos de tiempo multicarácter. De forma general en función del número de caracteres el modelo planteado es el siguiente:
Rasgo 1 y1= X1 β1 + Z1 a1 + e1
Rasgo 2 y2= X2 β2 + Z2 a2 + e2
Donde:
yi = vector de las observaciones para el i-ésimo rasgo.
βi = vector de los efectos fijos para el i-ésimo rasgo (grupo contemporáneo -rebaño, año-época-, regresión lineal de la edad final en el peso final y sexo.
ai = vector de los efectos aleatorios del animal para el i-ésimo rasgo.
ei = vector de los efectos residuales aleatorios para el i-ésimo rasgo.
Xi y Zi son matrices de incidencia que relacionan el vector de observaciones del i-ésimo rasgo con los efectos fijos y aleatorios (animal), respectivamente.
El modelo se puede ajustar a la estructura de matrices, y si los individuos se ordenan dentro de
características, el vector de observaciones (y) sería el siguiente:
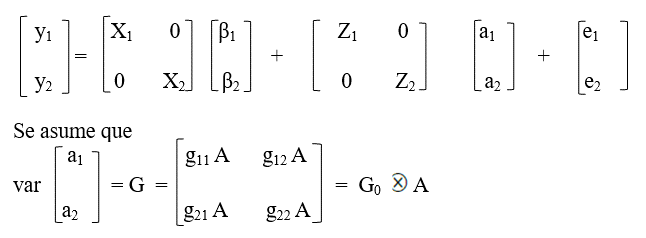
Donde:
g ij = ij-ésimo elemento de la matriz G0, lo que representa las varianzas y covarianzas para los efectos aleatorios del animal, con cada elemento definido como:
g11 = varianza genética aditiva directa para yi
g22 = varianza genética aditiva directa para y2
g12 y g21 = covarianza genética aditiva entre ambos caracteres;
A = matriz de relaciones genéticas aditivas de tamaño n x n;
= producto directo
Y
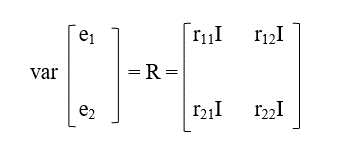
rij = elementos de la matriz de varianzas y covarianzas (R0) para los efectos residuales
I = matriz identidad de tamaño n x n.
Lo anterior se generalizó para el multicarácter para 3 rasgos.
En todos los casos se estimaron los componentes de varianza y covarianza, así como los parámetros genéticos, heredabilidades (h2) y correlaciones genéticas. Se utilizó el procedimiento de máxima verosimilitud restringida (REML) y el algoritmo libre de derivadas usando el software MTDFREML (Boldman, et al. 1995).
Se calcularon las correlaciones de Pearson y Spearman entre los valores genéticos de los padres evaluados en los diferentes ambientes (alto y bajo, así como entre los periodos de tiempo) como un posible indicador de la presencia de interacción genotipo-ambiente utilizando el procedimiento PROC CORR del SAS 9.4 (2014).
RESULTADOS Y DISCUSIÓN
En la Tabla 3 se presentan los estadígrafos generales para PA y PB. Hubo mayor variabilidad para PB y las medias generales para ambos grupos de caracteres fueron para PA 349,79 ± 37,17 y para PB 279,16 ± 32,51 kg. En PA los animales pesaron 25,3% más. La muestra fue ligeramente más homogénea con un coeficiente de variación 1% menor.
Tabla 3 Estadígrafos generales para los dos ambientes
| Ambiente | n | Media (Kg) | ± DE | CV (%) |
|---|---|---|---|---|
| Alto (PA) | 766 | 349,76 | 37,17 | 10,63 |
| Bajo (PB) | 766 | 279,16 | 32,51 | 11,65 |
n = animales con datos; ±DE =desviación estándar; CV =coeficiente de variación.
Con una muestra similar de la misma raza y bajo las mismas condiciones Suárez Tronco et al., (2021) encontraron valores más coincidentes con el PA. Resultados de ganado Criollo son escasos. Bajo el nombre de criollos se enuncian diferentes ecotipos que han sufrido un proceso de diferenciación a través del tiempo y no son estrictamente comparables y mucho más si se consideran pruebas de comportamiento, las que también difieren en sus características.
En razas criollas en Colombia en rebaños en conservación como parte del programa de recursos genéticos animales, en la raza Romosinuano, Ossa, Suárez y Pérez (2007) reportaron para peso a los 16 meses, 247,18 ± 33,81 kg y Martínez, Onofre y Polanco (2009) en animales criollos Sanmartinero reportaron valores medios de 222,58 ± 39,6 kg, todos inferiores a los resultados en PB, pero con menor edad. En ambos casos la variabilidad de los datos fue superior (13,67 y 17,79%, respectivamente).
En la Tabla 4 aparecen los resultados obtenidos en el análisis bicarácter considerando los dos conjuntos de datos como rasgos independientes según lo planteado por Falconer y Mackay (2006) como una vía para determinar posible interacción genotipo-ambiente.
La heredabilidad (h2) para peso final alto fue casi cuatro veces mayor que en el ambiente bajo, aparentemente por una mayor varianza aditiva, lo que corrobora la necesidad de que los animales expresen sus potencialidades genéticas con condiciones adecuadas de tenencia, lo que por demás es un requisito obvio para un sistema de producción eficiente.
Los resultados obtenidos se encuentran en el rango de algunos trabajos en Colombia en la raza Romosinuano de h2 = 0.21 (Ossa et al. 2014), pero valores más altos (h² =0.38) se han encontrado en el criollo Blanco Orejinegro y de h² = 0.37 en Romosinuano (Bejarano, 2016), así como h2 = 0,36 en ganado Costeño con Cuernos (Vergara et al. 2018). En cambio, Wasike et al. (2009), con animales Boran en Kenia, reportaron estimaciones de h² = 0.08 para características de crecimiento a los 18 meses de edad. Más recientemente en Romosinuano también se han reportado valores de h2 bajos (0.13) por López-Martínez, Ossa-Saraz y Santana-Rodríguez (2021).
Tabla 4 Componentes de varianza y covarianza, parámetros genéticos y sus errores estándar para PA y PB
| Estimados | PA | PB | |
|---|---|---|---|
| σ 2 A | 210,59 | 38,74 | |
| σA (AB) | 90,31 | ||
| σ 2 E | 582,79 | 521,17 | |
| σ E(AB) | 527,35 | ||
| σ 2 P | 793,38 | 559,91 | |
| σ P (AB) | 617,66 | ||
| h2 ± ee | 0,26 ± 0,12 | 0,07 ± 0,07 | |
| rg | 0,99 ± 0,08 | ||
| re | 0,96 ± 0,07 | ||
| rp | 0,93 ± 0,06 |
σ 2 A= varianza genética aditiva, σA (AB)= covarianza genética, σ 2 E= varianza residual, σ E(AB)= covarianza residual, σ 2 P= varianza fenotípica, σ P (AB)= covarianza fenotípica, h2 ± ee= heredabilidad y error estándar, rg, re y rp= correlaciones genética, residual y fenotípica, respectivamente. PA y PB= pesos finales en prueba de comportamiento, alto y bajo, respectivamente.
Teniendo en cuenta las diferencias notables entre las heredabilidades las precisiones de los valores genéticos de cada grupo deben haber sido diferentes y por tanto hubiera sido adecuado estimar una correlación ponderada por las precisiones de los VG.
En la figura 1 se presenta la distribución de los valores genéticos de los 72 sementales en el ambiente alto que fue similar a lo encontrado en el ambiente bajo, pero con una menor variabilidad de los datos. Se puede apreciar como 44 sementales de 72, el 61,1% se encuentran con VG entre -2,63 y +3,87 kg. Los rangos en general fueron desde -12,28 a +11,18 kg para PB y desde -28,63 a +26,08 kg para PA. Solo 15 sementales el 20,8% fueron mejoradores. Las correlaciones, tanto entre los VG (0,99) como entre los órdenes de mérito entre ambos ambientes (r=1) no demuestran la existencia de interacción genotipo ambiente.
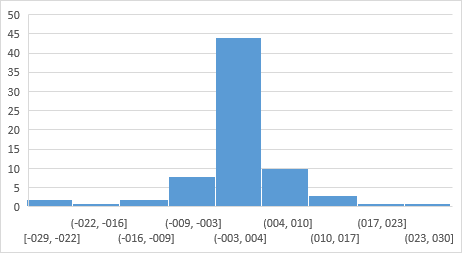
Los estadígrafos generales para las pruebas de comportamiento en los 3 periodos de tiempo analizados se presentan en la tabla 5. Se puede apreciar como después de un buen comportamiento durante los primeros años en que se instauraron las pruebas, estas tocaron fondo en el periodo posterior de crisis de la economía cubana y hay una ligera recuperación posterior.
Tabla 5 Comportamiento de los animales en prueba de comportamiento para peso final
| Año | Animales | Media (kg) | DS (kg) | CV (%) |
|---|---|---|---|---|
| Hasta 1990 | 323 | 414,89 | 61,40 | 14,62 |
| 1991-2000 | 371 | 299,94 | 51,71 | 17,24 |
| Después de 2000 | 752 | 312,13 | 32,39 | 10,38 |
En la tabla 6 se presentan los parámetros genéticos de forma independiente para los 3 periodos analizados. Solo en los dos periodos extremos (hasta 1990 con después de 2000) se puede hablar de rasgos independientes dado el valor de la correlación genética. Las correlaciones genéticas entre periodos adyacentes fueron superiores.
Continuando con los 3 diferentes periodos de tiempo para los 72 sementales que se encuentran representados no hubo cambios en el orden de mérito importantes y se muestra un paralelismo estable (Figura 2). Las correlaciones entre los VG variaron entre 0,93 comparando los periodos antes de 1990 y después de 2000 y 0,99 entre antes de 1990 y hasta el 2000, y las correlaciones de rango tuvieron la misma tendencia y se movieron entre 0,94 y 0,98. Las correlaciones entre los periodos más próximos tendieron a ser más altas. La amplitud entre los VG fue menor a medida que nos acercamos en el tiempo. Los VG antes de 1990 variaron entre -92,22 a +58,21 (Figura 3) y después de 2001 entre -21,74 a 20,84 (figura 4). Ambas distribuciones son parecidas y el mayor porcentaje de observaciones se encuentran desplazados hacia los valores negativos.
Tabla 6 Heredabilidades y correlaciones genéticas para el peso final en los 3 periodos analizados
| Hasta 1990 | 1991-2000 | Después de 2000 | |
|---|---|---|---|
| Hasta 1990 | 0,37 ± 0,20 | 0,93 ± 0,15 | 0,62 ± 0,15 |
| 1991-2000 | 0,27 ± 0,16 | 0,86 ± 0,16 | |
| Después de 2000 | 0,29 ± 0,18 |
Nota: heredabilidades en la diagonal y las correlaciones genéticas fuera de la diagonal.
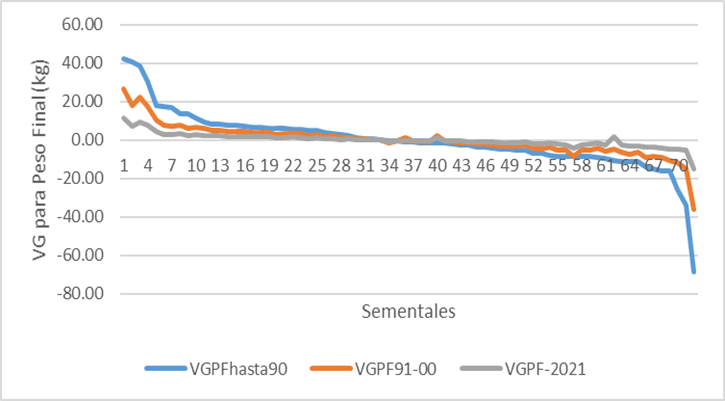
La tendencia genética con estos sementales se presenta en la figura 5 la que resulta muy errática (muy pocas observaciones) y tiende a cero, aunque es positiva (128 g/año). La tendencia fenotípica fue presentada para el mismo grupo de animales por Suárez Tronco et al (2021), demostrándose que el peso final en prueba de comportamiento se ha reducido en el tiempo y vemos en la figura 5 cómo la mejora genética no ha sido completamente capaz de contrarrestar los efectos ambientales (sistema de manejo y alimentación). En aquel trabajo se evidenció la influencia del índice temperatura humedad (ITH) que deprime el peso final en prueba de comportamiento como la variable climática principal. Se puede observar a partir de 1980, que después de un decrecimiento la tendencia vuelve a ser positiva. De todas formas, el R2 de la ecuación de regresión no fue significativa (Figura 5).
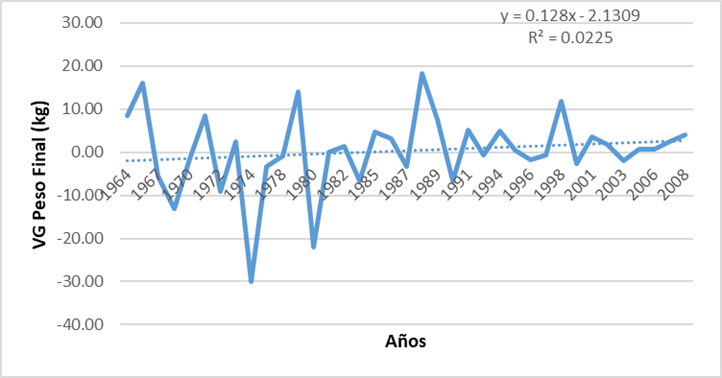
Figura 5 Tendencia genética del peso final en prueba de comportamiento para los 72 sementales considerados.
Ossa-Saraz et al (2014) en ganado Romosinuano encontraron tendencias genéticas para peso a 16 meses de edad similares (113 g/día) pero en un rebaño en conservación, mientras que Vergara-Garay, Simanca-Sotelo y Salgado-Otero (2014) en un rebaño multirracial de ganado de carne, también en Colombia, pero para la ganancia hasta 18 meses de edad encontraron tendencias negativas. Rocha-Rodrigues et al (2022) en ganado Nellore en Brasil encontraron tendencias genéticas para peso a 16 meses que oscilaron entre -1,29 y 12,51 kg/año, rango inferior al reportado en este trabajo, pero téngase en cuenta que se hizo con solo 72 sementales y por la vía macho.
Naves y Menéndez-Buxadera (2005) en ganado Criollo en Guadalupe analizaron el aumento de peso en 3 sistemas de pastoreo en pruebas de comportamiento y encontraron que el sistema de manejo y alimentación intensivo no representa el mismo carácter registrado en pastoreo o en un sistema intermedio y resaltan la importancia del ambiente pre destete que debe ser considerado en los modelos de análisis. Para este trabajo no tuvimos en consideración dicho ambiente, aunque para esta misma población de animales Criollos en Cuba (Suárez Tronco et al., 2022 a y b) estudiaron el peso al destete en relación con el índice temperatura humedad (ITH) y la carga calórica acumulada y en esta raza autóctona en Cuba desde hace más de 5 siglos, encontraron que el estrés calórico afecta el comportamiento pre destete, además se estimaron los componentes de (co)varianza genética y heredabilidad del estrés climático sobre el peso al destete (PD) y se pudo demostrar la existencia de una importante variabilidad genética en la forma de respuesta al estrés climático.
Carabaño et al (2019) en una revisión donde se aborda la selección para tolerancia al calor, y cómo medir dicha tolerancia en los animales y la variabilidad genética y su relación con las evaluaciones genéticas plantean que la selección de animales tolerantes y su antagonismo con la productividad puede ser un desafío, así como la relación existente con los sistemas de producción, y los complejos sistemas fisiológicos que intervienen en estos procesos.
Gredler-Grand et al., (2022) con ganado lechero irlandés puro y en cruces analizaron la edad de sacrificio a través de un gradiente ambiental y encontraron interacción genotipo ambiente en ambientes extremos.

