










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Ciencias Forestales
versión On-line ISSN 2310-3469
Rev CFORES vol.11 no.3 Pinar del Río sept.-dic. 2023 Epub 02-Sep-2023
Original article
Vascular epiphytes in a permanent plot in Andean forest in the "Francisco Vivar Castro" University Park, Loja, Ecuador
1Universidad nacional de Loja. Ecuador.
2Universidad de Pinar del Río "Hermanos Saíz Montes de Oca", Pinar del Río, Cuba
3Herbario "Reinaldo Espinosa", Loja Ecuador.
Most trees in Andean forests are covered by epiphytes. The objective of the study was to evaluate the floristic composition, structure and preference of vascular epiphytes in certain phorophytes. Three phorophytes were selected in each 20 x 20 m subplot of a one hectare permanent plot and recorded: D 1.30 > 20 cm, crown foliage, visibility and branching. The inventory of the epiphytes was carried out by climbing the phorophytes following their vertical distribution. Structural parameters such as density, abundance, frequency, importance value index and Shannon diversity were calculated. The habitat preference of epiphytes was calculated by the number of individuals per host and per section. 7,610 individuals of 12 species, 8 genera and 4 families were recorded. The most ecologically important species in all sections of the phorophytes are: Anthurium oxybelium, Peperomia galioides, Tillandsia tovarensis, Cyrtochilum aureum, Pleurothallis maxima, Tillandsia biflora. The vascular epiphytes recorded prefer to live in section 3, Tillandsia tovarensis, Pleurothallis maxima and Tillandsia biflora are the most abundant. Alnus acuminata and Cedrela montana are the most preferred phorophytes for vascular epiphytes.
Key words: Andean forest; diversity of vascular epiphytes; phorophyte; vertical distribution and habitat preference.
INTRODUCTION
Ecuador is one of the 17 megadiverse countries on the planet, due to its location in the center of the world, the presence of the Andes Mountains, the Coastal Region, the Amazon jungles and the Galapagos Islands (Aguirre 2018), it has great wealth floristics given by 18,568 species of vascular plants of which 25 % are endemic. A large part of this diversity and endemism of plants develop in the Andean forest, which is why it represents one of the most threatened biodiversity hot spots in the Northern Andes (Tejedor-Garavito et al., 2012).
In the case of Hoya de Loja, the Andean Forest is in danger of disappearing, about 3,459 hectares have been deforested during the period 2014 2016, due to conversion of use for agricultural expansion, forest fires and expansion of the urban area (MAE 2018). The disappearance of these not only causes the decline of forest species, but also affects one of the important ecological groups such as vascular epiphytes (MAE 2016 and Paredes-Ulloa et al., 2021).
Studying vascular epiphytes is important not only because they are a component of the diversity in tropical and Andean forests, but also because of the function they play in their recovery dynamics, since by stratifying vertically, they constitute habitat for fauna and offer a wide variety of niches and resources (Einzmann and Zotz 2016) for other groups. Therefore, the Andean Forest of the "Francisco Vivar Castro" University Park at an altitude of 2,250 m a.s.l., with an area of 12.93 ha (Aguirre et al., 2016) constitutes a site of interest for the study of this ecological group.
Under these considerations, the study aims to evaluate the composition, structure and habitat preference of vascular epiphytes in a permanent plot of one hectare of the Andean Forest.
MATERIALS AND METHODS
Study area
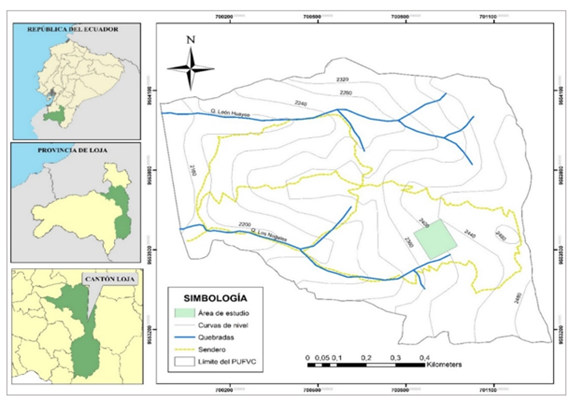
The study was carried out in the "Francisco Vivar Castro" University Park (PUFVC), property of the National University of Loja, in the province of Loja, located at the UTM coordinates: 700 5929 554 223 N, 700 9709 553 139 S, 701 3099 553 171 E, 699 9619 554 049 W, between 2 130 to 2 520 m asl and occupies an area of 99.13 ha (Figure 1). This area is part of the Project: Ecological processes of vegetation in the university park "Francisco Vivar Castro", National University of Loja. Phase III. 2022-2023.
Selection of forums
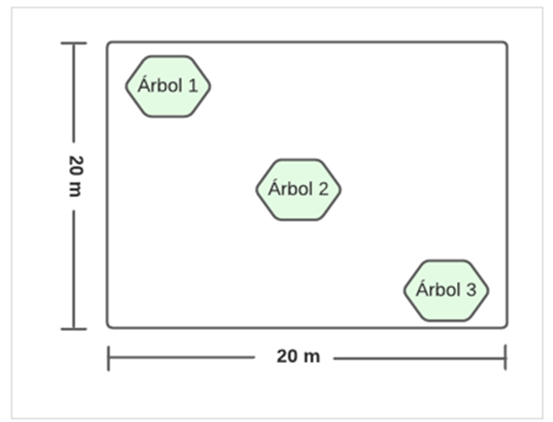
For this research, information from the tree census of a permanent plot of one hectare in the Andean Forest of the PUFVC was taken as a basis. In each of the 25 subplots of 20 x 20 m (400 m2), three trees (phorophytes) were selected, considering four aspects: D 1.30 m greater than 20 cm, crown width and visibility (height of crown and crown diameter) and branching (Figure 2).
Evaluation of vascular epiphyte assemblages in each phorophyte
The evaluation of epiphyte assemblages present in each phorophyte was carried out by direct observation with binoculars and climbing each tree under study. The individuals of vascular epiphytes (bromeliads, orchids, aroids and piperaces) present in each phorophyte were recorded. These were collected for later identification in the "Reinaldo Espinosa" Herbarium of the National University of Loja.
Data logging for structural parameters
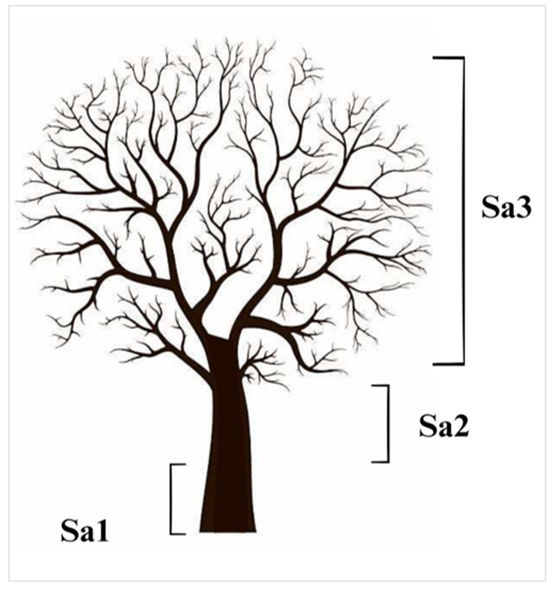
Given that the distribution of epiphytes on the phorophyte varies vertically and horizontally, each tree selected in the subplots was subdivided into three sections (Figure 3): section 1 (Sa1) from the base to 3 m; section 2 (Sa2) is the middle part, above section 1 and below the first branch; and section 3 (Sa3) from the first branch to the crown of the tree, this is a modification to the method of Johansson (1974).
Data analysis for composition and structure
With the data collected from the vascular epiphytes in each of the trees in the subplots, the structural parameters were calculated: relative density (RD), relative frequency (FR) and IVI. The species accumulation curve was made using the chao 2 index to check the representativeness of the sampling. To compare the density between vertical section of the phorophytes, the richness by botanical families and the density by species of epiphytes, a non-parametric Kruskal-Wallis test was carried out with a confidence level of 5 %. Statistical analyzes were used with the EstimateSWin910 statistical package.
The vascular epiphytes that have the greatest preference towards a host(s) were defined and the percentage of preference was calculated through a direct rule of three, because the magnitudes are directly proportional. In addition, the phorophytes with the greatest presence of epiphytic individuals in each section, the species with the greatest abundance of individuals and the sections that prefer these species were identified.
A cluster analysis was carried out to recognize the existence of groups among phorophytes according to the species of epiphytes that inhabit them; and another between the three sections. For this, the complete linkage method was used and 50 % similarity and 80 % respectively were considered as the cut-off point or reference line for defining the clusters.
RESULTS
Floristic composition and structure of vascular epiphytes
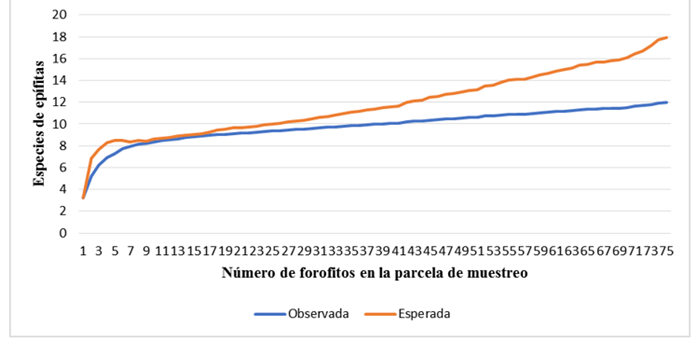
Figure 4 shows the species accumulation curve, which shows the sampling effort. This curve was adjusted with the Chao 2 estimator. The observed richness is 12 species, the non-parametric Chao 2 estimator estimates 18 species for the sample of 75 hosts. The sampling did not allow us to cover all the diversity of epiphytes, registering 67 % of the expected species.
12 species of epiphytes were recorded within eight genera and four families. The number of epiphytes recorded was 7,610 individuals in 75 phorophytes inventoried. Table 4 identifies the 21 host species of vascular epiphytes, with the sampling density and estimated density per hectare (Table 1).
Table 1. - Number of epiphyte individuals in the sampling and the estimated density for one hectare in the Andean Forest of the PUFVC
| host species | No.E/H | No em | No.H/ha | No.E/ha |
|
|
22 | 3 830 | 51 | 8 879 |
|
|
1 | 17 | 8 | 136 |
|
|
4 | 1 313 | eleven | 3 611 |
|
|
7 | 476 | 80 | 5 440 |
|
|
1 | 55 | 6 | 330 |
|
|
4 | 56 | 8 | 112 |
|
|
2 | 8 | 64 | 256 |
|
|
2 | 54 | fifteen | 405 |
|
|
1 | twenty | 39 | 780 |
|
|
1 | 4 | 1 | 4 |
|
|
2 | 65 | 23 | 748 |
|
|
4 | 497 | 73 | 9 070 |
|
|
1 | 70 | 7 | 490 |
|
|
2 | 173 | 30 | 2,595 |
|
|
5 | 128 | 44 | 1 126 |
|
|
2 | 263 | 19 | 2 499 |
|
|
2 | 62 | 14 | 434 |
|
|
4 | 213 | 36 | 1 917 |
|
|
1 | 42 | 66 | 2 772 |
|
|
6 | 211 | 22 | 774 |
|
|
1 | 53 | 2 | 106 |
Legend: No.E/H: Number of epiphytic individuals per host, No.E/m: Number of epiphytic individuals in the sample, No.H/ha: Estimated number of hosts per hectare; No.E/ha: Number of epiphytic individuals estimated for 1 ha.
Vertical structure of vascular epiphytes by sections
Table 2 shows the most abundant epiphytes by section; generally, in section 3 there was a greater number of epiphyte individuals and the unique presence of the orchid Cyrtidiorchis rhomboglossa. In section 1 the most abundant species were from the Piperaceae family: Peperomia alata and Peperomia galioides, followed by Anthurium oxybelium; while the least abundant ones corresponded to the Bromeliaceae and Orchidaceae families. In section 2 as in 3, the vascular epiphytes with the largest number of individuals corresponded to Tillandsia tovarensis (Bromeliacea), Cyrtochilum aureum and Pleurothallis maxima. The species with the fewest individuals were
Tillandsia cylindrica, Stelis emarginata and Epidendrum sp.
Tallandsia tovarensis is the only species that shows a drastic increase in the number of individuals, particularly between section 3 and the rest of the sections, presenting a marked presence in all the phorophytes of the plot. The species best represented in the number of individuals correspond to Pleurothallis maxima and Cyrtochilum aureum, which tend to increase towards the top of the trees (Table 2).
Table 2. - Structural parameters of the vascular epiphytes of section I in the permanent plot of the PUFVC
| Section 1 | |||||
| Family | Scientific name | d | Dr % | Fr % | IVI |
| Piperaceae |
|
292 | 54.07 | 36.78 | 45.43 |
| Araceae |
|
84 | 15.56 | 22.99 | 19.27 |
| Piperaceae |
|
44 | 8.15 | 14.94 | 11.55 |
| Orchidacea |
|
43 | 7.96 | 3.45 | 5.71 |
| Orchidacea |
|
3. 4 | 6.3 | 5.75 | 6.02 |
| Bromeliaceae |
|
28 | 5.19 | 9.2 | 7.19 |
| Bromeliaceae |
|
9 | 1.67 | 23 | 1.98 |
| Bromeliaceae |
|
3 | 0.56 | 1.15 | 0.85 |
| Araceae |
|
2 | 0.37 | 23 | 1.33 |
| Orchidacea |
|
1 | 0.19 | 1.15 | 0.67 |
| Total | 540 | ||||
| Section 2 | |||||
| Bromeliaceae |
|
350 | 41.03 | 36.27 | 38.65 |
| Orchidacea |
|
174 | 20.4 | 17.65 | 19.02 |
| Orchidacea |
|
158 | 18.52 | 10.78 | 14.65 |
| Piperaceae |
|
53 | 6.21 | 6.86 | 6.54 |
| Piperaceae |
|
51 | 5.98 | 7.84 | 6.91 |
| Bromeliaceae |
|
31 | 3.63 | 10.78 | 7.21 |
| Araceae |
|
24 | 2.81 | 5.88 | 4.35 |
| Bromeliaceae |
|
6 | 0.7 | 1.96 | 1.33 |
| Orchidacea |
|
5 | 0.59 | 0.98 | 0.78 |
| Orchidacea |
|
1 | 0.12 | 0.98 | 0.55 |
| Total | 853 | ||||
| Bromeliaceae |
|
4 846 | 78 | 39.47 | 58.73 |
| Orchidacea |
|
547 | 8.8 | 16.45 | 12.63 |
| Orchidacea |
|
333 | 5.36 | 11.84 | 8.6 |
| Bromeliaceae |
|
279 | 4.49 | 15.79 | 10.14 |
| Bromeliaceae |
|
187 | 3.01 | 13,16 | 8.08 |
| Piperaceae |
|
12 | 0.19 | 0.66 | 0.43 |
| Piperaceae |
|
8 | 0.13 | 1.32 | 0.72 |
| Araceae |
|
3 | 0.05 | 0.66 | 0.35 |
| Orchidacea |
|
2 | 0.03 | 0.66 | 0.35 |
| Total | 6 217 |
Note: D: Density; Dr:Relative Density; Fr:Relative frequency; IVI:Importance value index.
Comparison of species by family in each section of the phorophytes
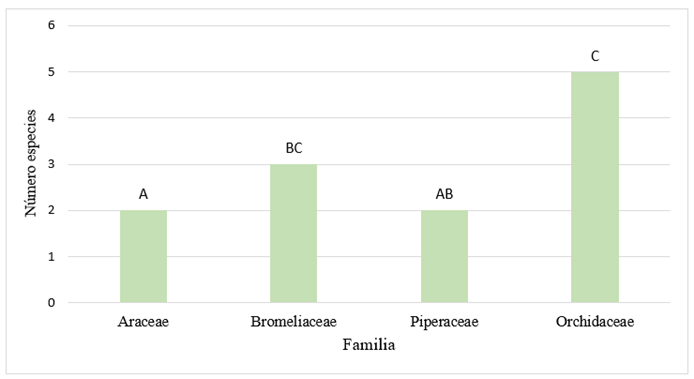
The Kruskal-Wallis test showed that there are no significant differences in the number of epiphyte species with respect to the phorophyte section. In the case of species richness per family, the test did show significant differences (Figure 5, P = 0.0189). The different letters indicate that there is a difference between the numbers of species per family.
Preference of vascular epiphytes towards hosts
It was detected that only four tree species are more preferred as hosts (Table 3), among these Alnus acuminata represents one of the most preferred hosts for epiphytes of any family, particularly for species of the Orchidaceae family.
Table 3. - Percentage of preference of the epiphytic species to the host in the permanent plot of the PUFVC
| Epiphytes | Host | % Pr | No.E |
|
|
22.52 | 25 | |
|
|
|
fifty | 1 |
|
|
|
fifty | 1 |
|
|
|
38.82 | 210 |
|
|
|
100 | 2 |
|
|
|
43.87 | 154 |
|
|
|
26.61 | 29 |
|
|
|
73.40 | 549 |
|
|
|
100 | 5 |
|
|
|
43.57 | 139 |
|
|
|
21.32 | 68 |
|
|
|
50.80 | 2 654 |
Note: % Pr: percentage of preference, No.E: Number of epiphyte individuals
Similarity between sections and between hosts by epiphytic load
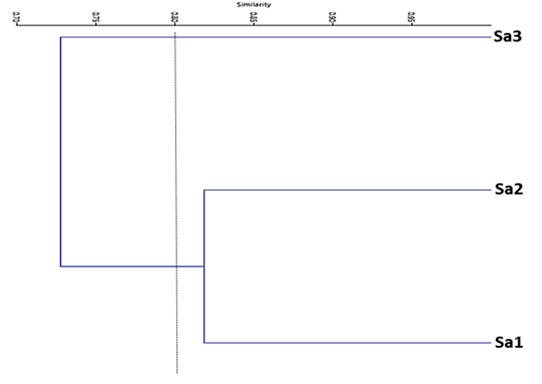
According to the position of the cut line in the dendrogram of the cluster analysis, two groups are distinguished (Figure 6), one that groups section 1 (Sa1) and section 2 (Sa2) and another for section 3 (Sa3).
The cluster analysis of the epiphytic species in relation to the host (Figure 7 and Table 4) shows that with 50 % similarity, 8 groups are identified. With the same association of epiphytes, the phorophytes Oreopanax rosei, Axinaea macrophylla and Morella interrupta and the group formed by Vismia baccifera, Alnus acuminata and Nectandra laurel are recognized. The phorophyte Siparuna muricata that differs the most in terms of epiphytic species in relation to the rest.
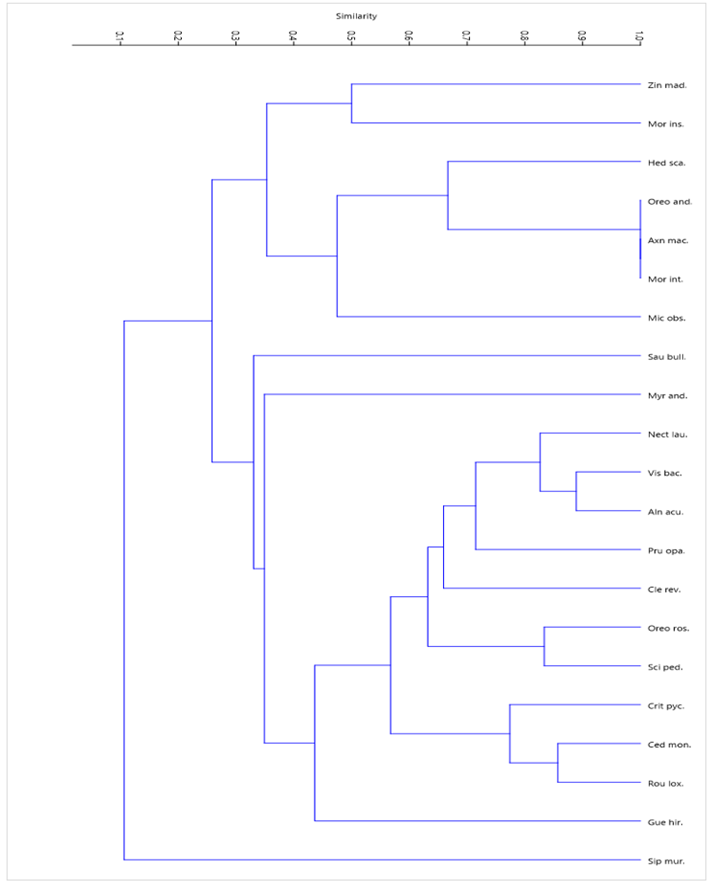 Legend: Sip mur.: Siparuna muricata (Ruiz y Pav.) A. DC, Gue hir.: Guettarda hirsuta (Ruiz y Pav.) Pers., Rou lox.: Roupala loxensis IM Johnst., Ced mon.: Cedrela montana Moritz ex Turcz, Cri pyc.: Critoniopsis pycnantha (Benth.) H. Rob., Sci ped.: Sciodaphyllum pedersenii, Pink Oreo: Oreopanax rosei Harms, Cle rev.: Clethra revoluta (Ruiz and Pav.) Spreng, Pru opa.: Prunus opaca (Benth.) Walp., Aln acu.: Alnus acuminata Kunth, Vis baca.: Vismia baccifera (L.) Triana and Planch., Nec lau.: Nectandra laurel Klotzsch ex Nees, Myr and.: Myrsine andina (Mez) Pipoly, Sau bull.: Saurauia bullosa Wawra, Mic obs.: Miconia obscura (Bonpl.) Naudin, Mor int.: Morella interrupta (Benth.) Lægaard, Axn mac.: Axinaea macrophylla (Naudin) Triana, Ore and.: Oreopanax andreanus Marchal, Hed scan.: Hedyosmum scabrum (Ruiz y Pav.) Solms, Mor ins.: Morus insignis Bureau, Zin mad.: Zinowiewia madsenii C. Ulloa and P. Jørg.
Legend: Sip mur.: Siparuna muricata (Ruiz y Pav.) A. DC, Gue hir.: Guettarda hirsuta (Ruiz y Pav.) Pers., Rou lox.: Roupala loxensis IM Johnst., Ced mon.: Cedrela montana Moritz ex Turcz, Cri pyc.: Critoniopsis pycnantha (Benth.) H. Rob., Sci ped.: Sciodaphyllum pedersenii, Pink Oreo: Oreopanax rosei Harms, Cle rev.: Clethra revoluta (Ruiz and Pav.) Spreng, Pru opa.: Prunus opaca (Benth.) Walp., Aln acu.: Alnus acuminata Kunth, Vis baca.: Vismia baccifera (L.) Triana and Planch., Nec lau.: Nectandra laurel Klotzsch ex Nees, Myr and.: Myrsine andina (Mez) Pipoly, Sau bull.: Saurauia bullosa Wawra, Mic obs.: Miconia obscura (Bonpl.) Naudin, Mor int.: Morella interrupta (Benth.) Lægaard, Axn mac.: Axinaea macrophylla (Naudin) Triana, Ore and.: Oreopanax andreanus Marchal, Hed scan.: Hedyosmum scabrum (Ruiz y Pav.) Solms, Mor ins.: Morus insignis Bureau, Zin mad.: Zinowiewia madsenii C. Ulloa and P. Jørg.Fig. 7. - Similarity between hosts due to the presence of epiphytes in the permanent plot
75 % of the epiphytes prefer Alnus acuminata as hosts, Cedrela montana 33 %, and Cletra revoluta 25 %, the rest of the tree species are used as hosts interchangeably (Table 4).
Tabla 4. - Porcentaje de preferencia de las especies epifitas hacia hospederos en la parcela permanente del bosque andino del PUFVC
| Epiphytic species | Host | % Preference |
|
|
22,52 | |
|
|
21,62 | |
|
|
11,71 | |
|
|
50 | |
|
|
50 | |
|
|
9,91 | |
|
|
38,82 | |
|
|
21,26 | |
|
|
13,68 | |
|
|
100 | |
|
|
100 | |
|
|
73,40 | |
|
|
10,16 | |
|
|
100 | |
|
|
43,87 | |
|
|
12,82 | |
|
|
11,97 | |
|
|
1,14 | |
|
|
11,97 | |
|
|
26,61 | |
|
|
10,09 | |
|
|
11,93 | |
|
|
22,94 | |
|
|
9,17 | |
|
|
43,57 | |
|
|
12,23 | |
|
|
16,30 | |
|
|
10,03 | |
|
|
3,14 | |
| Morella interrupta (Benth.) Lægaard | 0,31 | |
|
|
16,30 | |
|
|
21,32 | |
|
|
22,57 | |
|
|
50,80 | |
|
|
19,47 | |
|
|
0,06 | |
|
|
0,17 | |
| Oreopanax andreanus Marchal | 16,30 | |
|
|
1,01 | |
|
|
0,92 |
DISCUSSION
Floristic composition and structure of vascular epiphytes
In this study, individuals of epiphyte species were found that show preferences for certain phorophytes and sections within the phorophytes, with greater abundance in the tree crowns. The presence of 12 species from 8 genera and 4 families was determined, recorded in 75 sampled trees. Studies on epiphytes in Ecuadorian Andean forests are not so abundant, particularly due to the difficulty of field work (Paredes-Ulloa et al., 2021). In this regard, Henao-Díaz et al. (2012) reported a similar diversity and density of vascular epiphytes in Colombian sub-Andean and lowland forests and argued that this was in direct relation to host height and altitude above sea level. However, the present study shows a lower species richness, which could be due to the degree of forest intervention and its recovering condition (Aguirre et al., 2016). These alterations to natural forests, which produce changes in the elements of their structure, affect all the communities dependent on these ecosystems, mainly the epiphytes (Bartels 2012).
The families with the greatest number of species and individuals are Bromeliaceae and Orchidaceae, this is similar to what was reported by Paredes-Ulloa et al. (2021), which seems to be a regularity of the Andean forests of Ecuador. This representativeness is influenced by the morphological, ecological and physiological adaptations that these epiphytic species present in order to respond to various environmental restrictions such as light exposure, low nutrient availability, temperature changes and especially precipitation (Ding et al., 2016).
The high abundance of Tillandsia tovarensis in the forest can be explained by the tendency of the species to form conglomerates which sometimes almost completely cover the crowns of the phorophytes, not allowing the growth of other species around them. The Tillandsia genus, due to the characteristic of forming tanks or reservoirs with its leaves, is the key to reproduction and development through the retention of water and nutrients (Ding et al., 2016).
Regarding the richness by sections in the sampled phorophytes, there is great similarity in the number of species in the three sections of the trees, which determines that the variation in richness per vertical section is not significant. This result is different from Paredes-Ulloa et al. (2021) in a piedmont evergreen forest where epiphytes (orchids) are clearly concentrated in the upper canopy. However, in relation to the density of epiphytes there were significant differences between sections (Figure 6), so the existence of a selectivity of epiphytic individuals in the highest sites is recognized. This behavior of greater concentration of individuals in the upper canopy was consistent with the studies of Krömer et al. (2007) and Paredes-Ulloa et al. (2021), which should be related to the best availability of light for vegetative and photosynthetic activities.
So, in the vertical distribution, the preference of the species to a particular section is observed. An example of this is the bromeliads, which are present in all sections but in greater frequency and density in the upper one. Furthermore, in high sections the wind speed and solar radiation increase while the relative humidity decreases, which provides the formation of microclimates in the forest sections, a situation that affects the presence of some groups of vascular epiphytes.
In the case of orchids, there is no generality for some species, the density increases as one ascends through the sections of the host, such as in Cyrtochilum aureum and Pleurothallis maxima. However, this did not occur with Cyrtidiorchis rhomboglossa, Epidendrum sp. and Stelis emarginata, all these orchids. In this regard, Rasmussen and Rasmussen (2018) and Paredes-Ulloa et al. (2021) report that this family tends to have greater species richness but with low density of individuals. Another possible cause of the limited number of individuals could be related to external morphology since these species have poor anchorage , Rasmussen and Rasmussen (2018). The higher frequency and density of Peperomia alata and Peperomia galioides, in section 1 of the phorophytes could be related to the higher humidity (Mai et al., 2016 and Martínez-Bautista et al., 2019).
The species diversity in the different sections was medium to low in terms of composition and abundance, respectively compared to the results of Rasmussen and Rasmussen (2018), and may be the response to forest alterations in the last 30 years since the Wealth is positively associated with the age of succession (Paredes-Ulloa et al., 2021).
Habitat preference by vascular epiphytes
Characteristics such as the presence of emerging adult trees with abundant and thick branches, rough bark determine the presence of epiphytes, particularly their richness (Armijos et al., 2017). In this regard, Krömer et al. (2007) and Rasmussen and Rasmussen (2018) report that elements that facilitate the colonization of epiphytes are determinants in habitat preference such as the size of the tree, since it regulates the intensity of light captured by epiphytes in the canopy or interior, the structure of the tree, presence of the number of thick branches, a rough bark, the presence of metabolites that favor or inhibit its development and not to mention the retention of water and nutrients. However, Paredes-Ulloa et al. (2021) suggest that the most important characteristic is the diameter of the host, which facilitates the presence of a greater number of epiphytes.
The abundance of Alnus acuminata as a preferred phorophyte for nine species of vascular epiphytes could be related not only to the abundance of tree species in the area but also to the physiognomy of the specimens, since the vast majority had a large, highly branched appearance, with semi-rough bark and spaces between the branches like a slit that favors the preference of the phorophyte (Martínez-Meléndez et al., 2008 and Salazar-Ramirez et al., 2014). But Cedrela montana, with lower abundance in the area, had a high value as a preferred species, which is related to the presence of irregular longitudinal fissures in its trunk that determines the preference for its bark and tree structure.
CONCLUSIONS
In the Andean forest, 7,610 individuals of vascular epiphytes were recorded in four families and 12 species, where Tillandsia tovarensis, Pleurothallis maxima and Cyrtochilum aureum were the most abundant.
There is a distribution pattern in species and abundance in the studied families, Bromeliaceae and Orchidaceae towards section 3, while Piperaceae and Araceae were concentrated towards sections 1 and 2 of the phorophytes due to the higher humidity.
Two species of phorophytes are preferred: Alnus acuminata and Cedrela montana for their diameter, branching and bark characteristics.
In the Andean forest of the "Francisco Vivar Castro" University Park, vascular epiphytes have low and medium diversity in the three sections, reflecting its recovery and growth process.
REFERENCIAS BIBLIOGRÁFICAS
AGUIRRE, Z., 2016. Escenarios para la enseñanza y valoración de la biodiversidad en la región sur del Ecuador. Bosques Latitud Cero [en línea], vol. 6, no. 2, Disponible en: https://www.researchgate.net/publication/313854024_ESCENARIOS_PARA_LA_ENSENANZA_Y_VALORACION_DE_LA_BIODIVERSIDAD_EN_LA_REGION_SUR_DEL_ECUADOR. [ Links ]
AGUIRRE, Z., 2018. Biodiversidad ecuatoriana...estrategias, herramientas e instrumentos para su manejo y conservación [en línea]. Ecuador: Universidad Nacional de Loja (Cámara Nacional del Libro). ISBN 978-9942-35-685-7. Disponible en: https://www.researchgate.net/publication/329216867_BIODIVERSIDAD_ECUATORIANAESTRATEGIAS_HERRAMIENTAS_E_INSTRUMENTOS_PARA_SU_MANEJO_Y_CONSERVACION. [ Links ]
ARMIJOS MONTAÑO, A., ALVARADO CHAMBA, Y., QUITO TORRES, J., LEÓN GONZÁLEZ, T., GUAMÁN GUAMÁN, L., y PUCHA COFREP, D., 2017. Anatomía de la madera de diez especies forestales de bosque andino del sur del Ecuador. Cedamaz [en línea], vol. 7, Disponible en: https://www.researchgate.net/publication/323794556_Anatomia_de_la_madera_de_diez_especies_forestales_de_bosque_andino_del_sur_del_Ecuador_Wood_anatomy_of_ten_tree_species_from_Andean_forest_in_southern_Ecuador [ Links ]
BARTELS, S.F., y CHEN, H.Y.H., 2012. Mechanisms Regulating Epiphytic Plant Diversity. Critical Reviews in Plant Sciences [en línea], vol. 31, no. 5, [consulta: 04/10/2023]. ISSN 0735-2689. DOI 10.1080/07352689.2012.680349. Disponible en: Disponible en: https://doi.org/10.1080/07352689.2012.680349 . [ Links ]
DING, Y., LIU, G., ZANG, R., ZHANG, J., LU, X., y HUANG, J., 2016. Distribution of vascular epiphytes along a tropical elevational gradient: Disentangling abiotic and biotic determinants. Scientific Reports [en línea], vol. 6, no. 1, DOI 10.1038/srep19706. Disponible en: https://www.researchgate.net/publication/287992683_Distribution_of_vascular_epiphytes_along_a_tropical_elevational_gradient_Disentangling_abiotic_and_biotic_determinants [ Links ]
EINZMANN, H.J.R., y ZOTZ, G., 2016. How Diverse are Epiphyte Assemblages in Plantations and Secondary Forests in Tropical Lowlands? Tropical Conservation Science [en línea], vol. 9, no. 2, [consulta: 04/10/2023]. ISSN 1940-0829. DOI 10.1177/194008291600900205. Disponible en: Disponible en: https://doi.org/10.1177/194008291600900205 . [ Links ]
HENAO-DÍAZ, L.F., PACHECO-FERNÁNDEZ, N.M., ARGÜELLO-BERNAL, S., MORENO-AROCHA, M.M., y STEVENSON, P.R., 2018. Patrones de diversidad de epífitas en bosques de tierras bajas y subandinos. Colombia forestal [en línea], vol. 15, no. 2, Disponible en: https://www.redalyc.org/pdf/4239/423939618002.pdf [ Links ]
JOHANSSON, D., 1974. Ecology of vascular epiphytes in West African rain forest [en línea]. Uppsala, Sweden: Svenska vaxtgeografiska sallskape. ISBN 91-7210-059-1. Disponible en: https://www.diva-portal.org/smash/get/diva2:565496/FULLTEXT01.pdf. [ Links ]
KRÖMER, T., GRADSTEIN, S.R., y ACEBEY, A., 2007. Diversidad y ecología de epífitas vasculares en bosques montanos primarios y secundarios de Bolivia. Ecología en Bolivia [en línea], vol. 42, no. 1, [consulta: 04/10/2023]. ISSN 1605-2528. Disponible en: Disponible en: http://www.scielo.org.bo/scielo.php?script=sci_abstract&pid=S1605-25282007000400003&lng=es&nrm=iso&tlng=es . [ Links ]
MAI MORENTE, P., ROSSADO, A., BONIFACINO, J., y WAECHTER, J., 2016. Taxonomic revision of Peperomia (Piperaceae) from Uruguay. Phytotaxa [en línea], vol. 244, no. 2, DOI 10.11646/phytotaxa.244.2.2. Disponible en: https://www.researchgate.net/publication/290963498_Taxonomic_revision_of_Peperomia_Piperaceae_from_Uruguay. [ Links ]
MARTÍNEZ BAUTISTA, B., BERNAL RAMÍREZ, L., BRAVO AVILEZ, D., SAMAIN, M.S., AMEZCUA, J., y RENDON, B., 2019. Traditional Uses of the Family Piperaceae in Oaxaca, Mexico. Tropical Conservation Science [en línea], vol. 12, no. 2, DOI 10.1177/1940082919879315. Disponible en: https://www.researchgate.net/publication/337437346_Traditional_Uses_of_the_Family_Piperaceae_in_Oaxaca_Mexico. [ Links ]
MARTÍNEZ MELÉNDEZ, N., PÉREZ FARRERA, M.A., y FLORES PALACIOS, A., 2008. Estratificación vertical y preferencia de hospedero de las epífitas vasculares de un bosque nublado de Chiapas, México. Revista de Biología Tropical [en línea], vol. 56, no. 4, [consulta: 04/10/2023]. ISSN 0034-7744. Disponible en: Disponible en: http://www.scielo.sa.cr/scielo.php?script=sci_abstract&pid=S0034-7442008000400037&lng=en&nrm=iso&tlng=es . [ Links ]
MINISTERIO DEL AMBIENTE DEL ECUADOR, 2018. Estadísticas del patrimonio natural del Ecuador continental [en línea]. Ecuador: Ministerio del Ambiente. Disponible en: https://proamazonia.org/wp-content/uploads/2019/10/ECUADOR_Folleto_Patrimonio_Natural_compressed.pdf. [ Links ]
PAREDES ULLOA, C.O., FERRO DÍAZ, J., y LOZANO CARPIO, P., 2021. Efecto del estadio sucesional del bosque sobre la relación hospederos-orquídeas epífitas en la estación biológica Pindo Mirador, Ecuador. Revista Cubana de Ciencias Forestales [en línea], vol. 9, no. 1, [consulta: 04/10/2023]. ISSN 2310-3469. Disponible en: Disponible en: https://cfores.upr.edu.cu/index.php/cfores/article/view/678 . [ Links ]
RASMUSSEN, H., y RASMUSSEN, F., 2018. The epiphytic habitat on a living host: Reflections on the orchid-tree relationship. Botanical Journal of the Linnean Society [en línea], vol. 186, no. 4, DOI 10.1093/botlinnean/box085. Disponible en: https://www.researchgate.net/publication/324494440_The_epiphytic_habitat_on_a_living_host_Reflections_on_the_orchid-tree_relationship. [ Links ]
SALAZAR RAMÍREZ, L.F., PINEDA GÓMEZ, D.M., ESTÉVEZ VARÓN, J.V., y CASTAÑO VILLA, G.J., 2014. Riqueza y abundancia de aves frugívoras y nectarívoras en una plantación de aliso (Alnus acuminata) y un bosque secundario en los andes centrales de Colombia. Boletín Científico Centro de Museos Museo de Historia Natural [en línea], vol. 18, no. 1, [consulta: 04/10/2023]. ISSN 2462-8190. Disponible en: Disponible en: https://revistasojs.ucaldas.edu.co/index.php/boletincientifico/article/view/4461 [ Links ]
TEJEDOR GARAVITO, N., ÁLVAREZ, E., ARANGO CARO, S., ARAUJO MURAKAMI, A., BLUNDO, C., y BOZA ESPINOZA, T.E., 2012. Evaluación del estado de conservación de los bosques montanos en los Andes tropicales. Ecosistemas [en línea], vol. 21, no. 1-2, [consulta: 04 /10/2023]. ISSN 1132-6344, 1697-2473. Disponible en: Disponible en: https://www.redalyc.org/articulo.oa?id=54026849012 . [ Links ]
Received: July 25, 2023; Accepted: August 16, 2023

