










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión impresa ISSN 0258-5936
cultrop v.29 n.4 La Habana oct.-dic. 2008
Influencia de la concentración de melaza y extracto acuoso de soya sobre la velocidad específica de crecimiento de Bradyrhizobium elkanii ICA 8001
Ms.C. Gretel GómezI, Dra. María C. NápolesII, Dr.C. R. NúñezIII y Dr.C. J. MartínezIV
I Investigadora, email: gretole@gmail.com
II Investigadora Titular del departamento de Fisiología y Bioquímica Vegetal, Instituto Nacional de Ciencias Agrícolas (INCA), gaveta postal 1, San José de las Lajas, La Habana, CP. 32 7001
III Investigador Titular del Departamento de Microbiología Aplicada, Centro de Bioproductos Marinos, La Habana
IV Profesor Titular del Centro de Estudios de Proteínas, facultad de Biología, Universidad de La Habana (UH), Cuba.
ABSTRACT
The strain ICA 8001 from Bradyrhizobium elkanii is broadly used in Cuba for soybean inoculant production. One culture medium designed to multiply this microorganism contains molasses and aqueous soybean extract, which are elements of a variable composition and origin providing considerable amounts of carbonated substrates. Therefore, concentrations of total soluble carbohydrates and reducing carbohydrates as well as N, P, K, Ca, Mg and Na contents were determined, to get a partial nutritional characterization of these components. Analyses showed substantial quantities of these bacterium-absorbing elements within cell growth. The influence of molasses and aqueous soybean extract concentrations on the specific strain growth rate m (h-1) was estimated by varying molasses concentrations in Bradyfact medium from 3 to 20 g.L-1 and aqueous soybean extract from 5 to 20 g.L-1. Growth dynamics were recorded for each variant, besides calculating m. Results proved that higher than 10 and 12,5 g.L-1 molasses and aqueous soybean extract concentrations, respectively, inhibit Bradyrhizobium elkanii ICA 8001 growth. This phenomenon could be mathematically and physiologically explained through two new kinetic models. This research work contributes to Bradyrhizobium physiological knowledge, since it makes evident the relationship between culture medium composition and microorganism nutritional requirements, also how culture medium design may ensure more effective inoculants containing high cell concentrations.
Key words: Bradyrhizobium elkanii, molasses, seed extracts, soybean, inhibition, growth, models
RESUMEN
La cepa ICA 8001 de Bradyrhizobium elkanii es ampliamente usada en Cuba para la producción de inoculantes para soya. Uno de los medios de cultivos diseñados para la multiplicación de este microorganismo contiene melaza y extracto acuoso de soya, elementos de variable composición y origen, que aportan cantidades notables de sustratos carbonados. Por esta causa, se determinó la concentración de carbohidratos totales solubles y carbohidratos reductores, así como los contenidos de N, P, K, Ca, Mg y Na, para obtener una caracterización nutricional parcial de estos componentes. Los análisis mostraron la presencia de cantidades apreciables de estos elementos asimilables por la bacteria durante su crecimiento celular. La influencia de las concentraciones de melaza y extracto acuoso de soya sobre la velocidad específica de crecimiento m (h-1) de la cepa se estimó, variando las concentraciones de melaza en el medio Bradyfact de 3 a 20 g.L-1 y del extracto acuoso de soya de 5 a 20 g.L-1. Se obtuvieron las dinámicas de crecimiento para cada variante y se calculó la m. Los resultados evidenciaron que concentraciones de melaza y extracto acuoso de soya superiores a 10,0 y 12,5 g.L-1, respectivamente, inhiben el crecimiento de Bradyrhizobium elkanii ICA 8001. Este fenómeno pudo ser explicado matemática y fisiológicamente mediante dos nuevos modelos cinéticos. Este trabajo contribuye al conocimiento de la fisiología de Bradyrhizobium, pues evidencia la relación entre la composición del medio de cultivo y los requerimientos nutricionales del microorganismo, así como el diseño del medio de cultivo puede garantizar la obtención de inoculantes más efectivos que contengan elevadas concentraciones celulares.
Palabras clave: Bradyrhizobium elkanii, melaza, extractos de semilla, soja, inhibición, crecimiento, modelos
Introducción
En todo el mundo y desde hace mucho tiempo, se han utilizado los inoculantes comerciales a partir de cepas de Bradyrhizobium para inocular las semillas de soya (Glycine max).
En Cuba se han obtenido muy buenos resultados, lográndose suplir las necesidades de nitrógeno de este cultivo entre un 80 y 100 % (1). La cepa ICA 8001 de Bradyrhizobium elkanii es efectiva y eficiente para numerosas variedades comerciales cubanas o introducidas, por lo que es ampliamente usada en nuestro país en la producción de inoculantes.
La composición del medio de cultivo puede influir en diferentes aspectos de la fisiología del microorganismo: su nutrición, multiplicación, la producción de metabolitos primarios y secundarios (2). Por tanto, una etapa crucial para la obtención de biopreparados de alta calidad, es el desarrollo de un adecuado medio de cultivo.
Este paso incluye la selección del tipo y la concentración de los diferentes nutrientes (3).
Se ha demostrado el papel nutricional de la melaza y los extractos de semillas en la multiplicación celular de los microrganismos, por lo que estos sustratos se han convertido en valiosos componentes para el diseño de medios de cultivo (4, 5).
La composición química de la melaza (6) y del molinado de semillas de soya es muy compleja (7), y varía en función del origen de estos sustratos.
La mayoría de los sustratos son capaces de presentar una acción inhibitoria reversible en relación con su concentración sobre el crecimiento de las poblaciones microbianas, incluso la glucosa puede restringir el crecimiento de microrganismos al incrementar su concentración inicial en el medio de cultivo (8). Por todo lo anteriormente expuesto, los objetivos de este trabajo fueron caracterizar nutricionalmente los componentes melaza y molinado de soya, con el fin de conocer la cantidad de nutrientes que aportan al medio de cultivo Bradyfact; determinar la influencia de la concentración de melaza y extracto acuoso de soya sobre la velocidad específica de crecimiento de Bradyrhizobium elkanii ICA 8001, así como buscar modelos que expliquen matemática y fisiológicamente dicha influencia.
Materiales y Métodos
Microrganismo y material vegetal empleado. Se utilizó la especie Bradyrhizobium elkanii, cepa ICA 8001, proveniente de la colección del Instituto de Ciencia Animal (ICA), ubicado en Catalina, La Habana. Para la obtención del extracto acuoso se empleó la especie Glycine max (soya), variedad Incasoy 27.
Medios de cultivo. Se utilizaron los medios de cultivo Agar manitol extracto de levadura y Bradyfact (9). Dada la importancia de este último medio en la investigación, a continuación se detalla su composición (g.L-1): melaza 5,0; (NH4)2 SO4 0,5; K2HPO4 3 H2O 0,5; MgSO4.7H2O 0,2; NaCl 0,1; CaCO3 1,0 y extracto acuoso de soya (mL) 10.0. El extracto acuoso de soya se obtuvo a partir de la decantación y filtración de una disolución de 100 g de semillas molidas en un litro de agua destilada, la cual se agitó durante dos horas a 300 rpm en un agitador magnético y posteriormente se dejó en reposo durante el mismo tiempo.
La melaza utilizada provino del complejo agroindustrial “Héctor Molina”, provincia La Habana.
Condiciones de cultivo. Para la obtención de los pre-inóculos se partió de tubos de vidrio con medio Agar manitol extracto de levadura más indicador rojo congo en plano inclinado con Bradyrhizobium elkanii ICA 8001, cultivados durante una semana. Erlenmeyers de 250 mL con 50 mL de medio Bradyfact se inocularon con tres asadas de los tubos y se cultivaron durante 60 h. Pasado este tiempo, la biomasa microbiana fue inoculada al 8 % (v/v) en Erlenmeyers de 500 mL, con un volumen efectivo de medio de cultivo de 100 mL. Todas las fermentaciones se llevaron a cabo en zaranda orbital BIOZART 2013 a una velocidad de rotación de 100 rpm y una temperatura de 28 ± 2oC; los tiempos de cultivo variaron según el objetivo de cada experimento.
Determinación del crecimiento microbiano. El crecimiento se determinó por la concentración celular a través de la absorbancia de los cultivos, a una longitud de onda de 600 nm en un espectrofotómetro uv/visible Genesys 10 uv. Se utilizó como blanco el medio de cultivo Bradyfact sin inocular.
Caracterización nutricional de la melaza y del molinado de semillas de soya. Para la determinación de las concentraciones de carbohidratos totales solubles presentes en estos dos componentes, se empleó el método descrito (10) y la determinación de la concentración de carbohidratos reductores (CHR) se desarrolló a través de la metodología descrita (11). Los contenidos de N, P, K, Ca, Mg y Na se determinaron según el Manual de técnicas analíticas para la caracterización de abonos y suelos (12).
Influencia de la concentración de melaza sobre la velocidad específica de crecimiento de Bradyrhizobium elkanii ICA 8001. El estudio de la influencia de la concentración de melaza sobre la velocidad específica de crecimiento del microrganismo fue realizado con el medio Bradyfact, con concentraciones entre 3 y 20 g.L-1. Se tomaron alícuotas de 2 mL cada 8 h, lo que permitió obtener las dinámicas de crecimiento para cada variante mediante la determinación de la absorbancia a una longitud de onda de 600 nm.
La velocidad específica de crecimiento m(h-1) se calculó en la fase exponencial del crecimiento, utilizando el modelo de crecimiento no restringido (13). Para el cálculo se escogieron los mismos tiempos en todos los perfiles de fermentación.
Para calcular los parámetros cinéticos se empleó el modelo de Monod (13), que está descrito por la siguiente ecuación:
![]()
Influencia de la concentración de extracto acuoso de soya sobre la velocidad específica de crecimiento de Bradyrhizobium elkanii ICA 8001. Para evaluar el efecto de este componente del medio de cultivo sobre la velocidad específica de crecimiento de Bradyrhizobium elkanii ICA 8001, se siguieron las mismas condiciones y métodos que para la melaza, solo que las concentraciones de extracto acuoso de soya en el medio Bradyfact variaron entre 5 y 20 g·L-1.
Diseño de experimentos y análisis estadísticos. Se utilizaron diseños completamente aleatorizados para la concepción de todos los experimentos. Todos los datos se sometieron a la prueba de normalidad de Kolmogorov-Smirnov y homogeneidad de varianza de Bartlett.
Los experimentos se realizaron con tres repeticiones. Se hizo el análisis de varianza de clasificación simple y la prueba de comparación de medias de rangos múltiples de Duncan, en la cual las medias que no se diferencian se expresan con igual letra. Los datos fueron procesados con los paquetes estadísticos Statgraphics plus 5.0 y Excel 7.0 para Windows.
Resultados y Discusión
Caracterización nutricional de la melaza y el molinado de semillas de soya. Debido a la crucial importancia que juegan las melazas y los extractos de semillas en la nutrición microbiana (5, 14, 15, 16), estos compuestos son considerados macronutrientes y presentan un elevado potencial como materias primas de bajo costo para numerosos procesos fermentativos a nivel industrial.
En la Tabla I se muestran los resultados del análisis químico realizado a la melaza y al molinado de semillas de soya. Esta caracterización mostró cantidades apreciables de elementos asimilables por la bacteria durante su crecimiento celular. Ambos componentes contienen concentraciones considerables de carbohidratos solubles, nitrógeno, fósforo y un adecuado balance de los cationes: Na+, K+, Mg2+ Ca2+.

En el caso de la melaza, se apreció una concentración elevada de carbohidratos totales solubles de un 46 %, cifra muy cercana al valor promedio informado para las mieles cubanas de un 47 % (17), así como de carbohidratos reductores en comparación con el extracto de soya, el cual se destacó por presentar un alto contenido de nitrógeno.
Estos componentes aportan las siguientes concentraciones, según las proporciones en que se encuentran en el medio de cultivo Bradyfact (g.L-1): 2,54 de CHTS; 1,75 de CHR; 0,617 de nitrógeno; 0,067 de fósforo; 0,14 de Ca2+; 0,047 de Mg2+ y trazas de Na+ y K+.
Debido a la alta concentración de carbohidratos presentes en estos componentes del medio de cultivo (5, 17), se realizó un estudio de los parámetros cinéticos de la cepa frente a estos sustratos, con el objetivo de conocer si ocurría inhibición del crecimiento por exceso de sustrato a determinadas concentraciones de dichos elementos.
Influencia de la concentración de melaza sobre la velocidad específica de crecimiento de Bradyrhizobium elkanii cepa ICA 8001. A continuación se muestran los resultados del estudio cinético del crecimiento de la cepa ICA 8001 frente a diferentes concentraciones de melaza (Figura 1).
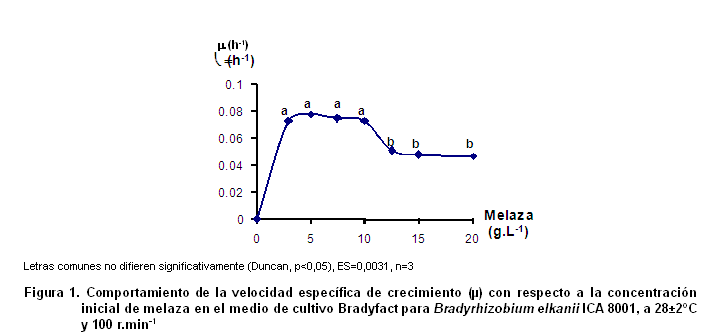
En este caso se pudieron apreciar dos comportamientos diferentes. Para el rango de concentraciones de melaza entre 3 y 10 g.L-1 se encontraron los máximos valores de velocidad específica de crecimiento (µ), correspondiéndose con una zona de saturación, debido a que dichas velocidades no difieren significativamente entre ellas. Cuando los valores de concentración superan los 10 g·L-1, los valores de µ disminuyen significativamente desde 0,073 hasta 0,047 h-1. Este resultado reveló que valores de concentración inicial de melaza superiores a 10 g.L-1 afectan la velocidad específica de crecimiento de Bradyrhizobium elkanii ICA 8001.
Muchos géneros bacterianos expresan un mecanismo de inhibición del crecimiento provocado por altas concentraciones de carbohidratos, debido a que los complejos enzimáticos responsables del transporte al interior de la célula pueden llegar a saturarse (18).
Por ejemplo, se observó la inhibición del crecimiento de una cepa de Rhizobium tropici cuando se utilizaron concentraciones iniciales de sacarosa en el medio mayores de 13 g.L-1 (19).
Para la producción de poli-b-hidroxibutirato (PHB) por Alcaligenes eutrophus se empleó la melaza de caña como suplemento nutricional. La adición de este componente a un medio sintético, a una concentración de 3 g.L-1, estimuló tanto la formación de biomasa como la producción de PHB. Sin embargo, cuando se suplementó con una concentración de 5 g.L-1, se evidenció un efecto inhibitorio sobre el crecimiento de este microrganismo (20).
Este fenómeno de inhibición también pudo ocurrir, debido a que las mieles de caña presentan elementos que resultan indeseables para el crecimiento celular, por su actividad inhibitoria, como suelen ser los ácidos fenólicos, ácidos orgánicos volátiles y ácidos grasos.
Entre los modelos matemáticos que describen el crecimiento microbiano, los más simples son los no estructurados y no segregados; el modelo de Monod es un ejemplo. Estos modelos matemáticos juegan un papel fundamental en las investigaciones actuales, porque contribuyen al ahorro de tiempo, ya que permiten calcular determinados parámetros cinéticos y, a su vez, la predicción de los resultados a obtener.
Los datos obtenidos para la cepa ICA 8001 no se ajustaron a los modelos de inhibición encontrados en la literatura. Para determinar los parámetros cinéticos, fue necesario proponer un modelo que fuera una modificación del modelo de Monod para la zona de inhibición y que permitiera explicar matemática y fisiológicamente el comportamiento de la µ de la cepa respecto a la concentración de melaza en nuestras condiciones experimentales.
El modelo de Monod clásico se ajustó a la zona donde no existe inhibición, hasta concentraciones de melaza de 10 g.L-1 y se pudo determinar la constante de afinidad por el sustrato (Ks=0,162 g.L-1) y la velocidad máxima de crecimiento (mmax=0,077h-1).
Para la zona de inhibición se realizó una modificación, como se presenta en la siguiente ecuación:
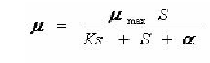
Donde a corresponde al término de inhibición y está en función de la concentración inicial de sustrato (S), la constante de inhibición del sistema (Ki) y el coeficiente de inhibición (n)
![]()
A partir de la ecuación y=1.476x-1.858 obtenida del gráfico lnS (y) vs lnS (x), se determinó la Ki=6,409 y el n=1,477 del sistema.
El modelo propuesto se presenta en la siguiente ecuación:
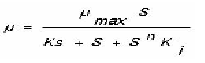
En la Figura 2 se muestran los resultados experimentales y los obtenidos a partir del modelo para la cepa estudiada.
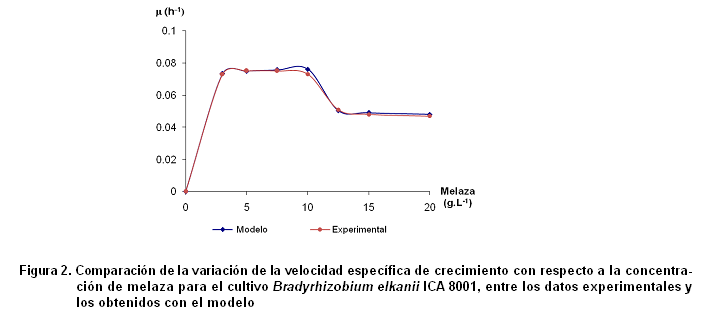
El modelo correlaciona con un valor r2= 0,993, los valores experimentales de µ y los obtenidos por el modelo propuesto.
Influencia de la concentración de extracto acuoso de soya sobre la velocidad específica de crecimiento de Bradyrhizobium elkanii ICA 8001. En la Figura 3 se muestra el comportamiento de la velocidad específica de crecimiento (µ) con respecto a la concentración inicial de extracto acuoso de soya en el medio de cultivo Bradyfact.
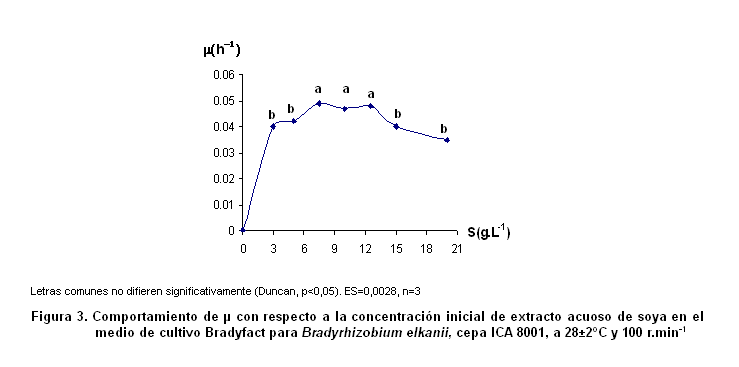
Al igual que en el caso de la melaza, se encontraron comportamientos diferentes. Entre 7,5 y 12,5 g.L-1 de extracto acuoso de soya, se observaron los más altos valores de µ y no se apreció una variación significativa entre ellos. Tanto para las menores concentraciones del extracto, 3 y 5 g.L-1, como para los valores por encima de 12,5 g.L-1, se obtuvieron µ significativamente inferiores.
En el caso del extracto acuoso de soya ocurrió el mismo fenómeno de inhibición que presentó la melaza, pues la velocidad específica de Bradyrhizobium elkanii, cepa ICA 8001, disminuye significativamente cuando se adicionaron al medio concentraciones iniciales mayores de 12,5 g.L-1 (Figura 3). Este resultado coincide con el informado para esta misma bacteria (9).
Este fenómeno de inhibición pudiera explicarse por diversas razones, como es el caso del exceso de sustrato carbonado derivado del empleo de dosis con elevadas concentraciones de este elemento. Aunque es más probable que la presencia de factores antinutricionales en la soya, tales como los inhibidores de tripsina, hemaglutininas y factores antivitamínicos (20), hayan podido influir negativamente en la fisiología del microorganismo.
La constante de afinidad por el sustrato (Ks) y la velocidad máxima de crecimiento (mmax) se calcularon mediante el empleo del modelo de Monod clásico, teniendo en cuenta la zona de concentraciones de extracto acuoso de soya hasta 12,5 g.L-1, donde no existe inhibición del crecimiento. Se obtuvieron valores de Ks = 0,943 g.L-1 y de mmax=0,052 h-1.
Para la zona de inhibición se realizó una modificación, como se presenta en la siguiente ecuación.

En la Figura 4 se muestran los resultados experimentales y los obtenidos a partir del modelo para la cepa estudiada.
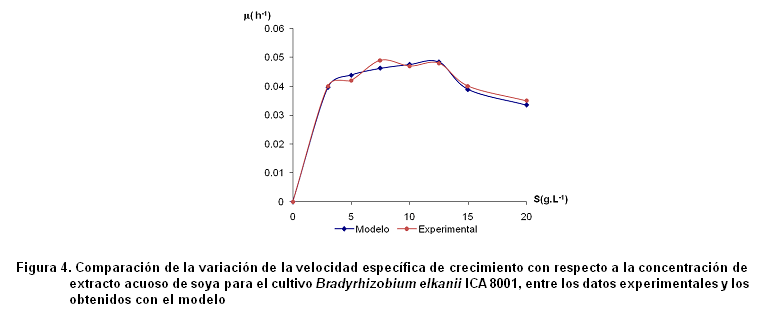
El modelo correlaciona con un valor r2=0,923, los valores experimentales de µ y los obtenidos por el modelo propuesto.
Los parámetros cinéticos calculados permitieron caracterizar la cepa ICA 8001 frente a los sustratos evaluados. El valor de Ks obtenido para la melaza es inferior al que se obtuvo para el extracto acuoso de soya, lo que explica una mayor afinidad de la cepa por el primer sustrato.
En cuanto al fenómeno de inhibición presente en ambos sustratos, se pudo observar que en el caso de la melaza fue más marcado, ya que la caída en los valores de m fue mucho más abrupta que la relacionada con el extracto acuoso de soya (Figuras 1 y 4). El cálculo de la constante de inhibición del sistema (Ki) permitió fundamentar matemáticamente este hecho, pues el valor para la melaza fue superior que el correspondiente al extracto acuoso de soya. Este comportamiento pudo estar relacionado con la presencia de mayores concentraciones de carbohidratos o de otros elementos antinutricionales en la melaza, en comparación con el extracto acuoso de soya.
Los modelos matemáticos propuestos para explicar el comportamiento de la población de Bradyrhizobium elkanii ICA 8001, frente a estos componentes del medio de cultivo, mostraron valores altos de R2, evidenciando la calidad del ajuste de estos a los datos experimentales.
En general, el trabajo realizado aporta conocimientos acerca de la fisiología de Bradyrhizobium, pues evidencia la relación entre la composición del medio de cultivo y los requerimientos nutricionales del microorganismo, así como el diseño de un medio de cultivo puede contribuir a la obtención de inoculantes más efectivos que contengan elevadas concentraciones celulares.
Referencias
1. Treto, E.; García, M.; Martínez, R. y Febles, J. M. Avances en el manejo de los suelos y la nutrición orgánica. [Consultado 4-2006]. Disponible en: <http://www.laneta. apc.org/desal/spip/article.php3?id_article=29>.
2. Bernal, G.; Illanes, A. y Ciampi, L. Isolation and partial purification of a metabolite from a mutant strain of Bacillus sp. Electronic Journal of Biotechnology, 2002, vol. 5 no. 1.
3. Núñez, R. Obtención, caracterización y aplicación de un bioproducto bacteriano para la biorremediación de derrames de hidrocarburos. [Tesis de doctorado]; Universidad de La Habana; 2003, 100 p.
4. Kato, K.; Watanabe, K. y Arima, Y. Rhizobium proliferation supporting substances in seed exudates of common bean (Phaseolus vulgaris L.). Soil Sci. Plant Nutr., 2005, vol. 51, p. 905-910.
5. Nápoles, M. C.; Martínez, J.; Costales, D.; Gómez, G. y Somers, E. Efecto del medio de cultivo en la multiplicación celular de Bradyrhizobium elkanii. Cultivos Tropicales, 2006, vol. 27, no. 1, p. 35-38.
6. Otero, M. A.; Reyes, A.; Carrera, E. y León, M. A. Composition and properties of cane molasses from northeastern Cuba. Int Sugar Journal, 1993, vol. 95, no. 1129, p. 4-8.
7. Smith, K. Advances in Feeding Soybean Meal. [Consultado 6-2005]. Disponible en <http://www.soymeal.org/ksmith1.html>.
8. Batista, C. Optimización de la producción de tensoactivo a partir de una cepa marina. [Tesis de Diploma]; ISPJAE; 2003, 50 p.
9. Nápoles, M. C. Inducción de la nodulación en soya (Glycine max (L.) Merrill) por Bradyrhizobium sp. Influencia del medio de cultivo. [Tesis de Doctorado]; Universidad de La Habana, 2003.
10. Dubois, M.; Gilles, K. A.; Hamilton, J. K.; Rebers, P. A. y Smith, F. Colorimetric method of determination of sugars and related substances. Anal. Chem., 1956, vol. 28, p. 350.
11. Nelson, N. A photometric adaptation of the Somogyi method for the determination of glucose. J. Biol. Chem., 1944, vol. 153, p. 375-380.
12. Paneque, V. M. y Calaña, J. M. Manual de técnicas analíticas para la caracterización de abonos y suelos. La habana:Instituto Nacional de Ciencias Agrícolas, 2002.
13. Monod, J. Recherches sur la croissance des cultures bacteriennes, These de Docteur 6s Sciences Naturelles, 1941, no. 2847, Paris.
14. Haq, I. U.; Ali, S. y Iqbal, J. Effect of volume of culture medium on enhanced citric acid productivity by a mutant culture of Aspergillus niger in stirred fermentor. Letters in Applied Microbiology, 2003, vol. 36, p. 302-306.
15. Furlan, S.; Schneider, A.; Merkle, R.; Carvalho-Jonas, M. F. y Jonas, R. Formulation of a lactose-free, low-cost culture medium for the production of a-D-galactosidase by Kluyveromyces marxianus. Biotechnology Letters, 2004, vol. 22, no. 7, p. 589-593.
16. Hiroko, R.; Megumi, M.C y Dantas, M C. Growth and antibacterial activity of Lentinula edodes in liquid media supplemented with agricultural wastes. Electronic Journal of Biotechnology, 2005, vol. 8, no. 2.
17. Colectivo de autores. Manual de derivados de la caña de azúcar. 2da. Ed. México D.:F.GEPLACEA/PNUD. 1990, p. 59-67.
18. Madigan, M. T. Martinko, J. M. y Parker, J. Brock Biology of microorganisms. 10 ed. Prentice Hall:Pearson Education, 2003.
19. Gómez, G. Algunos factores influyentes en la producción de PHB a escala de laboratorio. [Tesis de Diploma], Facultad de Biología. Universidad de La Habana, 2003, 52 p.
20. Beaulieu, M.; Beaulieu, Y.; Melinard, J.; Pandian, S. y Goulet, J. Influence of ammonium salts and cane molasses on growth of Alcaligenes eutrophus and production of Polyhydroxybutyrate. Applied and Environmental Microbiology, 1995 vol. 61 no. 1, p.165-169.
Recibido: 8 de febrero de 2008
Aceptado: 6 de noviembre de 2008
