










 Servicios personalizados
Servicios personalizados Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Los agroecosistemas son sistemas intervenidos de forma antrópica, es decir, su origen y mantenimiento están asociados a la actividad del hombre, quien ha modificado la naturaleza para obtener bienes y servicios (Sans, 2007). Están formados por componentes biológicos que pueden dividirse, según su función, en productores, consumidores y detritívoros o descomponedores. Estos componentes biológicos presentan una serie de interacciones y relaciones que explican sus funciones. Cuando alguno de estos componentes no funciona de manera adecuada, se pierden sus relaciones estructurales y sus interrelaciones funcionales con los demás componentes del sistema biológico, y se altera su integridad ambiental.
Ello ocurre cuando son sobrexplotados por la intervención humana y, como consecuencia, surge la degradación y la transformación del paisaje natural, lo que conlleva la pérdida de la biodiversidad. Según Zamar et al. (2015), los sistemas agropecuarios han sufrido transformaciones en su estructura y función, por su simplificación productiva, la intensificación en las prácticas agrícolas y la reducción de la heterogeneidad espacial, lo que genera una intensa degradación del medio biofísico y la disminución de la sustentabilidad y la sostenibilidad.
En los agroecosistemas se presentan relaciones tróficas no lineales, ya que el flujo de energía sigue diferentes rutas (Griffon, 2008). Dichas interacciones tróficas son un elemento clave de la dinámica comunitaria en estos y ocurren no solo entre niveles tróficos adyacentes, como cultivos y herbívoros, sino también como efectos indirectos a través de distantes niveles tróficos (Martínez-Romero y Leyva-Galán, 2014). Por eso, los estudios sobre redes tróficas son fundamentales para entender el funcionamiento de los ecosistemas.
Según Pimm et al. (1991), las redes tróficas son una variante del enfoque de sistema; se representan en términos de trofoespecies, que pueden ser basales (sin presas), intermedias (con presa y predador) y de tope (sin predador en la comunidad). Desde el punto de vista de su estructura o topología, están constituidas por nodos, eslabones y niveles tróficos que describen la diversidad, las relaciones alimentarias, la estabilidad y los procesos que ocurren dentro de un ecosistema (Pedroza et al., 2016).
El objetivo de este estudio fue definir las relaciones tróficas existentes en el agroecosistema de una granja en la región Sabanas, Sucre, Colombia
Materiales y Métodos
Localización. El estudio se realizó en la granja El Perico, ubicada en la región Sabanas (Departamento de Sucre), sobre las coordenadas 9º 12’ 41.7” N-75º 24’ 09.7” W (fig. 1). La temperatura promedio de la zona es de 26,8 º C, con una precipitación que varía entre 1 000 y 1 200 mm/año y una humedad relativa de 77 %. Inicialmente se hizo un reconocimiento del área, para identificar los sitios de muestreo donde se podía observar presencia de individuos, tanto en la cobertura vegetal como en el suelo.
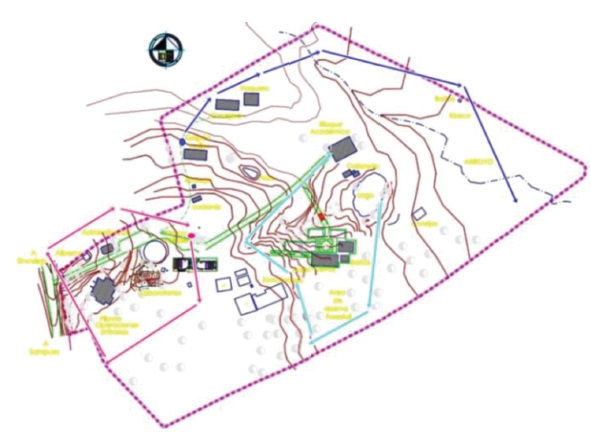
Para la obtención e identificación de los individuos se utilizó información secundaria de la zona: para los anuros, la de Cardozo y Caraballo (2007); para las hormigas, la de Bertel (2015); y para los murciélagos. la de Sampedro et al. (2007). Se incluyeron observaciones e identificaciones realizadas entre los años 2014-2017, en jornadas matutinas y vespertinas de trabajo, siempre teniendo en cuenta los límites de la granja.
El resto de la información se recolectó in situ: bovinos y ovinos, avistamientos y observaciones con el uso de fotografías, binoculares y claves dicotómicas (aves, reptiles); descripciones morfológicas, caracteres diagnósticos y el uso de guías botánicas (plantas); claves dicotómicas (mamíferos, zarigüeya, oso perezoso, dípteros, himenópteros, odonatos y coprófagos) y capturas directas (invertebrados descomponedores). Posterior a la identificación, los individuos se agruparon según su función en el agroecosistema (Pimm et al., 1991). La flora y los individuos vertebrados se identificaron hasta especie, mientras que los invertebrados, como algunos insectos, solo hasta Orden.
Se evaluó la riqueza de especies, tanto animal como vegetal, de acuerdo con lo planteado por Moreno (2001), quien utiliza la riqueza específica como la forma más sencilla de medir la diversidad. La riqueza de la red trófica se asimiló como equivalente del número de trofoespecies definidas. Se estableció la estructura trófica del agroecosistema organizando los grupos en función de sus estrategias tróficas más generales, y la clasificación se basó en consulta extensa de la información y los datos bibliográficos para conocer los hábitos alimenticios (tabla 1).
Análisis de los datos. Para visualizar el flujo de energía de la granja se diseñó una red trófica mediante un modelo estático, basado en la estructura de una matriz binaria, que indica la presencia (1) o ausencia (0) de conexiones. Se registraron las trofoespecies dentro del agroecosistema y las interacciones tróficas que pueden ocurrir entre ellas. Para este modelo de red se empleó el programa de libre acceso Gephi 091, un software diseñado para la confección de gráficos, manejo de la información, estructuras, formas y patrones, utilizado por Pedroza et al. (2016). Para la interpretación de este modelo se midieron los siguientes atributos: conectancia (C=L/S2) y densidad de uniones (L/S), según Pimm et al. (1991) y Dunne y Williams (2009), respectivamente.
Resultados y Discusión
Los individuos se identificaron en seis órdenes (Lepidoptera, Polydesmida, Araneae, Haplotaxida, Pulmonata y Blattodea), dos familias (Calliphoridae, Aphididae), ocho géneros (Camponotus, Labidus, Crematogaster, Ectatomma, Dorymyrmex, Forelius, Pheidole, Solenopsis) y ciento sesenta y seis especies (tabla 1).
Basado en el hábito alimenticio, se establecieron 11 categorías tróficas: productores (organismos autótrofos), herbívoros (organismos que se alimentan de plantas), nectarívoros (organismos que se alimentan del néctar de las plantas), granívoros (organismos que se alimentan de semillas), insectívoros (organismos que se alimentan de insectos), carnívoros (organismos que se alimentan de organismos herbívoros), omnívoros (organismos que se alimentan de plantas y animales), descomponedores (organismos heterótrofos que se alimentan de detritos), hematófagos (organismos que se alimentan de sangre), frugívoros (organismos que se alimentan de frutas) y carroñeros (organismos que se alimentan de material en descomposición), distribuidas en 48 trofoespecies (tabla 2). En términos de número de especies, los grupos más diversos fueron las trofoespecies omnívoras e insectívoras y el grupo menos variado fue el de las trofoespecies nectarívoras.
Tabla 2 Organización trófica de las categorías en la red trófica y número de trofoespecies por cada categoría.
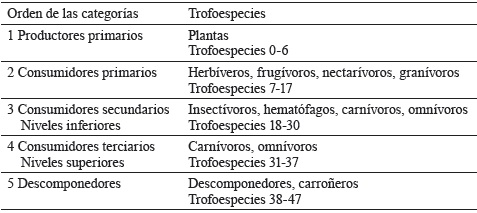
Los hábitos alimenticios antes mencionados se agruparon en: productores (gramíneas, maderables, maderables leguminosas, frutales, arbustos, plantas ornamentales y plantas acuáticas), consumidores primarios (herbívoros, granívoros, frugívoros y polinizadores), consumidores secundarios (insectívoros, hematófagos, omnívoros y carnívoros), consumidores terminales (carnívoros y carroñeros) y descomponedores (hongos, formícidos, diplópodos, dípteros, coleópteros, blatodeos, isópteros, anélidos y gasterópodos). Al agrupar varias especies en una trofoespecie, se asumió como criterio la función trófica de estos, o sea, que compartan presa y/o predador en el sistema. En muchos aspectos, es fundamental considerar la función y conocer qué hacen las especies constituyentes, en lugar de evaluar si están presentes o no en el agroecosistema (Pedroza et al., 2016).
La red trófica que integra la comunidad del agroecosistema presentó 156 conexiones, que representaron la relación predador-presa de los componentes biológicos (fig. 2). Este valor se considera alto, si se compara con lo informado por Roubinet (2016) quien observó solo 77 conexiones. La diferencia pudiera ser consecuencia del tipo de agroecosistema, ya que este último se desarrolló en un monocultivo, que según Altieri (1995) se caracteriza por presentar menos interacciones tróficas.

El tamaño máximo de la red fue de siete niveles tróficos y el mínimo de dos, por lo que la red tenía un tamaño medio de 4,5; ello es inusual en redes tróficas terrestres y acuáticas, que normalmente cuentan con tres o cuatro niveles (Pimm et al., 1991). Esta condición es consecuencia del elevado número de trofoespecies y la presencia de una gran cantidad de consumidores. Si se considera el tamaño de la red, se puede inferir que se trata de una red vulnerable, por la gran pérdida de energía que representan muchos niveles tróficos. Sin embargo, en términos de robustez, se considera una red altamente robusta, ya que si se retira el 50 % de la comunidad de productores la red se mantendría sostenible (Dunne et al., 2002). La red trófica se fortaleció por contar con siete trofoespecies basales que incluyeron 90 especies, encargadas de asimilar energía y pasarla a la mayoría de los consumidores. Adicionalmente, la red trófica contó con la presencia de 11 trofoespecies consumidoras primarias, que incluyeron 23 especies, lo que generó una alta redundancia ecológica en términos de relaciones funcionales; ello garantizó el flujo de energía hacia los niveles superiores y la circulación de nutrientes (De Ruiter et al., 2005).
Las trofoespecies predadoras y presas más importantes, de acuerdo con el número de interacciones, se muestran en las figuras 3 y 4. Su extinción o exclusión pudiera romper las relaciones entre los conjuntos de especies, y ello puede perjudicar seriamente la integridad de los ecosistemas (Perfecto et al., 2014).

Figura 3 Número de presas para las principales especies predadoras en la comunidad. El mayor valor corresponde a la trofoespecie más generalista de la comunidad.
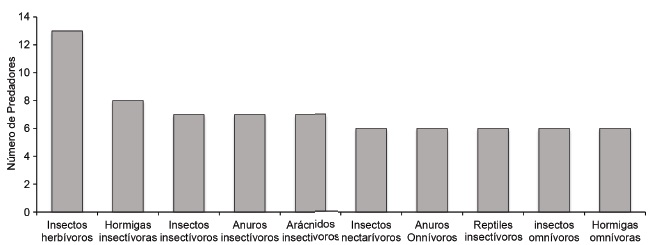
Figura 4 Número de predadores para las principales especies presa en la comunidad. El mayor valor corresponde a la trofoespecie que más transporta material a los niveles tróficos superiores en la comunidad
Roubinet et al. (2018) consideran que la trofoespecie 9 (insectos herbívoros) es industrial, ya que presenta 13 predadores que transfieren energía a los consumidores superiores; y la ausencia de estos insectos afectaría, a su vez, la estabilidad del sistema (Macfadyen et al., 2009).
Las otras trofoespecies, en su mayoría, tuvieron en promedio de tres a siete predadores, con excepción de las trofoespecies 8 (reptiles herbívoros 2), 10 (murciélagos frugívoros), 21 (murciélagos hematófagos) y 31 (murciélagos insectívoros) que tuvieron solo una salida en el sistema. Estos resultados se analizaron mediante la red trófica del sistema y demostraron la importancia de cada trofoespecie. Igualmente, sobresalió la presencia del mamífero Didelphis marsupialis (fig. 3, trofoespecie 37) como especie reguladora de la comunidad, por presentar 11 presas.
Dentro de las trofoespecies intermedias y las trofoespecies de tope, hubo una variedad de hábitos alimenticios; de estos, predominaron las relaciones de grupos insectívoros que abarcaron ocho trofoespecies e incluyeron 17 taxones y los omnívoros que abarcaron seis trofoespecies y 28 taxones. De acuerdo con lo señalado por Roubinet et al. (2018), esto garantiza un flujo de energía hacia las trofoespecies de tope.
Entre los descomponedores de la red se hallaron los artrópodos, que desempeñan un papel importante en los procesos de degradación de los restos de origen vegetal. Se crearon acciones mutualistas con microorganismos, al contar con bacterias o protozoos simbióticos permanentes. Los grupos más conocidos son las termitas (isópteros) y las cucarachas (blatodeos). En este sentido, el estudio de la entomofauna necrófaga y, en especial, la coprófaga, tiene un gran interés ecológico y económico, ya que la acción de fragmentación y enterramiento de los restos orgánicos favorece el desarrollo de los microorganismos y de las hifas micelianas que participan en la desintegración (Galante y García, 1997). En las áreas de pastos, como en el caso de la parcela A, esta acción reviste además un interés económico (Tovar et al., 2016), ya que evita la acumulación de excrementos sobre el suelo, lo cual disminuye la disponibilidad de las pasturas y su consumo por los animales.
En la mayoría de los niveles tróficos se notó la presencia de insectos. De acuerdo con sus características tróficas variables, se reconoce que es imprescindible su presencia en el sistema. Por ejemplo, las hormigas realizan importantes funciones, tales como el movimiento de suelo, el de semillas y la depredación; además han demostrado ser uno de los grupos de insectos más sensibles a los cambios ambientales (Zamar et al., 2015).
Para el caso de la densidad de uniones se halló un valor de 3,25, similar al encontrado por Roubinet (2016) en un monocultivo de cereales (3,6), pero diferente al de un agroecosistema de café (Perfecto et al., 2014). De ello se infiere que este atributo es muy variable y depende directamente de las trofoespecies involucradas en el estudio y de las funciones y relaciones que estas están realizando en el sistema. Al comparar los agroecosistemas mixtos con el monocultivo, hubo una diferencia significativa en cuanto a la diversidad y la función de las especies, lo que puede ser una característica poco adecuada para confrontar redes de diferentes tamaños y resoluciones (Macfadyen et al., 2009). También al contrastar individualmente los valores de densidad, se infiere una vinculación trófica media entre los nodos formados, ya que es probable que exista una alta diversidad funcional dentro de las distintas trofoespecies establecidas (Pimm et al., 1991).
La conectancia de la red fue de 0,13, valor que es considerado una conectividad media en el sistema (Dunne y Williams, 2009). Estos autores señalan que los valores medios de conectancia oscilan entre 0,1 y 0,15. Los valores del presente estudio difirieron de las conectancias informadas por Roubinet (2016) y por Macfadyen et al. (2009), quienes obtuvieron 0,36 y 0,29, respectivamente.
Estos autores estructuraron redes tróficas cuantitativas, que también consideraban estables por darle importancia a interacciones particulares de diferentes individuos, en dependencia de la fuerza de dichas interacciones. La diferencia en los resultados se debe a la cantidad de trofoespecies presentes en los diferentes agroecosistemas, ya que, en sus casos, estos eran sistemas en monocultivo y orgánico, respectivamente; los cuales cuentan con menor número de trofoespecies, por lo que ocurren menos interacciones. Del valor de conectancia se infiere que en el sistema habrá un mayor grado de recuperación ante perturbaciones ambientales (Francis et al., 2003).

