










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión impresa ISSN 1010-2752
Rev. Protección Veg. vol.28 no.2 La Habana mayo-ago. 2013
ARTÍCULO ORIGINAL
Biología y tabla de vida de Tamarixia radiata Waterston bajo condiciones controladas
Biology and life table of Tamarixia radiata Waterston under controlled conditions
Heyker L. Baños Díaz, Ileana Miranda, María de los A. Martínez
Dirección de Sanidad Vegetal. Centro Nacional de Sanidad Agropecuaria (CENSA). Apartado 10, San José de las Lajas, Mayabeque, CP 32700, Cuba. Correo electrónico: hlellani@censa.edu.cu.
RESUMEN
Con el objetivo de determinar bajo condiciones controladas, la duración del ciclo de vida, la sobrevivencia, proporción sexual, longevidad, fecundidad y otros atributos biológicos y poblacionales de Tamarixia radiata Waterson, se estableció una cría sobre Diaphorina citri Kuwayama en Murraya paniculata (Jack) a 70±10% de HR, 28±1°C y 14 :10 LD. El periodo de pre-momificación y el ciclo de desarrollo fueron de 3,08 y 7,27 días respectivamente, para un ciclo de vida total de 25,62 con una sobrevivencia promedio de 18,35 días. El parasitoidismo superó el 70% con una emergencia de 72,27%, a su vez, la tasa de ovoposición fue de 24 huevos/hembra/día, y una longevidad de 23 días como promedio para la especie, siendo mayor en las hembras. El tiempo de fecundidad de la hembra de T. radiata fue 0,94 días y la tasa de crecimiento reproductivo 333,77 huevos/día. Por otra parte, los parámetros poblacionales como la Tasa intrínseca de incremento (rm) y la Tasa finita de crecimiento (l) presentaron valores de 0,14 y 1,02, respectivamente. La Tasa neta reproductiva (R0) tuvo un valor estimado de 1,55 (hembras hijas/hembras madres). Estos parámetros fueron comparados con los de su hospedante D. citri, confirmando que T. radiata es un potencial agente de control biológico para D. citri.
Palabras clave: Diaphorina citri, control biológico.
ABSTRACT
With the aim of determining the duration of the life cycle, survival, sex ratio, longevity, fecundity and other biological and population attributes of Tamarixia radiata Waterson, it was reared on Diaphorina citri Kuwayama on Muraya paniculata (Jack) plants at 70±10% of HR, 28±1p C and 14 :10 LD. The period of pre-mummification and the development cycle was of 3,08 and 7,27 days, respectively, for a total life cycle of 25,62 with a survival of 18,35 days. The parasitism exceeded 70% with an emergence of 72,27%.On the oher hand, the rate of oviposition was 24 eggs / female / day, and a longevity of 23 days, which was higher in females. The time of fecundity for the T. radiata female was 0,94 days and the reproductive growth rate of 333,77 eggs/day. The population parameters such as the intrinsic rate of increase (rm) and the finite rate of increase (l) showed values of 0,14 and 1,02 respectively, and the net reproductive rate (R0) had an estimated value of 1,55 (daughters female/female mothers). These parameters were compared with those of its host D. citri, and they reaffirmed T. radiata as a potential biological control.
Key words: Diaphorina citri, biological control.
INTRODUCCIÓN
Tamarixia radiata Waterston (1922) es el principal enemigo natural del psíllido asiático de los cítricos (Diaphorina citri Kuwayama) (1, 2, 3).
Es un parasitoide nativo de la India que fue utilizado por diferentes países en programas de control biológico clásico con muy buenos resultados, pues disminuyó sustancialmente las poblaciones de fitófago, así como la enfermedad que estos trasmiten, comportándose superior a otros parasitoides como Diaphorencirtus aligarhensis debido a altas tasas de parasitación del psílido y el rápido establecimiento en nuevas áreas (4,5).
La liberación de T. radiata revivió la industria de los cítricos en la Isla de la Reunión, después de su introducción desde la India en 1978 (6) y el parasitoide causó la disminución sustancial de las poblaciones de D. citri en la Isla de Guadalupe a un año de la liberación (4, 6, 7, 2).
Además de parasitar los estadios de tercero a quinto instar, las hembras adultas del parasitoide se alimentan de las ninfas de primero y segundo instar (8, 5). T. radiata puede obtener proteína para el desarrollo del huevo alimentándose de la hemolinfa de las ninfas de psílidos, a los cuales accede a través de ovipositor (9). Una sola hembra de T. radiata es capaz de matar a más de 500 psílidos por una combinación de alimentación y de parasitoidismo. Las hembras también se alimentan de melaza excretada por los psílidos (9). Los adultos son fuertemente atraídos por las brillantes luces fluorescentes (5) y para la ubicación del hospedante las hembras se basan fundamentalmente en señales olfativas (10).
Dado que las características biológicas de un parasitoide pueden variar dependiendo de la población, la especie y biotipo del insecto huésped, la planta hospedera y las condiciones ambientales, resulta importante conocer los parámetros bioecológicos que permitan evaluar las características intrínsecas de las poblaciones de plagas y sus enemigos naturales (11).
La biología y las tablas de vida constituyen un componente importante para estimar los valores vitales de una población de insectos. Esta metodología es utilizada para estudiar la dinámica poblacional de una especie en particular, para seleccionar enemigos naturales y evaluar el impacto sobre una plaga por parte de un insecto benéfico (12).
Estudios preliminares de laboratorio demostraron que T. radiata tiene un gran potencial para el control de D. citri ya sea por parasitoidismo o por depredación de las ninfas más jóvenes y que además posee una alta capacidad reproductiva (9). Sin embargo, hasta el momento son escasos los antecedentes sobre sus parámetros de vida y la relación con los de su hospedante.
Este trabajo tuvo como objetivo determinar parámetros biológicos y de vida de Tamarixia radiata y su relación con los parámetros de vida de su hospedante.
MATERIALES Y MÉTODOS
Biología de T. radiata sobre D. citri bajo condiciones controladas
Para el estudio biológico de T. radiata se dispusieron 30 frascos de cristal de 30 ml de capacidad, con agua en su interior y la boca cubierta con papel de aluminio; sobre el cual se practicó un orificio por donde se introdujo una hoja de M. paniculata (Jack) con tres foliolos .Los frascos fueron introducidos en otro de mayor capacidad (500 ml) y posteriormente se inoculó la hoja con una ninfa de D. citri de quinto instar. Paralelamente se individualizaron 30 parejas de T. radiata una por cada frasco, para garantizar el apareamiento, las cuales se alimentaron con solución de miel de abejas y agua (50%). A las 48 horas la hembra fecundada fue liberada en los frascos, los cuales fueron colocados en una incubadora Friocell bajo condiciones de 14 horas luz, 70%±10% humedad relativa y temperatura constante de 28oC±1oC.
Transcurridas 24 horas, se retiró el parasitoide y se iniciaron las observaciones diarias para determinar los cambios de coloración en las ninfas producto del parasitoidismo, la aparición de las estructuras de fijación (fase de pre-momificación) y el momento de la emergencia del adulto, los cuales se mantuvieron bajo las mismas condiciones hasta la muerte. Se calculó la duración del ciclo de desarrollo, duración del ciclo vida, longevidad de la especie y relación sexual.
Para determinar la sobrevivencia se conformaron 3 grupos de 20 individuos, cada uno, compuesto por 10C y 10X con 24 horas de emergidos; los cuales fueron alimentados de tres formas diferentes (miel + agua; agua + pólen y solamente agua). Se registró la fecha de emergencia y la fecha de muerte para cada individuo y se evaluó el efecto del alimento sobre la sobrevivencia de los adultos. Para ello, los datos obtenidos fueron procesados mediante un ANOVA simple y las medias se compararon según Tukey para pW 0,01. Para el análisis estadístico se utilizó el paquete SAS System para Windows, versión 8.
Tabla de vida
Con el propósito de estudiar los parámetros poblacionales de un grupo de individuos nacidos dentro de un mismo intervalo de tiempo (cohorte), desde su nacimiento hasta la muerte se elaboró la tabla de vida para 30 hembras de T. radiata, las cuales se observaron diariamente.
Los datos obtenidos de la biología de la especie se utilizaron para calcular la tasa de sobrevivencia específica (lx), fecundidad específica (mx) diaria, tasa neta de reproducción (R0), tiempo medio generacional (T), tasa intrínseca de incremento natural (rm), doble tiempo generacional (DT), y tasa finita de incremento (l).
RESULTADOS Y DISCUSIÓN
Biología de T. radiata sobre D. citri bajo condiciones controladas
Tamarixia radiata depositó los huevos en la parte ventral cercano a las coxas del segundo y tercer par de patas de las ninfas de tercero a quinto instar de D. citri. El huevo fue de color amarillo y de forma ovalada, y dio lugar a una larva, la cual se fue alimentando de la ninfa hasta que consumió todo el interior de la misma. Las ninfas parasitadas permanecen prácticamente inmóviles y su coloración cambió a pardo rojizo. Una vez que la larva pasó a pupa, segregó una sustancia con la cual se fijó la ninfa a la hoja; este momento es conocido como período de pre-momificación.
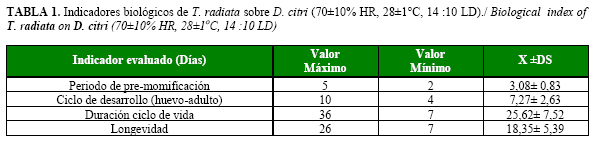
Durante nuestro estudio el periodo de pre-momificación ocurrió a los tres días, mientras que la emergencia de los adultos se registró a los seis días de iniciarse el parasitoidismo, alcanzando un 70%; aunque sólo el 72,27% de los parasitoides emergió, valor muy alentador si tenemos en cuenta que en el año 2001 los niveles de parasitoidismo informados en la Isla de la Juventud no excedieron el 37% (13).
Los resultados obtenidos relativos a la duración del ciclo de desarrollo y el ciclo de vida del insecto se encontraron dentro de los informados para la especie bajo estas condiciones, obteniéndose como promedio siete días y 26 días, respectivamente.
Según Fuavergue y Quilici (14) a 27oC, la fase de pre-momificación de este parasitoide es de 3,6 días y el ciclo de desarrollo de 9,1 días. Este último valor resultó superior en dos días al obtenido en este trabajo, lo cual está en correspondencia con los valores de temperatura prefijados. Por su parte, Chien (2), planteó que a 25oC el período de pre-momificación estaba alrededor de 4,9 días y el ciclo de desarrollo aumentó a 12 días. A su vez Gómez (15), bajo temperaturas entre 25-30°C, planteó que la duración del ciclo de desarrollo fue de 8,33-11 días.
Según Wang et al. (16) las variaciones de temperaturas pueden aumentar o disminuir la duración de los parámetros biológicos (ciclo de desarrollo, sobrevivencia y reproducción) de las especies de la familia Eulophidae. Estos autores también concluyeron que hembras de esta familia, pueden, a intervalos de temperatura entre 25-30°C, parasitar un mayor número de huéspedes y presentar un mayor índice fecundidad, en relación con los valores obtenidos a otras temperaturas.
Varios autores señalaron la influencia de la temperatura sobre la velocidad de desarrollo en eulófidos (17,18,19). Hondo et al. (20), evaluaron sobre la base de la tolerancia térmica del desarrollo, la reproducción y la eficiencia de siete especies de eulófidos y llegaron a la conclusión de que todas las especies analizadas se adaptaron a las altas temperaturas; sin embargo, se produjo una reducción de la capacidad reproductiva entre 15 y 20°C. De ahí la importancia de tener en cuenta este parámetro a la hora de elegir un agente de control biológico.
Por otra parte, el hecho de que la especie alcance la fase adulta en un breve periodo de tiempo, permite el desarrollo de hasta tres generaciones del parasitoide por una del hospedante, elemento básico de gran importancia a tener en cuenta para establecer una estrategia de control.
En el presente estudio, la longevidad de T. radiata estuvo alrededor de los 18 días como promedio, con valores máximos de hasta 26 días (Tabla 1), esto podría estar dado por la suplementación en su alimentación con ninfas de los primeros instares, pues se plantea que T. radiata tiene la habilidad de parasitar y alimentarse de las ninfas de D. citri hasta un 20% (15,21).
Chu et al. (21), señalaron una longevidad de las hembras de 23,6 y 14,8 días para los machos, con 80% de parasitoidismo y 71,2% de emergencia; a su vez Gómez (16), obtuvo valores de parasitoidismo entre 84,17% - 72,50% y de emergencia entre 88,41- 78,81%; valores cercanos a los obtenidos en este estudio.
La sobrevivencia de los adultos bajo las condiciones impuestas no excedió los dos días (Figura 1) difiriendo significativamente de la alcanzada en las jaulas de cría. Estos resultados indicaron que esta especie de parasitoide no está capacitada para soportar condiciones de estrés ambiental en relación con un déficit temporal de alimento y aislamiento, las cuales pueden influir en la sobrevivencia del insecto, que posee distribución agregada (13).
Por otra parte, la sobrevivencia fue mayor en las hembras, lo que evidenció que los machos de esta especie son más sensibles al estrés provocado por y la carencia de otras fuentes de alimento.
Fuavergue y Quilici (14), obtuvieron que la sobrevivencia disminuyó de 23 días (con miel y agua) a 2 días en ausencia de recursos azucarados suministrando solamente agua, lo cual reafirma la importancia de este recurso para esta especie, como para otros parasitoides.
Resultados similares a los alcanzados en nuestro estudio, fueron obtenidos por diversos grupos a nivel internacional, donde se destacó que la sobrevivencia de T. radiata es dependiente de la temperatura y de la presencia de alimentos.
La longevidad de los adultos alimentados con miel, aumentó notablemente en comparación con aquellos a los que solamente se le ofreció agua sin ninguna otra reserva de alimento (15).
Tabla de vida
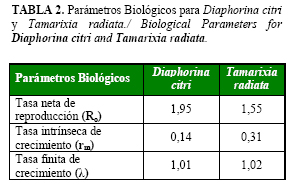
El tiempo de fecundidad para las hembras de T. radiata fue de 0,94 días. En este tiempo, la hembra de T. radiata presentó una Tasa de crecimiento reproductivo (GRR) de 337,77 huevos/día y una tasa de ovoposición diaria de 11,26 huevos/hembra/día. Por su parte la Tasa neta reproductiva (R0) tuvo un valor estimado de 1,55 (hembras hijas/hembras madres). Este resultado mostró que esta especie hace un aporte de casi dos hembras hijas por cada hembra madre en una generación, dato muy alentador si tenemos en cuenta que las hembras son las encargadas de controlar la plaga y mantener la permanencia del parasitoide en el campo.
Por otra parte, el tiempo medio generacional fue de 21,91 días y el tiempo en que se duplica la generación (doble tiempo generacional) fue de 34,65 días; teniendo en cuenta esto, es apropiado decir que se lograrían hasta 11 generaciones anuales del parasitoide, antecedente a tener en cuenta durante la realización de la cría masiva.
Los valores de la Tasa intrínseca de incremento (rm) y la Tasa finita de crecimiento (l) fueron de 0,31 y 1,02, respectivamente. Ambos parámetros están genéticamente determinados, el primero refleja la capacidad potencial de multiplicación de la población, mientras que el segundo es utilizado como punto de partida para el manejo de la población, para incrementar o reducir la dispersión o diseminación de una especie, ya que está regido por el ambiente y el ciclo de vida de las mismas (22).
En el caso de T. radiata, la tasa intrínseca de crecimiento (rm) que se obtuvo, resultó un buen indicador de la temperatura a la que el crecimiento de la población fue favorable. Este parámetro posee efectos en el desarrollo, reproducción y supervivencia de este parasitoide.
Estos valores difieren de los obtenidos por Gómez (23), quien señaló una tasa neta de reproducción de 58,63 (hembras hijas/hembras madres), una tasa intrínseca de crecimiento de 0,34 y una tasa finita de crecimiento de 1,40 (T:30±2°C, HR:70±10% y LD:14:10).
Por su parte Chu y Chien (21), informaron una tasa neta de reproducción, tasa intrínseca de crecimiento y tasa finita de crecimiento de 0,31 días, 11,37 días y 1,28, respectivamente.
Como se puede observar no existe coincidencia de estos valores con los resultados obtenidos durante este estudio, lo cual ratifica que las características reproductivas y de fecundidad de las especies de insectos están influidas por las condiciones imperantes en el medio en que se desarrollan estos.
Otros estudios con eulófideos destacaron la importancia de la temperatura en el desarrollo y la reproducción (24,25). Castillo et al. (25), obtuvieron que la fecundidad de Quadrastichus haitiensis aumentó cuando la temperatura fue de 30°C; sin embargo, la duración del desarrollo de huevo a adulto disminuyó; mientras que a temperaturas por debajo 15 °C no ocurrió el desarrollo de estos parasitoides.
Acosta y O'Nell (24) a temperaturas de 15, 24 y 30°C estudiaron las tablas de vida y fertilidad de poblaciones de eulóphidos provenientes de Honduras, Colombia y México, y encontraron biotipos adaptados a bajas temperaturas, lo cual es importante para la comprensión de la biogeografía de los parasitoides, sus hospedantes y plantas hospederas, así como la variabilidad de la dinámica en las poblaciones en regiones geográficamente diferentes. Dicha variabilidad podría servir como una base para la selección de poblaciones adaptadas a condiciones climáticas comunes en las regiones en las que se utilizaran para controlar el insecto blanco.
Por su parte, Viggiani et al. (26) señalaron también la comprensión de los parámetros biológicos de eulófidos bajo diferentes regímenes de temperatura, y comprobaron en condiciones de campo la ineficacia de estos parasitoides cuando las condiciones climáticas fueron más apropiadas para la plaga.
Dado que las características biológicas de un parasitoide pueden variar dependiendo de la población, la especie y biotipo del insecto huésped, la planta hospedera y las condiciones ambientales, resulta importante el conocimiento de los parámetros bioecológicos que permitan evaluar las características intrínsecas de las poblaciones de plagas y enemigos naturales (27).
Si, comparamos los parámetros de fecundidad de T. radiata con los de su hospedante D. citri (Tabla 2), se puede apreciar que la R0 o número de hembras por cada hembra en una generación, fue mayor para D. citri que para T. radiata, indicando con ello su alta capacidad de reproducción cuando es alimentada sobre M. paniculata.
La capacidad de multiplicación de la población (rm) fue mayor para T. radiata que para el psílido asiático de los cítricos, resultados que confirmaron la potencialidad de control que ofrece este parasitoide sobre la plaga. El valor marcadamente superior de la tasa intrínseca de crecimiento poblacional del enemigo natural, sugiere que, utilizándolo en un programa de control biológico, podría mantener en un bajo nivel la población de D. citri.
La tasa finita de crecimiento, fue de 1,02, para T. radiata valor que se considera como indicador de un buen candidato para el control de especies fitófagas (27), y fue mayor al obtenido para D. citri, lo que refuerza aún más la idea de que el parasitoide pudiera mantener en bajos niveles las poblaciones de la plaga.
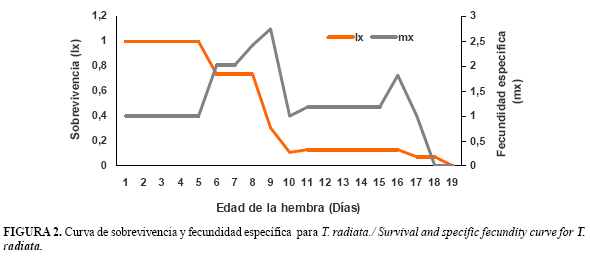
Al analizar la proporcion de individuos sobrevivientes de T. radiata durante su tiempo de vida, los resultados mostraron una reducción significativa de la sobrevivivencia a medida que aumenta la edad de las hembras, simulando una curva de tipo II (Figura 2).
La fecundidad específica (huevos/hembra/día) también disminuyó con el aumento de la edad de las hembras, hasta hacerce 0. Se observó que la mayor fecundidad ocurre en los primeros 9 días de ovoposición, mostrando un pico en el primer día, después de los cuales la ovoposición disminuyó considerablemente (Figura 2).
Gómez (15) informó resultados similares de la sobrevivencia y la fecundidad especifica de T. radiata a 30°C.En ambos casos ocurrió una disminución de estos parámetros después de los 18 días de emergida la hembra demostrando que el periodo de desarrollo de huevo a adulto disminuye a medida que aumenta la temperatura.
Los resultados presentados en este trabajo constituyen los primeros que se obtienen del enemigo natural bajo las condiciones de Cuba y lo ratifican como un agente de control biológico potencial para el manejo de D. citri en la citricultura nacional.
REFERENCIAS
1. Waterston J. On the chalcidoid parasites of psyllids (Hemiptera, Homoptera). Bulletin of Entomological Research. 1922;(13):41-58.
2. Chien Ching-Chin, Chiu Shui-Chen, Ku Shiu-Chih. Department of Applied Zoology Taiwan Agaricultural Research Institute Wufeng, Mass rearing and field release of an eulophid wasp. Taichung 41301, Taiwan, ROC, 2001-05-01.
3. Alemán J, Baños H, Ravelo J. Diaphorina citri y la enfermedad Huanglongbing: Una combinación destructiva para la producción citrícola. Rev Protección Veg. 2007;22(3):154-165.
4. Aubert B, Quilici S. Biological control of the African and Asian citrus psyllids (Homoptera: Psylloidea), through eulophid and parasites (Hymenoptera: Chalcidoidea) in Reunion Island. In Garnsey SM, Timmer LW, Dodds JA (eds.), Proceedings of the 9th Conference of the International Organization of Citrus Virologists, University of California, Riverside. 1984: 100-108.
5. Skelley LH, Hoy MA. A synchronous rearing method for the Asian citrus psyllid and its parasitoids in quarantine. Biological Control. 2004;(29):14-23.
6. Etienne J, Quilici S, Marival D, Franck A. Biological control of Diaphorina citri (Hemiptera: Psyllidae) in Guadeloupe by imported Tamarixia radiata (Hymenoptera: Eulophidae). Fruits. 2001;56(5):307-315.
7. Hoy MA, Nguyen R. Classical biological control of Asian citrus psylla. Citrus Industry. 2001;81:48-50.
8. Chien CC. The role of parasitoids in the pest management of citrus psyllid. Proceedings of the Symposium on Research and Development of Citrus in Taiwan, Taichung, Taiwan, 1995: 245-261.
9. Hoy MA, Nguyen R, Jeyparakash A. Classical biological control of Asian citrus psyllid in Florida. Florida IPM 2006. Disponible en: http://ipm.ifas.ufl.edu/agriculture/citrus/psyllid.shtml. Consultado: 14 marzo 2001.
10.Mann RS, Rouseff Rl, Smoot JM, Castle WJ, Stelinski LL. Sulfur volatiles from Allium spp. affect Asian citrus psyllid, Diaphorina citri Kuwayama (Hemiptera: Psyllidae) response to citrus volatiles. Bulletin of Entomological Research. 2010.
11.Vargas R, Rodríguez S. Manejo de Plagas en Paltos y cítricos. Dinámica de las poblaciones, Capitulo 7. 2008.
12.Crespo DC, La Rossa FR, Lecuona RE. Parámetros poblacionales de Muscidifurax raptor Girault and Sanders (Hymenoptera: Pteromalidae) sobre pupas de Musca domestica L. (Díptera: Muscidae). Boletin Sanidad Vegetal Plagas. 2003;29:243-248.
13.Fernández M, Miranda I. Comportamiento de Diaphorina citri Kuwayama (Hemiptera: Psyllidae). Parte I: Características morfológicas, incidencia y enemigos naturales asociados. Rev Protección Veg. 2005;20(1):27-31.
14.Fauvergue X, Quilici S. Etude de certain paramétres de la biologie de Tamarixia radiata (Waterson, 1922) (Hymenoptera: Eulophidae), ectoparasitoïde primaire de Diaphorina citri Kuwayama (Hemiptera: Psillidae), vectour asiatique du greening des agrumes. Fruits. 1991;46(2):179-185.
15.Gómez ML. Estudos bioecológicos de Tamarixia radiata (Waterston, 1922) (Hymenoptera: Eulophidae) para o controle de Diaphorina citri Kuwayama, 1907 (Hemiptera: Psyllidae). Tesis en Opción al grado científico de Doctor en Ciencias. Piracicaba. Brasil. 2009.
16.Wang X, Liu S, Guo S, Lin W. Effects of host stages and temperature on population parameters of Oomyzus sokolowskii, a larval-pupal parasitoid of Plutella xylostella. BioControl, Dordrecht. 1999;44:391-402.
17.Minkenberg OPJM. Temperature effects on the life history of the eulophid wasp Diglyphus isaea, an ectoparasitoid of leafminers (Liriomyza spp.), on tomatoes. The Annals of Applied Biology, Warwick. 1989;115:381-397.
18.Bazzocchi GG, Lanzonin A, Burgio G, Fiacconi MR. Effects of temperature and host on the pre-imaginal development of the parasitoid Diglyphus isaea (Hymenoptera: Eulophidae). Biological Control, Orlando. 2003;(26):74-82.
19.Duale AH. Effect of Temperature and Relative Humidity on the Biology of the Stem Borer Parasitoid Pediobius furvus (Gahan) (Hymenoptera: Eulophidae) for the Management of Stem Borers. Environmental Entomology, College Park. 2005;(34):1-5.
20.Hondo T, Koike A, Sugimoto T. Comparison of thermal tolerance of seven native species of parasitoids (Hymenoptera: Eulophidae) as biological control agents against Liriomyza trifolii (Diptera: Agromyzidae) in Japan. Applied Entomology and Zoology, Tokyo. 2006;(41):73-82.
21.Chu YI, Chien CC, Ku SC. Parasite strategy, morphology and life history of Tamarixia radiata (Hymenoptera, Eulophidae). Chinese Jour Entomology. 1991;11:264-281.
22.Neubert M, Castell H. Demography and dispersal: Calculation and sensivity analysis of invasion speed for estructured populations. Ecology. 2000;8(16):1613-1628.
23.Gómez TML, Nava ED, Parra PJR. Life Table of Tamarixia radiata (Hymenoptera: Eulophidae) on Diaphorina citri (Hemiptera: Psyllidae) at Different Temperatures. Journal of Economic Entomology. 2012;105(2):338-334.
24.Acosta NM, O´Neil RJ. Life history characteristics of three populations of Edovum puttleri Grissell (Hymenoptera: Eulophidae) at three temperatures. Biological Control, Orlando. 1999;16:81-87.
25.Castillo J, Jacas JA, Peña JE, Ulmer BJ, Hall DG. Effect of temperature on life history of Quadrastichus haitiensis (Hymenoptera: Eulophidae), an endoparasitoid of Diaprepes abbreviatus (Coleoptera: Curculionidae). Biological Control, Orlando. 2006;36:189-196.
26.Viggiani G, Bernardo U, Sasso R. Primi risultati sull'introduzione di Thripobius semiluteus Boucek (Hymenoptera: Eulophidae) in Italia per il controllo biologico di Heliothrips haemorrhoidalis (Bouché) (Thysanoptera). Atti Giornate Fitopatologiche, Perugia. 2000;16:521-526.
27.Castro Chacón Yerlin, López Silvia N. Biología de Eretmocerus mundus (Hymenoptera: Aphelinidae), parasitoide del complejo Hemisia tabaci (Homoptera: Aleyrodidae), en condiciones de laboratorio. Rev Soc Entomol Argent. 2010;69(1-2):45-56.
Recibido: 22-3-2013.
Aceptado: 28-5-2013.


{kind=link}
{kind=link}
{kind=link}
{kind=link}