










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista de Protección Vegetal
versión On-line ISSN 2224-4697
Rev. Protección Veg. vol.30 no.3 La Habana sep.-dic. 2015
ARTÍCULO ORIGINAL
Alternativa para el manejo de Phytophthora infestans (Mont.) de Bary en Solanum betaceum Cav. mediante inductores de resistencia
Management alternative for Phytophthora infestans (Mont.) de Bary in Solanum betaceum Cav. by resistance inducers
Julián Castaño MonsalveI, Joaquín Guillermo Ramírez GilI, Luis Fernando Patiño HoyosII, Juan Gonzalo Morales OsorioI*
IUniversidad Nacional de Colombia sede Medellín, Facultad de Ciencias Agrarias, Departamento de Ciencias Agronómicas. Bloque 11, Calle 59A, No 63-20, Medellín, Colombia, Núcleo El Volador. Tel: 0057 4 4309099, Fax: 0057 4 4309118.
IIPolitécnico Colombiano Jaime Isaza Cadavid-Facultad de Ciencias Agrarias, Carrera 48 No 7-151, Medellín, Colombia, tel: 0057 4 3197900.
RESUMEN
El tomate de árbol (Solanum betaceum Cav) es una fruta andina con amplias perspectivas de desarrollo comercial e industrial debido al potencial de consumo que tiene, tanto en fresco como en fruta procesada. Para buscar la sostenibilidad del cultivo se deben enfrentar retos tecnológicos, entre los que sobresale el manejo de plagas, el cual depende, casi exclusivamente, del uso de fungicidas e insecticidas para su control. En las dos últimas décadas apareció, y se ha incrementado notablemente, la enfermedad conocida como gota o peste negra, la cual ha ocasionado grandes pérdidas económicas en muchas zonas productoras. El agente causal se ha identificado como Phytophthora infestans sensu lato. Dada la importancia de esta enfermedad y la adaptabilidad del patógeno, este estudio tuvo como objetivo evaluar siete inductores de resistencia para el manejo de la enfermedad, aplicados tanto en aspersión al suelo como en la vía foliar y también se evaluó el posible efecto fitotóxico de estas moléculas. Los inductores no mostraron algún efecto significativo cuando se aplicaron en aspersión al suelo. Mediante aplicación foliar, el fosfito de potasio (Fos K, 0,75 cm3.l-1), quitosán (3 mg.ml-1), peróxido de hidrógeno (H202, 1,0 mM) y salicilato de sodio (SaNa 0,4 g.l-1) no mostraron reducción significativa del tamaño de la lesión ni del área bajo la curva del progreso de la enfermedad (AUDPC); mientras que el ácido b-aminobutírico (BABA, 1 g.l-1), metil jasmonato (MeJa 0,2 g.l-1) y acibenzolar-S-metil (ASM, 0,3 ml.l-1) lograron una reducción de 7,7; 5 y 4,28 veces en el diámetro de la lesión y de 18,5; 4,4 y 3,57 en el valor de la AUDPC, respectivamente.
Palabras clave: Phytophthora infestans sensu lato, resistencia inducida, tomate de árbol.
ABSTRACT
The tree tomato (Solanum betaceum Cav.) is an Andean fruit with wide perspectives for its commercial and industrial development. This fruit is consumed as both a fresh orprocessed product To search for crop sustainability, technological challenges must be confronted, among which pest management, which currently depends almost exclusively on fungicide and insecticide applications, stands out. Within the last two decades, the disease known as late blight or black pest appeared and notably spread causing large economic losses in several growing areas. The causal agent was identified as Phytophthora infestans sensu lato. Due to the disease importance and the high pathogen adaptability, the objective of the present study was to evaluate seven resistance inducers applied by spraying the soil or the foliage for this disease management. The potential phytotoxicity of these molecules was tested as well. None of the defense activators showed significant response when were applied to the soil. While foliar applications of potassium phosphite (0,75 ml.l-1), sodium salicylate (0,4 g.l-1), chitosan (3 mg.ml-1) and hydrogen peroxide (1,0 mM) had no significant effect on lesion diameter or the area under the disease progress curve (AUDPC), BABA (1g.l-1), methyl jasmonate (0,2 g.l-1 ), and ASM (0,3 ml.l-1) significantly reduced 7.7, 5 and 4.28 times the lesion diameter and 18, 5, 4,4 and 3.57 the AUDPC values, respectively.
Key words: Phytophthora infestans sensu lato, induced resistance, tree tomato.
INTRODUCCIÓN
El tomate de árbol (Solanum betaceum Cav.) es una fruta andina ampliamente cultivada en Colombia en la región alto andina, en zonas de relieve montañoso comprendidas entre 1500 y 3000 msnm. Esta especie presenta un gran potencial para incluirla en sistemas de cultivo con fines de exportación, debido a la gran aceptación de frutas exóticas en los mercados internacionales (1). Este cultivo presenta varios factores limitantes en su producción, principalmente las enfermedades. En los últimos años se informó la presencia de la enfermedad que se conoce como gota o peste negra, que produjo epidemias devastadoras en muchas zonas productoras.
El agente causal se identificó como Phytophthora infestans (Mont.) de Bary y recientemente se propuso una nueva especie, la cual se encontró asociada a tomate de árbol y que se denomina Phytophthora andina Adler & Flier (2). Hoy persiste la discusión sobre la aceptación de P. andina como nueva especie, por lo que la población del patógeno que ocasiona la gota en diversas especies de la familia Solanaceae se ha nombrado Phytophthora infestans sensu lato (3).
Esta enfermedad se controla casi exclusivamente con fungicidas de origen químico, ya que no existen variedades comerciales con resistencia. Los productos inductores de resistencia de las plantas a las enfermedades pueden constituir una importante alternativa para el manejo de diversas enfermedades en los cultivos agrícolas (4). La eficacia de estos inductores para el control de una enfermedad depende de numerosos factores, como son el genotipo del hospedante y del patógeno, la fitotoxicidad, la dosis y la frecuencia de aplicación, entre otros (4).
Con relación al uso de inductores de resistencia en el manejo de enfermedades causadas por oomicetos, se conoce que los Hidroxilados de quitosán resultaron ser elicitores potentes para inducir resistencia en tubérculos de papa hacia P. infestans; esto se evidencia por el menor número de células muertas en el tejido del tubérculo. La resistencia también se indujo por chitosán deaminado (5). Por otra parte, la aplicación de Acibenzolar-S-Metil (ASM) causó supresión del tizón ocasionado por Phytophthora capsici Leonian, en pimentón bajo condiciones de invernadero (6). Para el caso del ácido b aminobutírico (BABA), un aminoácido no proteico, se evidenció que puede inducir resistencia en papa al patógeno P. infestans (7).
La aplicación exógena de las hormonas ácido jasmónico (AJ) y ácido jasmónico metil-ester (MeJa) demostraron capacidad para inducir resistencia a enfermedades, pues retardan el progreso de P. infestans en papa (8). Con respecto al Peróxido de Hidrógeno (H2O2), se considera que puede estar involucrada en las respuestas de defensa. La aplicación del producto comercial Oxycom ®, el cual contiene entre 20-22% de H2O2, mostró una reducción de hasta 12% en el daño causado por el mildeo velloso en lechuga (9). Otro inductor, del que se ha informado ampliamente es el Fosfito de Potasio (Fos K), donde las aplicaciones de este lograron restringir el desarrollo de la enfermedad producida por P. infestans en papa y reducir la biomasa del patógeno en hojas (10). Así mismo, Kromann et al. (11), al evaluar diferentes productos basados en fosfonatos encontrados en los mercados de zonas de Perú y Ecuador productoras de papa, observaron que estos productos, aplicados a una dosis de 2,5 g.l-1, proporcionaron una eficacia similar a la ofrecida por fungicidas convencionales de contacto y propusieron la inducción de defensa dentro de los mecanismos para explicar el efecto de estos productos.
Dado el aumento de la incidencia y la severidad de P. infestans sensu lato en tomate de árbol, así como la alta dependencia del uso de fungicidas se planteó como objetivo analizar el efecto de siete inductores de resistencia sobre esta enfermedad para buscar métodos alternativos de control mediante la activación de los mecanismos de defensa en las plantas.
MATERIALES Y MÉTODOS
Ubicación
Este ensayo se realizó en la estación experimental Paysandú de la Universidad Nacional de Colombia, sede Medellín, ubicada en el corregimiento de Santa Elena, municipio Medellín, departamento de Antioquia, Colombia, a una altura de 2583 msnm, 2600 mm de precipitación promedio anual, temperatura de 14ºC; se clasifica como zona de vida, bosque muy húmedo montano bajo (bmh-MB) (12).
Material Vegetal
Se seleccionaron frutos sanos de tomate de árbol, de la variedad comercial rojo común que se siembra en Colombia. Se realizó la extracción de las semillas de los frutos maduros en escala 6 (13) y se dejaron fermentar durante 3 días; luego se lavaron con abundante agua y se pusieron a secar a la sombra. Las semillas se sembraron en bandejas de 128 cavidades en sustrato turba. Tres semanas después de la emergencia, las plántulas se trasplantaron a macetas de 1 kg de capacidad, con sustrato comercial compuesto de suelo, cascarilla de arroz, gallinaza y equinaza (Delpino™ Ltda., Medellín-Colombia) bajo condiciones de casa-malla, con temperatura promedio de 14°C y humedad relativa promedio de 80%. Las plantas se fertilizaron con 5g de fertilizante compuesto NPK (15:15:15) 20 días después del trasplante y fertilización foliar completa semanal (Complesal, Bayer CropScience™), siguiendo las recomendaciones del fabricante. Se aplicó riego para mantener las plantas a capacidad de campo.
Aislamiento e identificación de P. infestans sensu lato
El inóculo se preparó a partir de un aislamiento colectado de hojas de tomate de árbol (S. betaceum) que mostraron síntomas típicos ocasionados por P. infestans sensu lato (código Solanum betaceum 11). Los foliolos con síntomas se llevaron al laboratorio donde se lavaron con abundante agua, se secaron al aire y se seccionaron en trozos pequeños (~1 cm2). Luego se introdujeron en cabina de flujo laminar, donde se desinfectaron con etanol (70%) por 30 s, seguido de un lavado en agua destilada estéril por 30 s, luego en hipoclorito de sodio al 3% por 30 s y, por último, se realizó un lavado con agua destilada estéril por 30 s. Después de la desinfección, se cortaron trozos pequeños (3 mm2) y se sembraron en medio de cultivo agar-centeno (Difco, USA) y se incubaron durante 14 días a 18ºC en oscuridad. Las estructuras morfológicas del oomyceto se observaron en microscopio de luz (Nikon Eclipse Ni 200).
Un fragmento de cultivo (~5 mm de diámetro), tomado del borde de crecimiento, se transfirió a medio líquido de arveja (120 g.ml-1) y se incubó bajo las mismas condiciones descritas. El micelio se recuperó por filtración usando papel Whatman No 1 y, posteriormente, se liofilizó hasta su uso. Para extraer el DNA se siguió la metodología reportada por Goodwin et al. (14). El DNA purificado se resuspendió en 50 µl de Tris 10mM pH 8.5. Una vez que se extrajo el ADN de los aislamientos, se realizó la amplificación de las secuencias del intrón 1 del gen nuclear de copia única ras, en un termociclador Labnet Multigene Optimax (Labnet International Inc., USA), según la metodología informada por Gómez-Alpizar et al. (15). Los productos se enviaron a secuenciar a la empresa Macrogen (Korea). Las secuencias obtenidas se editaron mediante el programa BioEdit 6.0.6 y Clustal W (16) y se confirmó su identidad por comparación con las bases de datos moleculares, mediante el programa BLAST, disponible en la página web: http://blast.ncbi.nlm.nih.gov/Blast.cgi?CMD=Web& PAGE_TYPE=BlastHome (17). La secuencia del gen se usó junto con el análisis morfológico para identificar el aislamiento.
Inoculación
Para obtener el inóculo se aplicó una solución de esporangios del aislamiento identificado sobre foliolos frescos de tomate de árbol, los cuales se incubaron en cámara húmeda (18ºC, 90-100% de humedad relativa) hasta obtener abundante esporulación (7-10 días). Los esporangios se colectaron a partir de tejidos vegetales infectados mediante su lavado con agua destilada estéril y se cuantificaron en cámara de Neubauer. La inoculación del patógeno se realizó 24 horas después de la aplicación de los productos inductores utilizando una concentración de 3000-5000 esporangios ml.l-1, la cual asegura una infección homogénea (18). Esta se realizó aplicando 6 gotas/hoja, 3 a cada lado de la nervadura central en el envés de las 4 primeras hojas apicales que se encontraban totalmente expandidas. Cada gota tenía un volumen de 25 µl. Para garantizar que la gota conservara su forma y evitar la pérdida del inóculo, este se preparó en una solución de agua suplementada con agar al 0,1%. Después de la inoculación, las plantas se mantuvieron en condiciones de casa-malla bajo las condiciones descritas previamente. Para detectar los síntomas de gota, las plantas se examinaron cada dos días después de la inoculación por un periodo de 10 días.
Ensayos de fitotoxicidad
Se utilizaron siete inductores de resistencia de las plantas: fosfito de potasio (Agrifos®), acibenzolar-S-metil (ASM) (Boost®), quitosano de bajo peso molecular (Sigma-Aldrich® 448869), metil jasmonato (MeJa) (Sigma-Aldrich® 392707 95%), ácido b-aminobutírico (BABA) (Sigma-Aldrich A44207 97%), peróxido de hidrógeno al 50% (H2O2) (JM Químicos S.A., Medellín-Colombia) y salicilato de sodio (SaNa) (Alfa Aesar A17056 99%). Los compuestos y las dosis correspondientes que se analizaron en este trabajo se pueden visualizar en la Tabla 1.
Para la preparación del quitosano se tomaron cada una de las dosis y se disolvieron sobre la base de 0,075 g en 10 ml de ácido acético al 1% y se agregó Tween® 80 al 0,1%. Luego se llevó al agitador durante 12 horas a temperatura ambiente y a 200 rpm. El volumen final de 250 ml se completó con agua destilada. Así mismo, para la preparación del metil jasmonato, se dispuso de una solución madre con una concentración de 0,5 g 10 ml-1, con el fin de obtener la concentración requerida para cada dosis; la molécula grado Sigma-Aldrich se disolvió en 25 ml de etanol al 10% y se agregó TRITON X-100 al 0,1%. El volumen final de 250 ml se completó con agua destilada.
Tanto las pruebas de fitotoxicidad como las de inducción de resistencia se realizaron sobre plantas de 14 semanas de edad después del trasplante, que crecieron según se describió en material vegetal. Para cada inductor, las tres dosis se aplicaron tanto en aspersión al suelo como en aspersión foliar. Cada tratamiento se realizó sobre cinco plantas. En el suelo se aplicaron 25 ml por planta; para la aspersión foliar se aplicó un volumen aproximado de 25 ml hasta cubrir completamente el área foliar. Después de realizados los tratamientos, las plantas se mantuvieron en condiciones de casa-malla a 22-25 °C día y 10-16°C noche y 80% de humedad relativa en promedio, durante el tiempo de evaluación del experimento.
Ensayo de inducción de resistencia
Las plantas y la inoculación se realizaron como se describió previamente. Las dosis de los inductores de resistencia se seleccionaron a partir del experimento anterior y se usaron aquellas que no mostraron daño sobre los tejidos vegetales.
Variables evaluadas
Se determinó, como variables de respuesta, el diámetro de la lesión y el área bajo la curva del progreso de la enfermedad AUDPC (Area Under Disease Progressive Curve) con el fin de evaluar el desarrollo de la lesión para cada uno de los tratamientos, durante un periodo de diez días después de la inoculación.
Diseño experimental y análisis de los datos
Tanto el experimento de fitotoxicidad como el de inducción de resistencia se desarrollaron en un diseño completamente al azar con 5 repeticiones por tratamiento. A los datos se les realizó análisis de varianza (ANOVA) y posterior comparación de medias a través de la prueba de LSD, con una confiabilidad del 95%; para ello se utilizó el programa STATGRAPHICS Centurion XVI (Statpoint Technologies Inc.). Los experimentos se realizaron dos veces en el tiempo.
RESULTADOS Y DISCUSIÓN
Identificación del patógeno utilizado
Sobre la base de las observaciones morfológicas y de la secuencia del intrón 1 del gen nuclear de copia única ras, el aislamiento utilizado en este trabajo correspondió a Phytophthora infestans sensu lato. Los tres valores mayores de identidad estuvieron por encima del 93% y correspondieron a aislamientos de P. andina colectados en Ecuador: P13642 (93% de identidad, 5e-106, colectado en S. betaceum), P13576 (93% de identidad, 5e-106, colectado en Solanum sp. Anarrhichomenum complex) y P13655 (94% de identidad, 6e-105, colectado en Solanum hispidium) (17,19). Actualmente, la comunidad científica se encuentra en una discusión muy intensa acerca de si P.andina y P. infestans son la misma especie o si son diferentes; por esta razón se denomina el grupo de genotipos relacionados como P. infestans sensu lato (3).
Prueba de fitotoxicidad
En la prueba de fitotoxicidad se encontró que las plantas que se trataron con Fos K en las dosis de 5 y 7 ml.l-1, MeJa en 0,3 g.l-1 y ASM en 0,4 ml.l-1 presentaron lesiones producidas por los respectivos productos. Se confirmó que el origen del daño en los tejidos provenía de la aplicación de estos productos, debido a que se realizó la evaluación en el tiempo a las muestras y no se encontraron estructuras de microorganismos asociados cuando se realizó la observación en el microscopio de luz. Los resultados de toxicidad coinciden con lo informado en distintos trabajos, donde el Fos K puede llegar a causar fitotoxicidad en varios cultivos cuando se aplica en altas concentraciones (20). Por otra parte, bajo las aplicaciones de ASM, también se observaron síntomas fitotóxicos en tomate de árbol (S. betaceum Cav.) que se manifiestan en hojas, como son la deformación, el encrespamiento, las ampollas, el amarillamiento y la deformación de los nuevos brotes (21). Sobre la base de los resultados de las pruebas de toxicidad, se seleccionaron las dosis con mayor potencial de control, pero que no afectaron los tejidos de las plantas.
Dosis de los inductores evaluados para el manejo de la gota
De acuerdo con la prueba de fitotoxicidad, se definieron las siguientes dosis: fosfito de potasio (0.75 ml.l-1), ASM (0,3 ml.l-1), quitosano (3,0 mg.ml-1), MeJa (0,2 g.l-1), BABA (1,0 g.L-1), H2O2 (1,5 mM) y SaNa (0,4 g.l-1). En los otros productos no se encontró algún efecto tóxico sobre las plantas y se usó la mayor dosis evaluada para buscar mayor efecto sobre la inducción de resistencia.
Ensayo de inducción de resistencia sobre el control de la gota
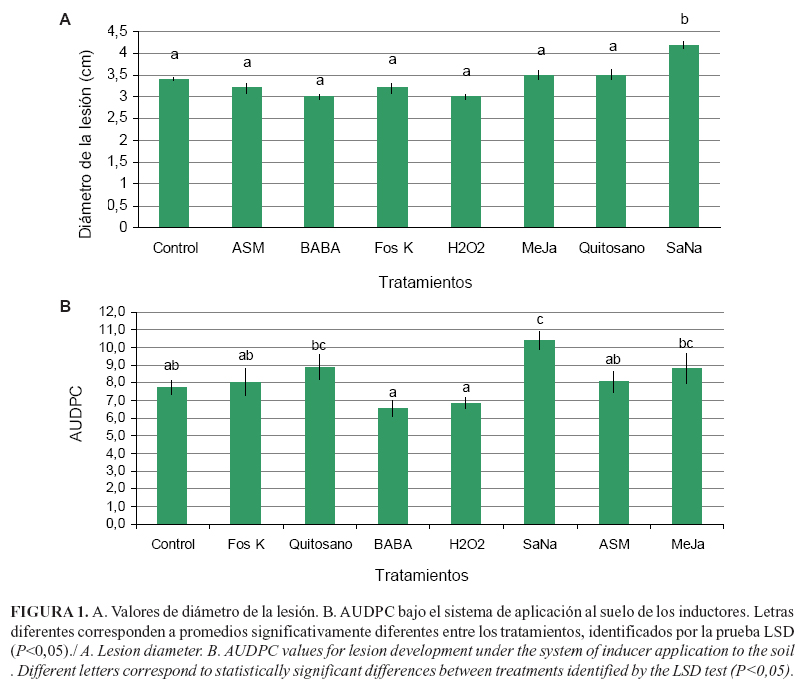
Cuando se realizó la aplicación de los inductores de resistencia en aspersión al suelo y posterior inoculación con el patógeno P. infestans, no se observó efecto significativo (P>0.05) sobre la disminución del diámetro de la lesión (Fig. 1 A) e, igualmente, sobre el grado de desarrollo de la enfermedad que se midió mediante la AUDPC de algún inductor con respecto al tratamiento control (Fig. 1 B). Por el contrario, se observó un valor superior cuando las plantas se trataron con SaNa. Esta inducción de mayor susceptibilidad puede ocurrir para determinados activadores de resistencia, donde el estilo de vida del patógeno y la vía de señalización de las defensas activadas son los factores más influyentes (22).
En el sistema de aplicación foliar
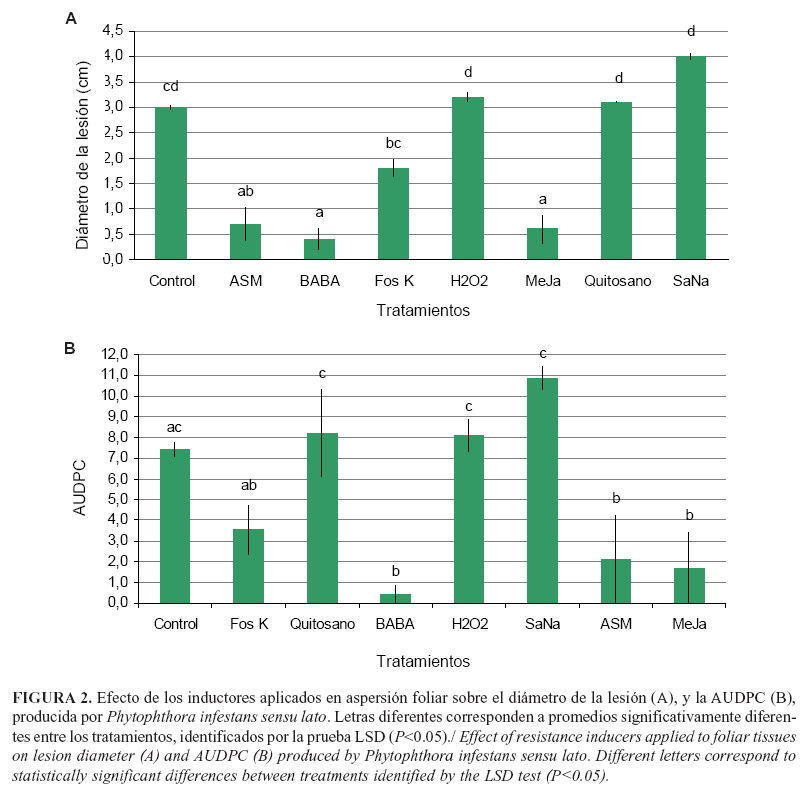
La aplicación foliar de BABA, MeJa y ASM logró disminuir (P<0,05) el tamaño de la lesión en 7,7; 5 y 4,28 veces con respecto al tratamiento control (Fig. 2 A). La comparación entre estos no mostró diferencias estadísticas (P>0,05), lo que indica que lograron el mismo nivel de control de la gota en tomate de árbol. En cuanto al diámetro de las lesiones, los tratamientos con Fos K, H2O2, quitosano y SaNa no mostraron diferencias significativas (P>0,05) en comparación con el tratamiento control (Fig. 2 A). Lo encontrado para la variable anterior se confirma por los valores hallados para AUDPC, donde se halló que el BABA, MeJa y ASM reducen el desarrollo de la enfermedad significativamente (P<0,05): 18,5; 4,4 y 3,57 veces con respecto al control (Fig. 2 B).
Estos resultados muestran evidencia que permite inferir que el modo de aplicación de los inductores podría ser importante para el control de la gota en tomate de árbol, donde las aplicaciones foliares son más efectivas que aquellas realizadas en aspersión al suelo, lo cual coincide con lo informado por Liljeroth et al. (7), quienes encontraron una reducción cerca del 50% del tamaño de la lesión de tizón en papa mediante aspersión foliar de BABA, en comparación con una reducción del 10-20% con aplicaciones de este inductor en aspersión al suelo. Contrariamente, también se ha reportado para BABA y ASM que estos, al aplicarse al suelo, presentaron control significativo de enfermedades producidas por Peronospora tabacina y Phytophthora sp. (23, 24). El hecho de no haber encontrado efecto de los tratamientos aplicados en aspersión al suelo sugiere que es posible que los productos hayan sufrido algún proceso de inactivación o disminución de la absorción por raíces, debido a factores físicos, químicos o biológicos presentes en el suelo.
Los niveles altos de reducción de la enfermedad, por efecto de la resistencia inducida por BABA, coinciden con numerosos informes de la acción de esta molécula sobre el desarrollo de enfermedades causadas por Phytophthora spp. (7), en los que evidenciaron reducciones hasta del 40-50% en el patosistema P. infestan en papa, comparado con el control no tratado. Se ha propuesto que el amplio espectro del BABA se basa en activación simultánea de mecanismos de defensa, dependientes e independientes del ácido salicílico (AS), donde para los primeros se requiere una proteína funcional NPR1, un gen receptor kinasa inducible por BABA y la producción de ácido pipecolico (25). Por otro lado, el componente de BABA, independiente de AS, se encuentra asociado con la deposición de callosa y requiere biosíntesis y percepción intacta de ácido abscísico (ABA) (26).
Con relación al MeJa, se informó que su efecto podría deberse a la rápida inducción de H2O2 durante las etapas tempranas del proceso de infección (27). Al respecto, Tejeda et al. (8) indicaron que las plantas de papa tratadas con ácido jasmónico (AJ) fueron claramente protegidas contra la infección de P. infestans. Maksimov et al. (28) informaron una significativa inducción de mecanismos de defensa en papa inoculada con P. infestans, cuando se trataron con ácido salicílico o ácido jasmónico y se identificó un incremento aún mayor cuando se usó una mezcla de ambas hormonas. Después de los tratamientos se registró incremento de compuestos fenólicos y la activación de la enzima peroxidasa en los lugares de ubicación del patógeno. En el mismo trabajo también se encontró que cuando se aplicó ácido jasmónico se activó la síntesis de H2O2; sin embargo, todo lo contrario sucedió cuando solo se aplicó ácido salicílico, pues no se identificó inducción de síntesis de H2O2. Por su parte, el uso del ASM ha sido ampliamente reportado en el manejo de enfermedades en las plantas (6, 21,24), lo cual coincide con lo encontrado en este trabajo, donde este compuesto logró disminuir los efectos negativos de la gota en tomate de árbol. Se encontró que el ASM actúa como análogo del AS en la vía de señalización de SAR e incrementa la expresión de genes relacionados con la defensa y que participan en vías metabólicas que conducen a la inducción de síntesis de lignina y compuestos fenólicos (29).
En este trabajo no se identificó un efecto positivo de la aplicación de los fosfitos de potasio, producto ampliamente utilizado en la agricultura en el manejo de enfermedades causadas por oomicetes con resultados satisfactorios (10, 11). Estos hallazgos indican la necesidad de estudiar muchos aspectos asociados a la utilización de los inductores de resistencia, con el fin de lograr obtener los resultados adecuados y así poder tener una opción más para el manejo de las enfermedades de las plantas. Dentro se estos aspectos es necesario estudiar los mecanismos específicos de acción que se presentan en S. betaceum para entender cómo la aplicación de estos inductores protege contra la gota.
Los trabajos que involucran los inductores de resistencia son casi inexistentes en el cultivo de tomate de árbol. Los resultados encontrados en este trabajo se perfilan como un punto de referencia para el control de la enfermedad, conocida como gota, producida por P. infestans sensu lato en el cultivo de tomate de árbol, mediante el uso de inductores de resistencia, donde el ASM, el BABA y el MeJa se proyectan como un componente importante en el futuro manejo integrado de esta enfermedad.
CONCLUSIÓN
Los productos BABA, metil jasmonato y ASM, cuando se aplicaron en forma de aspersión foliar, provocaron reducción significativa de los niveles de la enfermedad denominada gota del tomate de árbol.
AGRADECIMIENTOS
Este trabajo fue financiado por la Universidad Nacional de Colombia, sede Medellín, mediante el proyecto titulado: «Alternativas de manejo de la gota del tomate de árbol (Solanum betaceum Cav), mediante el uso de inductores de resistencia y caracterización del organismo causal (Phytophthora infestans sensu lato)», «Convocatoria para cofinanciar proyectos nuevos de investigación, desarrollo tecnológico e innovación vigentes a profesores de la Facultad de Ciencias Agrarias, Sede Medellín», RESOLUCIÓN A-0891 DE 2012 Mayo 18.
REFERENCIAS
1. Lagos T, Benavides C, Paredes R, Lagos L. Distribución del tomate de árbol Cyphomandra betacea (Cav.) Sendt. y caracterización eco-climática en las zonas del cultivo de Nariño. Rev Colomb Cienc Hortic. 2011;5(1):11-19.
2. Oliva R, Kroon L, Chacón G, Flier W, Ristaino J, Forbes G. Phytophthora andina sp. nov., a newly identified heterothallic pathogen of solanaceous hosts in the Andean highlands. Plant Pathol. 2010;59:613-625.
3. Cárdenas M, Grajales A, Sierra R, Rojas A, González-Almario A, Vargas A, et al. Genetic Diversity of Phytophthora infestans in the Northern Andean Region. BMC Genet. 2011;12:23.
4. Walters D. Induced resistance: destined to remain on the sidelines of crop protection? Phytopar. 2010;38:1-4.
5. Vasyukova N, Ozeretskovskaya O, Chalenko G, Gerasimova N, L'vova A, Il'ina A, et al. Immunomodulating Activity of Chitosan Derivatives with Salicylic Acid and Its Fragments. Appl Biochem Microbiol. 2010;46(3):346-351.
6. Matheron M, Porchas M. Suppression of Phytophthora root and crown rot on pepper plant treated with acibenzolar-S-methyl. Plant Dis. 2002;86:292-297.
7. Liljeroth E, Bengtsson T, Wiik L, Andreasson E. Induced resistance in potato to Phytophthora infestans-effects of BABA in greenhouse and field tests with different potato varieties. Eur J Plant Pathol. 2010;127:171-183.
8. Tejeda M, Martínez N, Olalde V, Délano J. Jasmonic Acid accelerates the expression of a pathogen-specific lipoxygenase (POTLX-3) and delays foliar blight development in potato (Solanum tuberosum L.). Rev Mex Fitopatol. 2007;25(1):18-25.
9. KimY, Blee K, Robins J, Anderson A. Oxycom™ under field and laboratory conditions increases resistance responses in plants. Eur J Plant Pathol. 2001;107:129-136.
10.Florencia M, Lobato M, Feldman M, Daleo G, Andreu A. Potassium phosphite primes defense responses in potato against Phytophthora infestans. J Plant Physiol. 2012;169:1417-1424.
11.Kromann P, Pérez W, Taipe A, Schulte-Geldermann E, Sharma B, Andrade-Piedra J, et al. Use of phosphonate to manage foliar potato late blight in developing countries. Plant Dis. 2012;96:1008-1015.
12.Holdridge L. Tropical Science Center. Life Zone Ecology. San José, Costa Rica. 1967.
13.ICONTEC- Instituto Colombiano de Normas Técnicas y Certificación. Norma técnica colombiana (NTC-4105). Bogotá, D.C. 1997.
14.Goodwin S, Spielman L, Matuszak J, Bergeron S, Fry W. Clonal diversity and genetic differentiation of Phytophthora infestans populations in Northern and Central Mexico. Phytopathology. 1992;82:955-961.
15.Gomez-Alpizar L, Hu C, Oliva R, Forbes G, Ristaino J. Phylogenetic relationships of Phytophthora andina, a new species from the highlands of Ecuador that is closely related to the Irish potato famine pathogen Phytophthora infestans. Mycol. 2008;100:590-602.
16.Hall T. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucl Acids Symp Ser. 1999;41:95-98.
17.Morgulis A, Coulouris G, Raytselis Y, Madden T, Agarwala R, Schäffer A. Database indexing for production MegaBLAST searches. Bioinfor. 2008;24:1757-1764.
18.Whisson S, Boevink P, Moleleki L, Avrova A, Morales J, Gilroy E, et al. A translocation signal for delivery of oomycete effector proteins into host plant cells. Nature. 2007;450(7166):115-118.
19.Blair J, Coffey M, Martin F. Species tree estimation for the late blight pathogen, Phytophthora infestans, and close relatives. PLoS ONE. 2012;7(5):e37003. doi:10.1371/journal.pone.0037003.
20.Bock C, Brenneman T, Hotchkiss M, Wood B. Evaluation of a phosphite fungicide to control pecan scab in the southeastern USA. Crop Prot. 2012;36:58-64.
21.Mejía D, Rodas E, Patiño L, González E. Efecto del acibenzolar-s-metil sobre el desarrollo de la virosis causada por potyvirus en tomate de árbol. Agron Col. 2009;27(1):87-93.
22.Walters D, Havis N, Sablou C, Walsh D. Possible trade-off associated with the use of a combination of resistance elicitors. Physiol Mol Plant Pathol. 2011;75:188-192.
23.Cohen Y. 3-Aminobutyric acid induces systemic resistance against Peronospora tabacina. Physiol Mol Plant Pathol. 1994;44(4):273-288.
24.Ji P, Yin J, Koné D. Application of acibenzolar-S-methyl and standard fungicides for control of Phytophthora blight on squash. Crop Prot. 2011;30:1601-1605.
25.Navarova H, Bernsdorff F, Doring A, Zeier J. Pipecolic acid, an endogenous mediator of defense amplification and priming, is a critical regulator of inducible plant immunity. Plant Cell. 2012;24:5123-5141.
26.Ton J, Mauch-Mani B. Beta-amino-butyric acid-induced resistance against necrotrophic pathogens is based on ABA-dependent priming for callose. Plant J. 2004;38:119-130.
27.Zhu Z, Tian S. Resistant responses of tomato fruit treated with exogenous methyl jasmonate to Botrytis cinerea infection. Sci Hortic. 2012;142:38-43.
28. Maksimov I, Sorokan' A, Chereoanova E, Surina O, Troshina N, Yarullina L. Effects of Salicylic and Jasmonic Acids on the Components of Pro/Antioxidant System in Potato Plants Infected with Late Blight. Russ J Plant Physiol. 2011;58(2):299-306.
29.Malolepsza U. Induction of disease resistance by acibenzolar-S-methyl and o-hydroxyethylorutin against Botrytis cinerea in tomato plants. Crop Prot. 2006;25:956-962.
Recibido: 12-3-2015.
Aceptado: 3-9-2015.
*Autor para correspondencia: Juan Gonzalo Morales Osorio. Correo electrónico: jgmoraleso@unal.edu.co.


{kind=link}
{kind=link}
{kind=link}