










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkVaccimonitor
versión On-line ISSN 1025-0298
Vaccimonitor v.19 n.1 Ciudad de la Habana ene.-abr. 2010
ARTICULOS ORIGINALES
Evaluation of the potential of Mycobacterium smegmatis as vaccine Candidate against tuberculosis by in silico and in vivo studies.
Evaluación del potencial de Mycobacterium smegmatis como candidato vacunal contra la tuberculosis mediante estudios in sílico e in vivo.
Le Thuy Nguyen Thi1, Reinier Borrero Maura2, Sonsire Férnandez2, Giselle Reyes2, José Luis Perez2, Fátima Reyes2, María de los Angeles García2, Midrey Fariñas2, Juan Francisco Infante2, Yanely Tirado2, Alina Puig2, Gustavo Sierra2, Nadine Álvarez2 , Juan Carlos Ramírez2, María Elena Sarmiento2, Mohd-Nor Norazmi3, Armando Acosta2*
1 Biotechnology Centre of Ho Chi Minh City, 176 Hai Ba Trung Street, Ho Chi Minh City, Vietnam.
2 Finlay Institute. Center for Vaccine Research & Production. Ave 27 No 19805, La Lisa, Havana City, AP16017 CP11600, Cuba.
3 School of Health Sciences, Universiti Sains Malaysia, 16150 Kubang Kerian, Malaysia
* Dr. en Ciencias Médicas y Médico Especialista de 2do. Grado en Inmunología, Investigador Titular y Profesor Titular. Director del Dpto.de Biología Molecular de la Vicepresidencia de Investigaciones del Instituto Finlay.
email: aracosta@finlay.edu.cu
ABSTRACT
In this study, we scanned multiple published databases of gene expression in vivo of M. tuberculosis at different phases of infection in animals and humans, to select 38 proteins that are highly expressed in the active, latent and reactivation phases. The selected proteins were predicted for T and B epitopes. For each proteins, the regions containing T and B epitopes were selected at the same time to look for identical epitopes on M. smegmatis based on sequence alignments. Preliminary studies of humoral immunogenicity and cross-reactivity with M. tuberculosis in mice using two M. smegmatis-derived experimental vaccines were carried out, demonstrating the immunogenicity of M. smegmatis proteoliposomes and the recognition of M. tuberculosis proteins by the sera of animals immunized with this vaccine candidate. The conjunction of in silico and in vivo studies suggested the potential for future evaluation of M. smegmatis as vaccine candidate against tuberculosis using different strategies.
Keywords: Mycobacterium tuberculosis, Mycobacterium smegmatis, epitopes, prediction.
RESUMEN
En este estudio se revisaron múltiples bases de datos publicadas, relacionadas con experimentos de expresión de genes de M. tuberculosis in vivo en diferentes estadios de la infeccción en humanos y animales. Se identificaron 38 proteínas con elevada expresión en las fases activa, latente y de reactivación de la infección. Se llevó a cabo la predicción de epítopes T y B en dichas proteínas. Las regiones de cada proteína que contenían simultáneamente epítopes T y B se seleccionaron y utilizaron para identificar regiones idénticas en M. smegmatis mediante el alineamiento de secuencias. Se llevaron a cabo estudios de inmunogenicidad humoral y reactividad cruzada con M. tuberculosis en ratones inmunizados con dos vacunas experimentales obtenidas a partir de M. smegmatis, demostrándose la immunogenicidad de los proteoliposomas y el reconocimiento de proteínas de M. tuberculosis por el suero de ratones vacunados con este candidato vacunal. Los resultados obtenidos con los estudios in sílico e in vivo sugieren la potencialidad para evaluación futura de candidatos vacunales obtenidos a partir de M. smegmatis para la prevención de la tuberculosis.
Palabras claves: Mycobacterium tuberculosis, Mycobacterium smegmatis, epítopes, predicción.
INTRODUCTION
Mycobacterium tuberculosis, the main agent of tuberculosis (TB) in humans caused 9.2 million new cases and 1.7 million deaths in 2006. It is estimated that up to one third of the world's population has been infected with M. tuberculosis (1). The current vaccine, Mycobacterium bovis Bacillus Calmette-Guerin (BCG) is the most widely used in humans. However, the uncertain efficacy of BCG vaccine in the prevention of pulmonary TB in adults, and the emergence of extensively drug resistant M. tuberculosis strains have further enhanced the sense of urgency in the development of new TB vaccines (2). With the complete genome sequence of M. tuberculosis and the advances in the sequencing of BCG and other mycobacteria, an enormous flow of valuable information has been made available for new vaccine strategies (2, 3).
Vaccine design is highly suited to the application of in silico techniques, for both discovery and development of new and existing vaccines (3). Bioinformatic tools allow selecting regions of microbial genomes that trigger immune responses against vaccine components. In order to give a bigger impact to the bioinformatic studies it is important to take into consideration the data obtained from studies of gene expression in vivo of M. tuberculosis. This combination will focus the studies on a smaller group of relevant antigens to be used in vaccine development (3).
The fast-growing and nonpathogenic soil bacterium Mycobacterium smegmatis which shares many features and identical genomic sequences with the pathogenic M. tuberculosis (4) is a potential candidate for developing new TB vaccines. The use of M. smegmatis is expected to have more advantages for TB vaccine development, feasible for research in normal laboratories, saving time and cost. With the aim to evaluate the antigenic similarity between M. smegmatis and M. tuberculosis, a combination of in silico and in vivo studies was carried out.
MATERIALS and METHODS
Identification and selection of in vivo expressed genes of M. tuberculosis
A search using the Internet Google Search Tool to find articles reporting in vivo studies of M. tuberculosis gene expression in humans and animals was carried out. The genes that are highly expressed in common in humans and animals at all stages of infection: activation, latency and reactivation were selected for epitope prediction.
Epitope prediction
The amino acid sequences and the features of the selected genes were obtained from the Reference Sequence (RefSeq) database (http://www.ncbi.nlm.nih.gov/RefSeq/) at the National Center for Biotechnology Information (NCBI) (http://www.ncbi.nlm.nih.gov/).
T-cell epitope prediction
HLAPred server was used for prediction of T-cell epitopes in this study. The program is developed by Bioinformatics Centre Institute of Microbial Technology, India (http://www.imtech.res.in/ raghava/hlapred/). All the alleles from the list of 36 HLA class I and 51 HLA class II were selected for the prediction.
B-cell epitope prediction
Two servers, Bcepred and ABCpred, were combined for linear B-cell epitope prediction. Bcepred server is accessible at http://www.imtech.res.in/raghava/bcepred/ and ABCpred web server is accessible at www.imtech.res.in/raghava/abcpred/.
For Bcepred server we combined of 7 physico-chemical properties of amino acids (hydrophilicity, flexibility, accessibility, polarity, exposed surface and turns and antigenic propensity). For ABCpred server we predicted B epitope length of 16 amino acids.
Searching for identical epitopes on M. smegmatis
The regions in the sequences of the 38 selected proteins of M. tuberculosis containing T and B epitopes at the same time were used to identify the identical epitopes on M. smegmatis genome using BLAST.
(http://www.ncbi.nlm.nih.gov//genomes/geblast.cgi?gi=20087)
Prediction of subcellular localization of proteins from M. tuberculosis and M. smegmatis
The subcellular localization of the selected M. tuberculosis proteins were defined according to the report of the identification and localization of 1044 M. tuberculosis proteins using two-dimensional, capillary high-performance liquid chromatography coupled with mass spectrometry (2DLC/MS) method (5).
The amino acid sequences of M. smegmatis proteins containing identical epitopes with M. tuberculosis and their features were obtained from the Reference Sequence (RefSeq) database (http://www.ncbi.nlm.nih.gov/RefSeq) at NCBI. To determine the subcellular location of the proteins of M. smegmatis and the proteins of M. tuberculosis that were not in the first list of 1044 proteins, we used TBpred server. TBpred is freely available at http://www.imtech.res.in/raghava/tbpred/
Evaluation of the immunogenicity and the cross-reactivity with M. tuberculosis on mice immunized with M. smegmatis vaccine candidates
M. smegmatis culture
Lyophilized cells of M. smegmatis from Molecular Biology Laboratory, Finlay Institute, were used in this study. The cells were grown at 37 °C, shaked at 200 rpm in culture broth medium containing glycerol 0.05 % and 0.05 % tween 80.
Preparing proteoliposomes from M. smegmatis
Biomass of M. smegmatis was obtained from centrifugation of the broth culture at 17700 x g (centrifuge J2-21, Beckman, EUA) during 1 hour, at 4 oC. The biomass was suspended in buffer (10-30 mM Tris, 2-10mM EDTA, pH 7-8.5) and washed twice. After suspension of the pellet in the same buffer, sodium deoxycholate (5-15%) was added with the ratio 0.1-0.25 mL/g biomass. After centrifugation with the same conditions, the supernatant was ultra-centrifugated at 50000-70000 x g (centrifuge L7-65 Beckman, EUA) for 2-8 hours at 4 °C . The sediment was resuspended in the buffer and filtrated with Sartorius Minisart-plus filter 0.45-0,2 µm.
Mice immunization
Fifteen female Balb/c mice of 6-8 weeks of age (16-18 g weight) obtained from the National Center of Laboratory Animals (CENPALAB), La Habana, Cuba, were used. The animals were housed in appropriate biological containment facilities at Animal Laboratory, Finlay Institute. All the experimental procedures were carried out following the guide for the use of Laboratory Animals (EEC Council Directive, 1987). The animals were distributed into 3 experimental groups (n=5): (i) negative control group consisted of mice receiving one dose at T0 of PBS; (ii) mice receiving two doses of proteoliposomes of M. smegmatis (50 µg/dose/mouse) in Freund's incomplete adjuvant at T0 and T21 (day 21 after the first immunization); (iii) mice immunized by a single dose at T0 with live cells of M. smegmatis (1 x 106 cfu).
Mice of all groups were injected via the subcutaneous route. 42 days after the first inoculum, the blood of mice of all groups was collected and centrifugated to obtain the sera.
ELISA to determine antibody responses
Flat bottomed polystyrene ELISA plates (MaxiSorp, Nunc) were coated (100 µL/well) 16 hours at 4 °C with 5 µg/mL M. smegmatis proteoliposomes, 5 µg/mL whole cell lysate of M. tuberculosis. 106 cfu/mL of live-cell M. smegmatis were coated 16 hours at 37 °C. The antigens were dissolved in the coating buffer (Carbonate/Bicarbonate 0,1 M, pH 9,6) and added to the plates. The plates were blocked with blocking buffer (PBS with 3% skim milk) for 1 hour at 37 oC. Serum samples were diluted 1/50 with 1% skim milk (w/v) in PBS (NaCl 137 mM, KCl 2.7 mM, Na2HPO4.7H2O, KH2PO4 1.4 mM, pH 7.4 and were added (100 µL/well) into each well. The samples were assayed in duplicate and were incubated 1 hour at 37 °C . The plates were incubated for 1 hour at 37 °C with the peroxidase-conjugated secondary antibody (affinity-purified anti-mouse IgG; Sigma, cod A-4416) in a dilution of 1/1000 (100 µL/well). The reaction was developed with the solution of o-phenylenediamine (OPD; Sigma): 5 mg of OPD in 12.5 mL of sodium citrate buffer 0.1M, pH 5, H2O2 5 µL (100 ìL/well). The reaction was stopped with 50 µL/well of H2SO4 2N after 20-30 minutes incubation with OPD solution. The absorbance was recorded at 492 nm with an ELISA plate reader (Multiskan Multisoft). Each experiment was carried out twice.
Statistical analysis
ELISA results were analyzed using ANOVA to determine significant differences among the groups at the 95% confidence level. Multiple range test were used to determine pairs with significant differences.
RESULTS
Identification and selection of in-vivo highly expressed genes of M. tuberculosis
In this study, we analyzed published articles of gene expression of M. tuberculosis during different phases of infection in animals and humans (6-24): from humans, animals (mainly on mice and rabbits) and from both humans and mice. These studies were directed to identify M. tuberculosis genes of responsible for the adaptation and evasion from the host immune responses and drugs as well as for the reactivation of infection and invasion of the bacteria. The expression profile under different conditions of 3924 genes from M. tuberculosis during different stages of infection in different hosts was identified (11, 12, 16-19).
A total of 38 M. tuberculosis genes that were reported significantly up-regulated at the active, latent and reactivation phases were selected from the whole genome. The amino acid sequences obtained and the features of the selected genes from the Reference Sequence (RefSeq) database the subcellular localization of the corresponding proteins were identified.
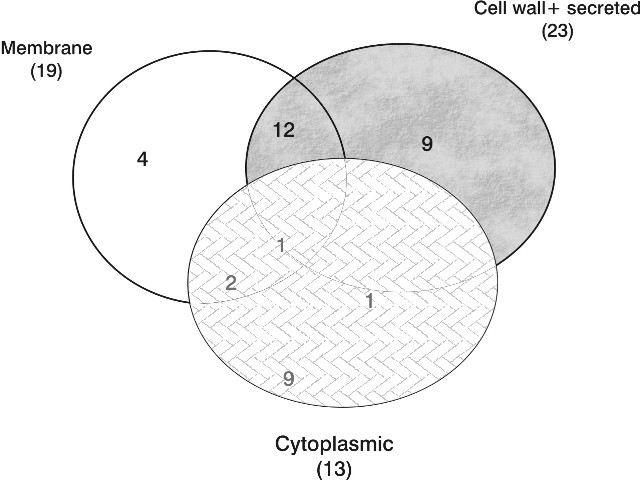
Subcellular localization prediction provides important information about a protein that allows making inferences regarding to the protein's function, to annotate genomes, and particularly in the case of bacterial pathogens to identify potential diagnostic, drug and vaccine targets. The distribution of subcellular location of 38 selected proteins are shown in Figure 1.
Searching for identical epitopes on M. smegmatis
The regions containing T and B epitopes at the same time of 38 selected M. tuberculosis proteins were the bases to look for identical epitopes on M. smegmatis. In these regions, 1367 T-cell epitopes and 140 B-cell epitopes were identified on M. tuberculosis. Using BLAST a search was made on the whole genome of M. smegmatis. 68 identical T-cell epitopes and 11 identical B-cell epitopes on 18 proteins of M. smegmatis were recorded from the alignments.
From the prediction, 5 proteins are secreted, 8 proteins are located in cytoplasm and 5 proteins locate in all of the cell compartments of the bacteria.
It is interesting to note that all the 18 identified M. smegmatis proteins containing the identical epitopes with M. tuberculosis have assigned functions reported in the Reference Sequence database at NCBI that correlate with the functions of the 13 M. tuberculosis identified target-proteins. This finding is consistent with the close similarity between M. smegmatis and M. tuberculosis at the genomic and functional levels. This similarity is the bases of the use of M. smegmatis as a model for the studies of the biology of M. tuberculosis and for the development of drugs against tuberculosis.
Regarding the proteins of M. tuberculosis containing common epitopes with M. smegmatis, the distribution according to the functional categories of the 13 identified target-proteins is shown in Figure 2. These 13 proteins mainly relate to the intermediary metabolism and respiration; virulence, detoxification, adaptation; lipid metabolism; cell wall and cell processes; conserved hypothetical functions of tubercle bacilli that were demonstrated highly expressed and important for the bacteria to adapt to different host conditions during different stages of infection (11, 12, 16-19, 21, 22).
Figure 1. A Venn diagram of the subcellular distribution of 38 identified proteins of M. tuberculosis.
Figure 2. Classification of M. tuberculosis proteins according to functional categories. The distribution according to functional categories of the selected 38 highly expressed genes in vivo and of the subgroup of these genes sharing identical epitopes with proteins of M. smegmatis (13 target-proteins) is shown. The distribution of genes of M. tuberculosis whole genome is shown as reference.
Evaluation of the immunogenicity of M. smegmatis and the cross-reactivity with M. tuberculosis on immunized mice
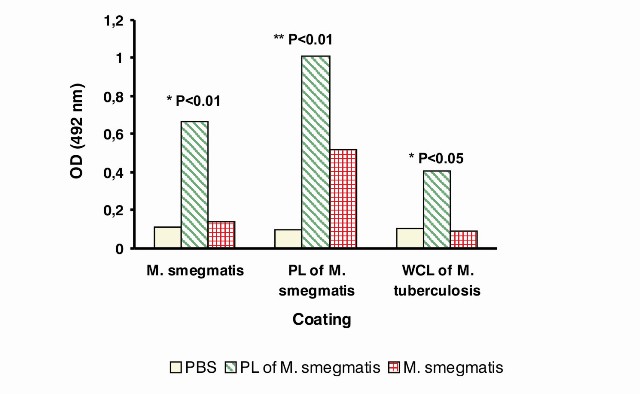
In vivo studies to evaluate the immunogenicity and the cross-reactivity of two vaccine candidates from M. smegmatis: proteoliposomes (PL) and live cells were carried out by an indirect ELISA to determine the recognition M. smegmatis and M. tuberculosis antigens by IgG antibodies of mice immunized with these two experimental vaccines. The results are shown in Figure 3.
Regarding the immunogenicity study, there was a statistical increase of the specific antibodies in the group of mice immunized with PL of M. smegmatis against whole cells of M. smegmatis and whole cell lysate of M. tuberculosis compared with the other groups (Figure 3). In the case of the responses against PL of M. smegmatis, animals immunized with PL had statistical increases of antibodies compared with the negative control group (mice immunized with PBS) (Figure 3). These results showed a better immunogenicity of PL with the adjuvant and immunization schedule used.
In the case of mice immunized with live cells of M. smegmatis, there were not statistically significant differences with negative control group against all the antigens tested. In this case, whole cells or PL of M. smegmatis and whole cell lysate of M. tuberculosis were not recognized by the antibodies of mice immunized with live cells of M. smegmatis. It should be taken into account that only one dose of immunization was used (as usually used of BCG vaccination). In a study using M. smegmatis as carrier vector for the expression of HIV proteins a more intensive immunization schedule was used with repeated immunizations in consecutive days (4).
DISCUSSION
The expression of different genes during in vivo infection is very important for the rational design of vaccines, considering that only antigens expressed in vivo will have an effective interaction with the immune system and enable to elicit protective immune responses, so the study of this group of antigens should have the priority for the identification of relevant antigens/epitopes to be included in new vaccines.
One of the main objectives of this study was to identify M. tuberculosis genes that are highly expressed in vivo in order to establish a pre-selected pool of antigens for the search of potential protective epitopes. In our case, we searched for genes expressed in vivo in humans and animals and in all phases of infection.
This is important in all strategies considering the need to have the correlation between the animal model and human, and on the other hand to have the inclusion of epitopes able to be operative in all the stages of the infection where vaccination could be used (primary infection, latency and clinical infection).
Figure 3. Study of the immunogenicity and cross-reactivity in mice of two vaccine candidates from M. smegmatis. Serum samples of three groups of mice immunized with PBS (negative control), proteoliposomes (PL) of M. smegmatis and M. smegmatis live cells (n=5 for each group) were collected 42 days after the first injection and analyzed by ELISA for the presence of specific IgG antibodies against: M. smegmatis whole cells [A], PL of M. smegmatis [B] and whole cell lysate (WCL) of M. tuberculosis [C]. Data are plotted as the mean of optical density (OD) measurement at 492 nm. (*) denotes a statistically significant difference between PL vs. all groups, (**) denotes a statistically significant difference between PL vs. PBS.
The distribution of the 38 highly expressed genes found in this study is relevant for the development of vaccine candidates targeting these antigens because they represent proteins of high importance for the survival of the bacteria during different phases of infection. Consequently, the immune responses elicited against these proteins should have a detrimental effect upon the bacteria, thus contributing to the protection. On the other side, the high representation of the cell wall component proteins gives the opportunity to stimulate specific humoral immune responses. In this respect, the potential of humoral antibodies in the protection against mycobacterial infections was demonstrated (31).
The potential advantages of M. smegmatis as carrier or experimental vaccine against tuberculosis is related to the fact that it is rapidly destroyed by phagolysosomal proteases in the phagosomes of infected cells (32-34), which is different in other mycobacterial strains such as BCG that survive in host cells for months by inhibiting phagosome maturation. Nevertheless, M. smegmatis can induce cytokine production by macrophages better than pathogenic mycobacterial species and can activate and induce the maturation of dendritic cells better than BCG by upregulation of major histocompatibility complex (MHC) class I and costimulatory molecules. M. smegmatis can also access the MHC class I pathway for presentation of mycobacterial antigens more efficiently than BCG (4).
The recognition of M. tuberculosis whole cell lysate by animals immunized with PL of M. smegmatis demonstrated the humoral cross-reactivity between M. smegmatis and M. tuberculosis which matches the results obtained by in silico studies where five identical B-cell epitopes were identified from secreted, cell wall and membrane proteins of M. smegmatis compared with M. tuberculosis.
It should be considered that, in addition to the linear epitopes identified by the prediction algorithm, there could be identical conformational B-cell epitopes between M. smegmatis and M. tuberculosis reinforcing the humoral cross-reactivity. Beside, we could detect the humoral reactivity against lipid and carbohydrate components that are present in the vaccine candidates used in this study and in the different preparations used for the coating of the ELISA systems (PL, whole M. smegmatis and whole cell lysate of M. tuberculosis). In recent studies carried out in our laboratory (29) and in collaboration with NIBSC, UK (unpublished results) we demonstrated the humoral immunogenicity of mycobacterial lipidic components in humans. In the case of M. smegmatis it was revealed by the cryo-electron microscope that the outermost layer of M. smegmatis is an outer membrane with a bilayer structure of lipids (30). These components could be recognized by the humoral immune responses.
In the in vivo study, it was only studied the humoral cross-reactivity between M. smegmatis and M. tuberculosis, so it could not be ruled out the presence of cross-reactivity at cellular level after the immunization with the M. smegmatis vaccine candidates, a fact that should be explored in future studies taking into account the relevant role assigned to the cellular immune response in the protection against tuberculosis.
Studies carried out in our laboratory have demonstrated the recognition of proteins of M. smegmatis by IgG antibodies from pulmonary tuberculosis patients in a whole cell ELISA assay. In those studies the levels of specific antibodies in tuberculosis patients had a statistical increase compared with healthy controls (unpublished results). The result demonstrated that during in-vivo infection in humans M. tuberculosis expresses epitopes present in M. smegmatis, which matches the in silico and in vivo results obtained in current study, in addition it supports the idea of the potential use of M. smegmatis for the development of TB vaccine candidates.
Acknowledgements
This work was partially supported by the MOSTI Grant No:07-01-05-MEB007.
REFERENCES
1. World Health Organization. Global tuberculosis control: surveillance, planning, financing: WHO report 2008. WHO Library Cataloguing-in-Publication Data:1-37.
2. Dietrich J, Lundberg CV, Andersen P. TB vaccine strategies - What in needed to solve a complex problem? Tuberculosis 2006;86:163-8.
3. Davies MN, Flower DR. Harnessing bioinformatics to discover new vaccines. Drug Discovery Today 2007; 12:389-96.
4. Cayabyab MJ, Hovav AH, Hsu T, Krivulka GR, Lifton MA, Gorgone DA, et al. Generation of CD8+ T-Cell responses by a recombinant nonpathogenic Mycobacterium smegmatis vaccine vector expressing human immunodeficiency virus type 1 Env. J Virology 2006; 56:1645-52.
5. Mawuenyega KG, Forst CV, Dobos KM, Belisle JT, Chen J, Bradbury EM, et al. Mycobacterium tuberculosis functional network analysis by global subcellular protein profiling. Molecular Biology of the Cell 2004; 16:396-404.
6. Wallis RS, Perkins M, Phillips M. Induction of the antigen 85 complex of Mycobacterium tuberculosis in sputum: A determinant of outcome in pulmonary tuberculosis treatment. J Infectious Diseases 1998;178:1115-21.
7. Fenhall G, Stevens L, Moses L, Bezuidenhout J, Betts JC. In Situ detection of Mycobacterium tuberculosis transcripts in human lung granulomas reveals differential gene expression necrotic lesions. IAI 2002; 70:6330-8.
8. Timm J, Post FA, Gail L. Differential expression of iron-, carbon, and oxygenresponsive mycobacterial genes in the lungs of chronically infected mice and tuberculosis patients. PNAS 2003; 100:14321-6.
9. Shi L, Jung YJ, Tyagi S. Expression of Th1-mediated immunity in mouse lungs induces a Mycobacterium tuberculosis transcription pattern characteristic of nonreplicating persistence. PNAS 2003; 100:241-6.
10. Shi L, North R, Gennaro ML. Effect of growth state on transcription levels of genes encoding major secreted antigens of Mycobacterium tuberculosis in the mouse lung. IAI 2004; 72:2420-4.
11. Talaat AM, Lyons R, Howard ST, Johnston SA. The temporal expression profile of Mycobacterium tuberculosis infection in mice. PNAS 2004; 101:4602-7.
12. Karakousis PC, Yoshimatsu T, Lamichhane G, Woolwine SC, Nuermbeger EL. Dormancy phenotype displayed by extracellular Mycobacterium tuberculosis within artifical granulomas in mice. J Exp Med 2004; 200: 647-57.
13. Tufariello JM, Jacobs WR, Chan J. Individual Mycobacterium tuberculosis resuscitation-promoting factor homologues are dispensable for growth in-vitro and in-vivo. Infect Immun 2004; 72:515-26.
14. Dubnau E, Chan J, Mohan VP, Smith I. Responses of Mycobacterium tuberculosis to growth in the mouse lung. IAI 2005; 73:3754-7.
15. Delogu G, Sanguinetti M, Posteraro B. The hbhA gene of Mycobacterium tuberculosis is specifically upregulated in the lungs but not in the spleens of aerogenically infected mice. IAI 2006; 74:3006-11.
16. Rachman H, Strong M, Ulrichs T, Grode L, Schuchhardt J. Unique transcriptome signature of Mycobacterium tuberculosis in pulmonary tuberculosis. IAI 2006;74:1233-42.
17. Talaat AM, Ward SK, Wu CW, Rondon E. Mycobacterial bacilli are metabolically active during chronic tuberculosis in murine lungs: Insights from genome-wide transcriptional profiling. J Bacteriology 2007; 189:4265-74.
18. Garton NJ, Waddell SJ, Sherratt AL, Lee SM, Smith RJ. Cytological and transcript analyses reveal fat and lazy persisterlike bacilli in tuberculous sputum. PLoS Med 2008; 5:1371-82.
19. Rustad TR, Harrell MI, Liao R, Sherman DR. The enduring hypoxic response of Mycobacterium tuberculosis. PloS ONE 2008;7:1502-9.
20. Srivastava V, Jain A, Srivastava BS. Selection of genes of Mycobacterium tuberculosis upregulated during residence in lungs of infected mice. Tuberculosis 2008; 88:171-7.
21. Kesavan AK, Brooks M, Tufariello JA, Chan J, Manabe YC. Tuberculosis genes expressed during persistence and reactivation in the resistant rabbit model. Tuberculosis 2009; 89:17-21.
22. Davies AP, Dhillon AP, Young M. Resuscitation-promoting factors are expressed in Mycobacterium tuberculosis-infected human tissue. Tuberculosis 2008;88:462-8.
23. Álvarez N, Borrero R, García MA, Martínez I, Acosta M, Padrón MA, et al. Obtención y caracterización parcial de un extracto lipídico de la membrana externa de Mycobacterium smegmatis. VacciMonitor 2009;18:15-9.
24. Zhang M, Kim KJ, Iyer D, Lin YG, Belisle J, McEnery K, et al. Effects of Mycobacterium tuberculosis on the bioelectric properties of the alveolar epithelium. Infect Immun 2007; 65:692-8.
25. Olivares N, León A, López Y, Puig A, Cádiz A, Falero G, et al. The effect of the administration of human gamma globulins in a model of BCG infection in mice. Tuberculosis 2006; 86:268-72.
26. Kuehnel MP, Goethe R, Habermann A, Mueller E, Rodeh M, Grifiths S, Valentin-Weigand M. Characterization of the intracellular survival of Mycobacterium avium ssp. paratuberculosis: phagosomal pH and fusogenicity in J774 macrophages compared with other mycobacteria. Cell Microbiol 2001; 3:551-66.
Recibido: Mayo de 2009
Aceptado: Diciembre de 2009


{kind=link}
{kind=link}
{kind=link}