










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCuban Journal of Agricultural Science
versión On-line ISSN 2079-3480
Cuban J. Agric. Sci. vol.51 no.2 Mayabeque abr.-jun. 2017
ARTÍCULO ORIGINAL
Estatus racial e índices de infestación de colmenas (Apis mellifera L.) por Varroa destructor (Anderson y Trueman) en Mayabeque, Cuba
Racial status and index of hive (Apis mellifera L.) infestation by Varroa destructor (Anderson and Trueman) in Mayabeque, Cuba
Anisley Pérez and J. Demedio
Universidad Agraria de La Habana “Fructuoso Rodríguez Pérez”. Apartado Postal 18-19, CP 32700, San José de Las Lajas, Mayabeque, Cuba
RESUMEN
Con el objetivo de evaluar el estatus racial e índices de infestación de colmenas (Apis mellifera L.) y las tasas de infestación por Varroa destructor en abejas adultas (TIA), se evaluaron rasgos fenotípicos y genotípicos importantes de la abeja melífera en un Centro de Cría de Abejas Reinas de Mayabeque, Cuba. Para ello se determinó la coloración de muestras de abejas obreras por la escala de amarillas, intermedias y negras. Mediante el método de la regla fija se realizó un estudio biométrico de las obreras de esas mismas colmenas y del diámetro de las celdas de obreras y se evaluaron las tasas de infestación por el ácaro Varroa destructor (TIA). Las variables coloración, longitud del ala anterior derecha y diámetro de las celdas de obreras, en conjunto, unidas al comportamiento defensivo habitual indicaron que el fenómeno de la africanización no está presente en las colmenas estudiadas. Al tomar como referencia los trabajos de los años 80, se percibe reducción del ala anterior y ennegrecimiento de estas abejas como consecuencia de estar sometidas a un proceso de selección para “tolerancia a Varroa”, desarrollado por el hombre y la influencia de la naturaleza. Esta última característica procede, obviamente, de la más rústica raza original Apis mellifera mellifera. Los rasgos evaluados y las tasas de infestación por Varroa, que presentaron valores por encima de 5 %, se corresponden con los de las razas europeas que dieron origen a la actual abeja cubana.
Palabras clave: Apis mellifera, estatus racial, varroosis, africanización
ABSTRACT
In order to evaluate the racial status and index of hive infestation (Apis mellifera L.) and infestation rates of Varroa destructor in adult bees, important phenotypic and genotypic traits of the honeybee were evaluated in the Centro de Cría de Abejas Reinas (CCR) in Mayabeque, Cuba. For this purpose, the coloration of worker bee samples was determined according to yellow, intermediate and black scale. Using the fixed rule method, a biometric study of workers from these same hives and the diameter of cell for workers was carried out and infestation rates of Varroa destructor were evaluated. The variables coloration, length of the right forewing and diameter of the worker cells, together with the usual defensive behavior, indicated that the phenomenon of Africanization is not present in the studied hives. Taking studies of the 80s as a reference, there is a reduction of the forewing and a blackening of these bees as a result of being subjected to a selection process for "tolerance to Varroa", developed by humans and the influence of nature. This last characteristic comes, obviously, from the most ancient original breed Apis mellifera mellifera. Evaluated traits and infestation rates of Varroa, which showed values over 5 %, correspond to those of European breeds that gave origin to the current Cuban bee.
Key words: Apis mellifera, racial status, varroosis, Africanization
INTRODUCCIÓN
La abeja melífera occidental Apis mellifera, originaria de Europa, África y Asia Sudoriental, fue clasificada por Carolus Linnaeus en 1758. Debido a los diferentes factores ambientales existentes en cada región, se originaron conjuntos de individuos que, aunque pertenecen a la misma especie, se adaptaron a un medio particular y originaron grupos, denominados razas geográficas, subespecies o ecotipos. La abeja negra o alemana (Apis mellifera mellifera), la italiana o amarilla (Apis mellifera ligustica), la carniola o cárnica (Apis mellifera carnica) y la caucásica o Apis mellifera caucásica son las principales razas geográficas introducidas al nuevo mundo por los colonizadores (Civantos et al. 2012).
En un programa para el desarrollo de la apicultura de Brasil a cargo de Warwick Kerr, se introdujeron en 1956 abejas de la raza africana Apis mellifera scutellata y un año después, se escaparon algunos enjambres que se cruzaron libremente con la población de abejas europeas existente. Se originó así el agresivo híbrido, conocido como abeja africanizada, que se ha extendido por casi toda América (Verde et al. 2012).
La dispersión del híbrido introdujo cambios importantes en la apicultura. Inicialmente se produjo un descenso en la producción de miel, debido al abandono de los apicultores por el aporte africano (Ratcliffe et al. 2011) que se explica por el comportamiento defensivo (agresivo) de estos híbridos. Más tarde, cuando se elaboraron mejores programas de manejo de estas nuevas abejas se lograron resultados productivos superiores a los obtenidos hasta el momento con sus parientas europeas. Es esencial entonces conocer el grado de africanización que tienen las abejas para poder tomar mediadas de manejo adecuadas. La morfometría es un instrumento que ayuda a reconocer las diferentes especies y razas (Pereira et al. 2013).
Con el propósito de determinar el grado de africanización de las abejas, se han propuesto diferentes análisis morfométricos (Rinderer et al. 1993). Algunos, como el método Fabis (Sylvester y Rinderer 1987), se tornan difíciles desde el punto de vista económico y práctico, pues tienen en cuenta muchas características. En su lugar, se ha propuesto otro método más sencillo, que incluye solo la longitud del ala anterior derecha, por ser el rasgo único que mejor discrimina entre europeas y africanizadas (Rinderer et al. 1986). Se ha informado que las medias de la longitud del ala anterior derecha son inferiores para las abejas africanizadas (Souza et al. 2009).
Al estar Cuba ubicada en un área geográfica donde están ampliamente extendidos estos híbridos, y al existir antecedentes de introducción intencional de material biológico dañino, es ilógico esperar la presencia de este híbrido en el país (Demedio 2001). Por ello, es evidente la necesidad de determinar, al menos, variables morfológicas (Medina-Flores et al. 2014) que permitan descartar la posible presencia de esos híbridos en Cuba y que, unido a la coloración, sirvan de base para la comparación entre colmenas de diverso comportamiento higiénico, de acicalamiento, defensivo y otros, en correspondencia con los índices de infestación del ácaro Varroa. A partir de estas premisas, el objetivo de este estudio fue determinar el estatus racial y los índices de infestación por V. destructor de colmenas (Apis mellifera L.) provenientes de Mayabeque, Cuba.
MATERIALES Y MÉTODOS
La investigación se llevó a cabo en 15 colmenas (10 maternas y 5 paternas) del Centro de Cría de Abejas Reinas de Mayabeque durante febrero, marzo, abril y mayo de 2007, y febrero y mayo de 2008. Se efectuaron tres estudios: 1) determinación de la coloración de muestras de obreras, basada en las categorías amarilla, intermedia y negra (Voroshilov 2008); 2) estudio biométrico de obreras de esas mismas colmenas por la longitud del ala anterior derecha (Rinderer et al. 1986 y Souza et al. 2009) y del diámetro de las celdas de sus panales de cría de obreras; 3) determinación de las tasas de infestación por V. destructor en abejas adultas (TIA). Este Centro produce aproximadamente 2 600 reinas por año y cubre más del 50 % de la demanda provincial.
En cada ensayo se tomaron muestras al azar de 50 abejas por colmena (Morales-Pérez 2012), depositadas en pomos con tapas horadadas, previamente identificados. Las muestras se sometieron a congelación como método de sacrificio de las abejas. Posteriormente, se determinó su coloración sobre la base de tres categorías (amarillo, negro e intermedio), para lo que se utilizó una lupa simple. Se tomaron como base los colores de las razas de abejas que dieron origen a las abejas existentes en Cuba en la actualidad. Las colmenas siempre se trabajaron con los medios de protección utilizados habitualmente en el país.
Para medir el ala anterior derecha, se desprendió y se montó en láminas portaobjetos con bálsamo de Canadá, a razón de 25 alas por lámina. Se dejaron secar durante 48-72 h para medirlas posteriormente en un microscopio estereoscópico con ocular micrométrico.
En correspondencia con los meses de evaluación, se realizó el diagnóstico de las tasas de infestación por Varroa en abejas adultas, según el método clásico de De Jong et al. (1982), a partir de muestras de ≥ 200 abejas adultas por colmena (OIE 2008) tomadas al azar, para establecer su posible relación con los patrones de coloración de las obreras de estas colmenas.
Los datos se procesaron mediante estadística descriptiva y las medias se compararon por el Test de Rangos Múltiples (Duncan 1955) y el de comparación de proporciones del paquete Statgraphics Plus 5.1 (Statistical Graphics Crop 2000).
RESULTADOS Y DISCUSIÓN
Las colmenas mostraron su actitud defensiva habitual reconocida por los apicultores. Se constató en el CCR un claro predominio del color negro, prácticamente como único puro, con casi nula frecuencia del propiamente amarillo y gran población de individuos que clasifican como intermedios por la presencia de ambos colores en los segmentos abdominales (figura 1). Llama la atención el mantenimiento de las proporciones de abejas de color negro, entre 62 % y 70 %, en las colmenas maternas durante los meses estudiados. Algo diferente ocurrió en las paternas, originarias de la zona, entre las que el color negro cayó significativamente a 38 % en mayo de 2008. Lo más probable es que esto se pueda explicar por sustituciones de las reinas originales y, es menos posible, que se pueda atribuir a la alternancia en la utilización por parte de las reinas de espermatozoides de diferentes zánganos (Verde et al. 2012).

Se observaron fluctuaciones dentro de los grupos paternos y maternos. Este comportamiento fue observado por Sanabria (2004) y Díaz-Rieumont (2006) en un apiario de selección de la zona y se puede considerar una evidencia de la alternancia de utilización de paquetes de espermatozoides de diferentes zánganos. No se descarta la participación de individuos de apiarios manejados o silvestres en un radio que, según Pérez-Piñeiro (2007), puede superar los 10 km.
En otro estudio, Sanabria (2007) observó porcentajes de abejas negras entre 36 y 42%, pero en un apiario experimental sin tratamiento, claramente inferiores al 48 y 59 %. Es significativa la presencia de abejas negras en este CCR, especialmente en las colmenas maternas, pero también en las paternas. Esto se pudiera explicar a partir de la preservación de las reinas maternas originales (negras) procedentes del apiario de selección del Centro de Investigaciones Apícolas.
Se observan además diferencias entre las medias de la longitud del ala anterior derecha de las obreras provenientes de colmenas maternas y paternas (figura 2). Las menores dimensiones correspondientes a enero deben tener su origen en el efecto ambiental que impone el envejecimiento de panales (Aguirre 2005), más que causas de origen genético. El retraso en la renovación de la cámara provoca reducción de la capacidad de las celdas de cría y en consecuencia, el desarrollo de abejas más pequeñas (Verde et al. 2013), aunque en este caso, muy superiores a los valores en abejas africanizadas (Sanabria 2007).

En los primeros trabajos biométricos realizados en Cuba hace más de 20 años (Díaz-Millán 1981a,b y Díaz-Millán y Domínguez 1981), se determinó una media nacional de 9.60 mm para abejas negras y para las amarillas, con mínimo de 9.50 mm en zonas de la región central y 9.70 mm en Sagua de Tánamo, Guantánamo. Se tuvo en cuenta que estuvieran por encima de las medidas informadas para las razas que introducidas originalmente en Cuba. Si hoy se obtuvieron las medias que aparecen en la figura anterior, se ha producido un evidente acortamiento de esta variable, lo que pudiera explicarse por dos factores. En primer lugar, no se tuvo en cuenta la edad de los panales de la cámara de cría, ya que cuanto más viejos producen abejas de menor tamaño (Piccirillo y De Jong 2003 y Sanabria 2007). El otro factor a considerar, sobre el que no se han localizado estudios precedentes, está relacionado con el efecto de la varroosis y su segura modificación del genofondo poblacional, debido al gran número de colmenas que perecieron en la etapa inicial y también con los procesos de selección que se han desarrollado sobre la base de la tolerancia al parásito, expresada por bajas tasas de infestación, y la productividad, que ocupa el lugar primordial (MINAG 2014).
A pesar de las reducciones señaladas, aún la media menor de las paternas del CCR está por encima de las halladas en Latinoamérica en poblaciones africanizadas. En Colombia, Salamanca et al. (2001, 2004) informaron medias de 8.50 mm – 8,74 mm. Ruttner (1988) obtuvo en España valores de 9.41 mm y 9.82 mm en A. mellifera iberica. En localidades de La Palma, Islas Canarias, y la Península Ibérica, Padilla et al. (2001a, b) refirieron medias de 9.41 mm – 9.82 mm. En México Aguirre (2005) consideró no africanizadas aún a abejas con medias de 9.19 mm a 9,29 mm. Estas medidas, relativamente bajas, las atribuyó al envejecimiento de la cámara de cría Las medias de 9.22 mm (Portugal y España), 9.09 mm (Marruecos) y 9.28 mm (Francia), determinadas por Arias et al. (2006), no correspondieron a africanizadas cuando se contrastaron por pruebas de electroforesis (MDH-1) o ADNmt.
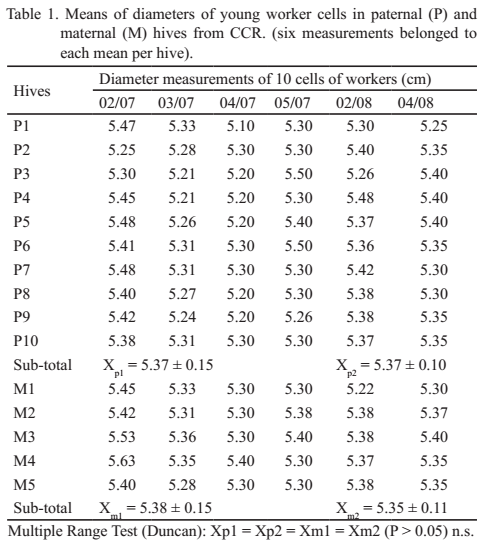
Desde hace tiempo, se sabía que razas africanas como A. mellifera adansonii y A. mellifera scutellata construyen celdas más pequeñas (Guzmán-Novoa et al. 2011), por lo que esta característica se ha considerado en el Programa de Control de la Abeja Africanizada en México. Resulta evidente en este trabajo (tabla 1) que todas las longitudes medias estén por encima del nivel correspondiente a las abejas europeas (5.20 mm), y por encima de las que corresponden a las africanizadas o africanas (4.90 mm).
Otros resultados obtenidos en Brasil (De Jong 2001, Piccirillo 2001 y Piccirillo y De Jong 2003) y por autores norteamericanos (Hall et al. 2015) apoyan lo antes referido, por cuanto las africanas no sobrepasan los 4.90 mm en panales naturales y 5.20 mm en obra sobre láminas de cera estampada para europeas, que son las utilizadas en Cuba. Precisamente en esta zona del país, Sanabria et al. (2015) determinaron una media superior a 5.27 mm, lo que unido a otras características le permitió concluir que no existía africanización.
La tasa media de infestación en abejas adultas de las colmenas maternas (270 ácaros/5 224 abejas) no difirió respecto a las de colmenas paternas (574 ácaros/10 780 abejas) (figura 3). La explicación a este comportamiento resulta difícil, debido a la preferencia de las hembras fundadoras del ácaro Varroa por las celdas de zánganos y por su mayor éxito reproductivo aquí; además de que son precisamente las colmenas paternas las encargadas de aportar los machos para la fecundación en el CCR, lo que supone una elevada cantidad de cría de zánganos en ellas (Verde et al. 2013).

Esta situación, que a primera vista podría evaluarse como favorable, se debe a la poca presencia de cría de zánganos en las colmenas paternas, que de modo natural deben experimentar un incremento más acelerado del parasitismo. A esto se añade que las maternas provienen de un apiario de selección, en el cual se toma como criterio el mantenimiento de bajas tasas de infestación, sin el auxilio de tratamientos.
Las tasas medias de infestación por encima de 5%, considerado el umbral de peligro en Cuba (Verde et al. 2013) indican la existencia de colmenas con valores por encima de ese nivel. Esto no se corresponde con las situaciones informadas para colmenas africanizadas de América Continental (Maggi et al. 2012 y Galindo-Cardona 2013), de manera que constituye un elemento adicional en correspondencia con el estatus racial de origen europeo de la abeja cubana actual. Los rasgos biométricos, la conducta defensiva apreciada y la tasa de infestación por Varroa destructor, están en correspondencia con el estatus de origen europeo de las colmenas estudiadas.
REFERENCIAS
Aguirre, J. L. 2005. La varroasis en colmenas de Baja California Sur. El agente etiológico y opciones para su control. Ph.D. Thesis, Universidad Autónoma de Baja California Sur – Universidad Agraria de La Habana, México - Cuba, 113 p.
Arias, M. C., Rinderer, T. E. & Sheppard, W. S. 2006. “Further characterization of honey bees from the Iberian Peninsula by allozyme, morphometric and mtDNA haplotype analyses”. Journal of Apicultural Research, 45(4): 188–196, ISSN: 0021-8839, DOI: 10.1080/00218839.2006.11101346.
Civantos, E., Thuiller, W., Maiorano, L., Guisan, A. & Araújo, M. B. 2012. “Potential Impacts of Climate Change on Ecosystem Services in Europe: The Case of Pest Control by Vertebrates”. BioScience, 62(7): 658–666, ISSN: 0006-3568, DOI: 10.1525/bio.2012.62.7.8.
De Jong, D. 2001. “Genética aplicada a los programas de selección”. In: VII Congreso Internacional de Actualización Apícola - I Foro de Vinculación Apícola, Veracruz, México: Asociación Nacional de Médicos Veterinarios Especialistas en Abejas (ANMVEA) – Universidad Veracruzana, pp. 83–87.
De Jong, D., Roma, D. de A. & Goncalves, L. S. 1982. “A comparative analysis of shaking solutions for the detection of Varroa jacobsoni on adult honeybees”. Apidologie, 13(3): 297–306, ISSN: 0044-8435, 1297-9678.
Demedio, J. 2001. La varroasis de las abejas en una zona de la provincia de La Habana. Agente etiológico, índices de infestación y control biotécnico y químico. Ph.D. Thesis, Universidad Agraria de La Habana, La Habana, Cuba, 107 p.
Díaz-Millán, M. 1981a. “Características morfológicas de la abeja (Apis mellifica) en las diferentes regiones de Cuba, donde se seleccionarán colmenas para constituir apiarios de reserva”. Agrotecnia de Cuba, 13(1): 8–17, ISSN: 2414-4673, 0568-3114.
Díaz-Millán, M. 1981b. “Estudio morfométrico de la abeja (Apis mellifica) en la antigua provincia de Las Villas”. Agrotecnia de Cuba, 13(2): 14–22, ISSN: 2414-4673, 0568-3114.
Díaz-Millán, M. & Domínguez, D. A. 1981. “Estudio morfométrico de la abeja (Apis mellifica) en la antigua provincia de Las Villas”. Agrotecnia de Cuba, 13(2): 121–131, ISSN: 2414-4673, 0568-3114.
Díaz-Rieumont, S. 2006. Estudio del comportamiento de los índices de infestación por Varroa destructor, su relación con dos mecanismos conductuales de la abeja melífera (Apis mellifera L.) y sus características morfológicas. Graduated Thesis, Universidad Agraria de La Habana, La Habana, Cuba, 71 p.
Duncan, D. B. 1955. “Multiple Range and Multiple F Tests”. Biometrics, 11(1): 1–42, ISSN: 0006-341X, DOI: 10.2307/3001478.
Galindo-Cardona, A. 2013. “Especies Invasoras. Apis mellifera L., de África vinieron todas”. Boletín de la Sociedad Entomológica Argentina, 23(1): 3–4, ISSN: 1666-4612, 1515-1557.
Guzmán-Novoa, E., Correa-Benítez, A., Espinosa-Montaño, L. G. & Guzmán-Novoa, G. 2011. “Colonización, impacto y control de las abejas melíferas africanizadas en México”. Veterinaria México, 42(2): 149–178, ISSN: 0301-5092.
Hall, H. G., Zettel-Nalen, C. & Ellis, J. D. 2015. African Honey Bee: What You Need to Know. EDIS, University of Florida, Available: <http://edis.ifas.ufl.edu/mg113>, [Consulted: July 4, 2017].
Maggi, M., Medici, S., Quintana, S., Ruffinengo, S., Marcángeli, J., Martinez, P. G., Fuselli, S. & Eguaras, M. 2012. “Genetic structure of Varroa destructor populations infesting Apis mellifera colonies in Argentina”. Experimental and Applied Acarology, 56(4): 309–318, ISSN: 0168-8162, 1572-9702, DOI: 10.1007/s10493-012-9526-0.
Medina-Flores, C. A., Guzmán-Novoa, E., Hamiduzzaman, M. M., Aréchiga-Flores, C. F. & López-Carlos, M. A. 2014. “Africanized honey bees (Apis mellifera) have low infestation levels of the mite Varroa destructor in different ecological regions in Mexico”. Genetics and Molecular Research, 13(3): 7282–7293, ISSN: 1676-5680.
MINAG (Ministerio de la Agricultura de Cuba) 2014. Programa de mejoramiento genético de la abeja Apis mellifera con abejas localmente adaptadas al ácaro Varroa destructor. Cuba: CIAPI.
Morales-Pérez, G. E. 2012. Detección de Abeja Africana (Apis mellifera scutellata) en la Comarca Lagunera. Graduated Thesis, Universidad Autónoma Agraria Antonio Narro, Coahuila, México, 46 p.
OIE (Office International des Épizooties) 2008. Manual of diagnostic tests and vaccines for terrestrial animals: mammals, birds, and bees. 6th ed., Paris: Office International des Épizooties, 1343 p., Available: <http://books.google.com/books?id=xmZWAAAAYAAJ>, [Consulted: July 4, 2017].
Padilla, A. F., Hernández, F. R. & Reyes, L. J. 2001a. “Estudio biométrico de la abeja melífera (Apis mellifera, Linneo 1758) (Hymenoptera, Apidae) de la isla de La Palma del Archipiélago Canario. II. Ángulos y longitudes de las alas.”. Zoología Baética, 12: 23–35, ISSN: 1130-4251.
Padilla, F., Valerio, M. J., Campano, F., Jiménez, E., Puerta, F., Flores, J. M. & Bustos, M. 2001b. “Estudio biométrico de poblaciones de abejas (Apis mellifera L.) del centro de Portugal y de Madeira”. Archivos de Zootecnia, 50(190): 67–77, ISSN: 0004-0592.
Pereira, R. A., Morais, M. M., Francoy, T. M. & Gonçalves, L. S. 2013. “Hygienic Behavior of Africanized Honey Bees Apis mellifera Directed towards Brood in Old and New Combs during Diurnal and Nocturnal Periods”. Insects, 4(4): 521–532, ISSN: 2075-4450, DOI: 10.3390/insects4040521.
Pérez-Piñeiro, A. 2007. Manual de Apicultura. La Habana, Cuba: Agrinfor, 154 p., ISBN: 959-246-130-9.
Piccirillo, G. A. 2001. Efeito do tamanho da célula do favo de cria sobre a variabilidade morfológica das abelhas africanizadas (Apis mellifera) e sobre a infestação e reprodução do ácaro Varroa jacobsoni. M.Sc. Thesis, Universidade de São Paulo, Brasil, 133 p.
Piccirillo, G. A. & De Jong, D. 2003. “The influence of brood comb cell size on the reproductive behavior of the ectoparasitic mite Varroa destructor in Africanized honey bee colonies”. Genetics and Molecular Research, 2(1): 36–42, ISSN: 1676-5680.
Ratcliffe, N. A., Mello, C. B., Garcia, E. S., Butt, T. M. & Azambuja, P. 2011. “Insect natural products and processes: New treatments for human disease”. Insect Biochemistry and Molecular Biology, 41(10): 747–769, ISSN: 0965-1748, DOI: 10.1016/j.ibmb.2011.05.007.
Rinderer, T. E., Buco, S. M., Rubink, W. L., Daly, H. V., Stelzer, J. A., Riggio, R. M. & Baptista, F. C. 1993. “Morphometric identification of Africanized and European honey bees using large reference populations”. Apidologie, 24(6): 569–585, ISSN: 0044-8435, DOI: 10.1051/apido:19930605.
Rinderer, T. E., Sylvester, H. A., Brown, M. A., Villa, J. D., Pesante, D., Collins, A. M., Spencer, R., Kleinpeter, S. & Lancaster, V. 1986. “Field and simplified techniques for identifying africanized and european honey bees”. Apidologie, 17(1): 33–48, ISSN: 0044-8435, DOI: 10.1051/apido:19860104.
Ruttner, F. 1988. Biogeography and Taxonomy of Honeybees. Berlin, Heidelberg: Springer Berlin Heidelberg, ISBN: 978-3-642-72651-4, Available: <http://link.springer.com/10.1007/978-3-642-72649-1>, [Consulted: July 4, 2017].
Salamanca, G. G., Benítez, R., Montes, M. & Fernández, K. 2001. Estudio puntual del nivel de defensividad en colmenas de abejas Apis mellifera medido en distintas horas del día. Apiservices, Available: <http://www.apiservices.biz/es/articulos/ordenar-por-popularidad/729-estudio-puntual-del-nivel-de-defensividad>, [Consulted: July 4, 2017].
Salamanca, G., Tello, D. J. & Alentar-Arnaut, V. 2004. “25 años de la abeja africanizada a su paso por Colombia (1978-2003)”. In: I Congreso Cubano de Apicultura, La Habana, Cuba: APICUBA - MINAG.
Sanabria, J. L. 2004. Evaluación de dos mecanismos de defensa de las abejas Apis mellifera frente al ácaro Varroa destructor. M.Sc. Thesis, Centro Nacional de Sanidad Agropecuaria (CENSA), La Habana, Cuba, 54 p.
Sanabria, J. L. 2007. Índices de infestación, estatus racial y expresión de mecanismos de resistencia en colmenas sin control antivarroa. Ph.D. Thesis, Universidad Agraria de La Habana, La Habana, Cuba, 121 p.
Sanabria, J. L., Demedio, J., Pérez, T., Peñate, I., Rodríguez, D. & Lóriga, W. 2015. “Índices de infestación por Varroa destructor en colmenas sin medidas de control”. Revista de Salud Animal, 37(2): 118–124, ISSN: 0253-570X.
Souza, D. L., Evangelista-Rodrigues, A., Ribeiro, M. N., Padilla-Álvarez, F., Farias, E. S. L. & Pereira, W. E. 2009. “Análises morfométricas entre Apis mellifera da mesorregião do sertão paraibano”. Archivos de Zootecnia, 58(221): 65–71, ISSN: 0004-0592.
Statistical Graphics Crop 2000. STATGRAPHICS® Plus. (ser. Profesional), version 5.1, [Windows], Available: <http://www.statgraphics.com/statgraphics/statgraphics.nsf/pd/pdpricing> .
Sylvester, H. A. & Rinderer, T. E. 1987. “Fast Africanized bee identification system (FABIS) manual”. American Bee Journal, 127: 511–516, ISSN: 0002-7626.
Verde, M., Demedio, J. & Gómez, T. 2013. Apicultura. Salud y Producción. Guía técnica para el apicultor. 1st ed., vol. 2, Cuba: ACPA, 208 p., ISBN: 978-959-7190-21-9.
Verde, M., Gómez, T. & Demedio, J. 2012. Salud Apícola. Generalidades. 1st ed., vol. 1, Cuba: ACPA, 207 p., ISBN: 978-959-7190-15-8.
Voroshilov, U. 2008. Determinación del estatus racial y parasitológico (varroosis) de colmenas (Apis mellifera L.), ante la amenaza de la africanización. Graduated Thesis, Universidad Agraria de La Habana, La Habana, Cuba, 72 p.
Recibido: 21/6/2016
Aceptado: 20/6/2017
Anisley Pérez, Universidad Agraria de La Habana “Fructuoso Rodríguez Pérez”. Apartado Postal 18-19, CP 32700, San José de Las Lajas, Mayabeque, Cuba. Email: anisley_perez@unah.edu.cu


{kind=link}