










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkIntroducción
Pseudotsuga menziesii (Mirb.) Franco, es una especie con alto potencial comercial, debido a la calidad de su madera, producción de taninos, así como a su preferencia como árbol de navidad (Arteaga y Zenil, 2005). Está incluida entre las especies en peligro de extinción en la Norma Oficial Mexicana 059-SEMARNAT (SEMARNAT, 2010), con un estatus de protección especial. Por esta razón, es indispensable ayudar al bosque natural a regenerarse mediante reforestaciones e implementar plantaciones con esta especie de alto valor comercial (Arteaga y Zenil, 2005).
El consumo de árboles de navidad en México ha aumentado en los últimos años, comercializándose entre 1.6 y 2.0 millones de árboles anualmente. La balanza comercial con los Estados Unidos y Canadá es negativa, por lo que es importante incrementar la producción de este bien, mejorando la tecnología del proceso productivo.
La fertilización química es una actividad silvícola que en las plantaciones de árboles de navidad puede aumentar la tasa de crecimiento, el vigor del árbol, el color y la densidad del follaje, la resistencia a plagas y enfermedades y la retención de acículas; esto último, aún después de que el árbol ha sido cortado (CONAFOR, 2011).
Los recursos que determinan la productividad del sitio corresponden a los elementos que controlan procesos básicos del crecimiento de la planta a nivel fisiológico; entre estos se incluye la luz o radiación fotosintéticamente activa incidente, agua disponible, CO2 atmosférico, oxígeno (a nivel radicular y aéreo), temperatura y nutrimentos disponibles (Espinosa et al., 2017). Para lograr una adecuada tasa de crecimiento y desarrollo de la planta se deben suministrar los nutrimentos necesarios según la fertilidad de sitio y los requerimientos de la especie. Las limitaciones nutrimentales y climáticas siempre determinan el crecimiento potencial de los cultivos (Espinosa et al., 2017).
En Huayacocotla, Veracruz, existe una plantación de P. menziesii, establecida en agosto del año 2011 con fines de producción de árboles de navidad; sin embargo, el desempeño de esta especie en el área parece inadecuado y el lento crecimiento de los árboles en el sitio puede deberse a problemas de baja fertilidad del suelo u otros factores que afectan su crecimiento, tales como la temperatura, la precipitación, el tipo de suelo entre otros, aun cuando las especies nativas como Pinus patula Schl. et Cham., P. montezumae Lamb., P. pseudostrobus Lindl. y P. leiophylla Schl. et Cham. presentan tasas de crecimiento aparentemente normales. En el contexto anterior, esta investigación se realizó con el fin de evaluar los efectos de nitrógeno, fósforo, potasio y micronutrimentos en la reactivación del crecimiento de brinzales de P. menziesii, a cuatro años de su establecimiento y determinar si la reducida tasa de crecimiento de los brinzales de P. menziesii se puede enmendar mediante fertilización química con N, P, K y micronutrimentos.
Materiales y métodos
El experimento se realizó de agosto de 2015 a octubre de 2018, en la localidad de Palo Bendito, Huayacocotla, Veracruz, dentro una plantación de árboles de navidad de Pseudotsuga menziesii (Mirb.) Franco de cuatro años de edad. El predio se encuentra en las coordenadas geográficas 20º 7' 19.59'' N, 98º 29' 30.59'' O. En esta área predomina un clima templado húmedo con abundantes lluvias en verano, con una temperatura media anual entre 8 y 12 ºC y precipitación media anual de 1,379.7 mm (INEGI, 2009). Los suelos de la región se clasifican mayormente como cambisoles y regosoles (INEGI, 2009) (Figura 1).

Fig. 1 - Localización del área del experimento de fertilización de Pseudotsuga menziesii en el ejido "Palo Bendito", Huayacocotla, Veracruz
Diseño experimental y de tratamientos
Se condujo un experimento factorial 24 con una distribución de los tratamientos en forma completamente al azar. Los factores de estudio fueron: nitrógeno (N), fósforo (P), potasio (K) y micronutrimentos (Mi); cada factor se evaluó en dos niveles (dosis/árbol) como se muestra en la tabla #1. Las fuentes de N, P y K fueron urea (46-00-00), superfosfato de calcio triple (00-46-00, 21CaO) y sulfato de potasio (00-00-50), respectivamente; mientras que la fuente de micronutrimentos fue el producto comercial Sagaquel combi® (3.10 % Fe, 4.05 % Zn, 0.05 % B, 0.04 % Cu, 0.002 % Co, 0.25 % Mn, 5.332 % S, 1.05 % Mg, 0.005 % Mo). Los 16 tratamientos resultantes fueron replicados cinco veces y la unidad experimental fue un árbol. (Tabla 1)
Tabla 1 - Niveles de N, P, K y Micronutrimentos (dosis de material fertilizante por árbol) probados en el experimento de fertilización de Pseudotsuga menziesii en Huayacocotla, Veracruz
N=nitrógeno, P=fósforo, K=potasio
Las dosis de N, P y K se definieron en base a las concentraciones críticas existentes para Pseudotsuga menziesii publicadas por Zas (2003) y utilizando el principio del método racional de Rodríguez Suppo, adaptado para tejido vegetal.
La aplicación de los materiales fertilizantes de presentación granulada se realizó en forma localizada sobre el suelo de la zona de goteo de los brinzales, mientras que los micronutrimentos se aplicaron de manera foliar.
Variables evaluadas
Las variables evaluadas fueron: diámetro basal, incremento de diámetro basal, longitud y diámetro basal del brote principal, peso seco de 100 acículas y retención de follaje. El diámetro basal se midió en la base del tallo de los árboles con un vernier digital marca Truper, sobre una marca hecha con un marcador indeleble al momento de establecer el experimento. A partir de esta variable se estimó el incremento de diámetro basal. La longitud del brote principal se midió con un flexómetro graduado marcar Truper, desde la base del brote hasta la punta del mismo. El diámetro del brote principal se midió en la base, con un vernier digital Truper. El peso seco de 100 acículas se determinó separando 100 acículas provenientes de la parte media de las ramillas de cada muestra recolectadas el 29 de enero de 2018 conforme a lo indicado por Moilanen et al. (2010). Las muestras estuvieron compuestas de tres ramillas por árbol, de cada una de las cuales se obtuvo entre 33 y 34 acículas de su parte media, para completar 100 acículas por árbol o unidad experimental. De esta forma, se procesaron tres repeticiones por tratamiento. Los grupos de 100 acículas fueron secados en una estufa de circulación forzada marca FELISA, a 70 oC por 48 h.
Para la evaluación de la retención de follaje se tomaron tres muestras o ramas de tres unidades experimentales por tratamiento; una de la parte baja, otra de la parte media y una más de la porción alta de la copa del árbol. Las muestras se colocaron en una habitación ventilada durante un mes, para después recolectar el follaje retenido y no retenido para secarlo hasta tener un peso constante. Con los datos obtenidos se calculó la proporción de follaje retenido.
El estado nutrimental de los brinzales, en respuesta a los tratamientos, se evaluó con la técnica de análisis gráfico de vectores (Timmer y Stone, 1978), utilizando el dato de peso seco de 100 acículas, las que posteriormente fueron preparadas para su procesamiento en laboratorio.
Las muestras de 100 acículas se enviaron al laboratorio Salvador Alcalde Blanco del Colegio de Postgraduados, para determinar en extractos de digestión ácida, las concentraciones de N por el método micro-Kjeldahl, de P, K, Fe, Cu, Zn, Mn y B mediante espectrofotometría de plasma inductivamente acoplado (ICP-AES).
Los análisis de suelo se realizaron mediante un muestreo en forma de zig zag en el área experimental. Se tomaron nueve muestras simples de suelo a una profundidad de 30 cm. Para obtener las muestras simples de suelo, primero se limpió la superficie eliminando la capa herbácea y la materia orgánica presente sobre el suelo y después se cavó un hoyo de forma cuadrada de 30 cm por lado, con una profundidad de 30 cm. Posteriormente, se limpió el hoyo sacando toda la tierra caída al momento de cavar, para luego tomar la muestra simple de suelo desde la superficie hasta 30 cm de profundidad. De las nueve muestras simples que se tomaron, se hicieron grupos de tres muestras compuestas. En el laboratorio se determinó pH, Capacidad de Intercambio Catiónico (CIC), contenido de materia orgánica, N, P, K, Ca, Mg y micronutrimentos.
Análisis estadístico
Las variables diámetro basal, incremento de diámetro basal, longitud y diámetro del interverticilo principal y el peso seco de 100 acículas se analizaron mediante un Análisis de Covarianza (ANCOVA) y pruebas de comparación de medias de Tukey (á =0.05), empleando como covariable el diámetro inicial (diámetro basal 2015). De igual manera, con los datos de retención de follaje se realizó un Análisis de Varianza (ANOVA) y prueba de comparación de medias (Tukey, á=0.05). Las concentraciones foliares de nutrimentos analizados se interpretaron mediante la técnica de análisis de vectores (Timmer y Stone, 1978).
Resultados y discusión
Variables morfológicas
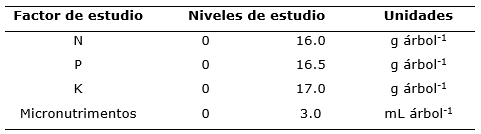
El Análisis de Covarianza (ANCOVA) mostró que las interacciones N*Mi, N*P*Mi (Tabla 2) son estadísticamente significativas. Por otro lado, la prueba de Tukey (á=0.05) indicó que no existen efectos significativos de la aplicación de N, P, K y Mi en el diámetro basal 2018 (Figura 2). Estos resultados muestran que no cualquier tratamiento de fertilización produce incrementos de diámetro de los árboles, aspecto que debe tenerse presente al momento de implementar un programa de fertilización en plantaciones forestales (Vásquez et al., 2015).
Tabla 2 - Análisis de covarianza para el diámetro basal 2018 en el experimento de fertilización de Pseudotsuga menziesii en el ejido "Palo Bendito", Huayacocotla, Veracruz
| Fuente | DF | Suma de cuadrados | Cuadrado de la media | F-Valor | Pr > F |
| Modelo | 16 | 4802.357298 | 300.147331 | 11.33 | <.0001 |
| N*Mi | 1 | 110.779245 | 110.779245 | 4.18 | 0.0451 |
| N*P*Mi | 1 | 139.550445 | 139.550445 | 5.27 | 0.0251 |
| DM15 | 1 | 4330.935263 | 4330.935263 | 163.42 | <.0001 |
| Error | 63 | 1669.642657 | 26.502264 | ||
| Total corregido | 79 | 6471.999955 |
N= nitrógeno, P=fósforo, Mi= micronutrimentos, DM15= diámetro basal 2015 (antes de aplicados los tratamientos)
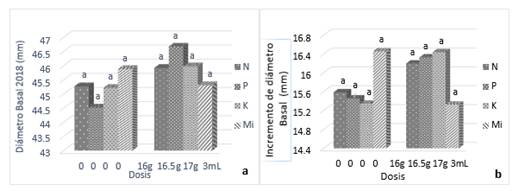
Fig. 2 - Efectos de los factores probados en el experimento (dosis aplicadas de N, P, K y micronutrimentos) de fertilización de árboles de Pseudotsuga menziesii sobre el diámetro basal alcanzado en 2018 y el incremento en diámetro basal
Para el caso del incremento de diámetro basal, el análisis de covarianza muestra que no hubo efectos significativos de los factores probados (Tabla 3); (Figura 2). Esto se puede deber a la alta variabilidad del diámetro basal inicial de los brinzales entre unidades experimentales, dado que la selección de las unidades experimentales se basó en la altura de los brinzales y no en el diámetro basal. El incremento de diámetro basal de brinzales solo fue influenciado de manera significativa por la interacción N*P*Mi (Tabla 3). En la figura #2b, se puede apreciar que no hay efectos significativos de los niveles de los nutrimentos probados, aunque las tendencias de N, P y K indican que la aplicación de cualquiera de ellos generó mayor incremento de diámetro basal que los correspondientes tratamientos sin fertilización. En base a los efectos de los fertilizantes aplicados (Figura 2), el N es levemente deficiente en el sitio de estudio; en cambio, el P y K son más deficientes, ya que con su aplicación los brinzales respondieron de manera más categórica en cuanto al incremento de diámetro basal, aunque sin diferencias estadísticamente significativas. La falta de efectos más pronunciados ante la aplicación de los nutrimentos se debe, posiblemente, a que las dosis fueron bajas con respecto a los niveles de demanda y condiciones de fertilidad de la plantación. En efecto, el sitio experimental parece ser de baja fertilidad dado que, en la región, la principal arcilla que conforma el suelo es la caolinita, material típicamente de baja capacidad de intercambio catiónico (Hernández, 2017). Por otro lado, los requerimientos nutrimentales en el tejido vegetal de Pseudotsuga menziesii, especialmente los relativos a K son elevados (0.80 %; (Weetman et al., 1992), si se comparan con los de Pinus patula, 0.63 %; (Sánchez, 2013), que es una de las coníferas naturales del sitio de estudio. Esto significa que, probablemente, las dosis aplicadas de K fueron insuficientes para promover mayores respuestas a la aplicación.
Tabla 3 - Análisis de covarianza para el incremento de diámetro basal en el experimento de fertilización de Pseudotsuga menziesii en el ejido "Palo Bendito", Huayacocotla, Veracruz
| Fuente | DF | Suma de cuadrados | Cuadrado de la media | F-Valor | Pr > F |
| Modelo | 16 | 556.706092 | 34.794131 | 1.31 | 0.2179 |
| N*P*Mi | 1 | 136.0593613 | 136.0593613 | 5.13 | 0.0269 |
| DM15 | 1 | 166.8861830 | 166.8861830 | 6.30 | 0.0147 |
| Error | 63 | 1669.642657 | 26.502264 | ||
| Total corregido | 79 | 2226.348749 |
N= nitrógeno, P=fósforo, Mi= micronutrimentos, DM15= diámetro basal 2015
Por otra parte, el ANCOVA realizado sobre los datos de la longitud de brote principal, con un valor p=0.267, indica que no existen diferencias estadísticamente significativas entre tratamientos (Tabla 4). La prueba de Tukey (á=0.05) indica que no existen diferencias significativas entre los niveles cero y uno de N, P, K y Mi (Figura 3); no obstante, a pesar de la gran variabilidad que normalmente presentan las variables relacionadas con la altura de árboles incluyendo la longitud de brote central, se observó una tendencia a incrementar la longitud del brote cuando se fertilizó con K (Figura 3). Esto puede significar que este nutrimento presente mayor deficiencia en el sitio de estudio, aunque Vázquez et al. (2018) encontraron que el K favoreció el crecimiento de los brinzales en Pinus greggii Engelm, aun cuando este nutrimento no fue deficiente en el sitio experimental. Los autores proponen que este efecto se debió, probablemente, a que K promueve la eficiencia en el uso del agua (Pereira Pérez et al., 2016).
Tabla 4 - Análisis de covarianza para longitud de brote principal en el experimento de fertilización de Pseudotsuga menziesii en el ejido "Palo Bendito", Huayacocotla, Veracruz
| Fuente | DF | Suma de cuadrados | Cuadrado de la media | F-Valor | Pr > F |
| Modelo | 16 | 2603.12187 | 162.69512 | 1.24 | 0.2670 |
| DM15 | 1 | 766.0564970 | 766.0564970 | 5.84 | 0.0188 |
| Error | 59 | 7743.60800 | 131.24759 | ||
| Total corregido | 75 | 10346.72987 |
DM15= diámetro basal 2015
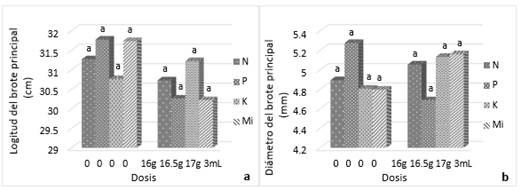
Fig. 3 - Pruebas de Tukey (á=0.05) para las variables longitud del brote principal (a) y diámetro del brote principal (b) en el experimento de fertilización de árboles de Pseudotsuga menziesii en ejido "Palo Bendito", Huayacocotla, Veracruz
Como en el caso de diámetro del brote principal, el ANCOVA mostró que no existen efectos significativos de los factores probados (p=0.2121) en el diámetro del brote principal (Tabla 5). Se encontraron solo efectos significativos en esta variable de la interacción P*K. La prueba de Tukey (á=0.05) confirma que no existen diferencias entre las dosis aplicadas de los nutrimentos (Figura 3). Por otra parte, si bien los efectos del K no fueron significativos, se observó que su aplicación incrementó el diámetro de brote respecto al tratamiento sin K. En coincidencia con el comportamiento de otras variables como diámetro basal e incremento de diámetro basal, existen incrementos del crecimiento, aunque estos no son significativos. Este tipo de tendencia puede indicar deficiencia de K en el sitio, de modo que su aplicación tiende a incrementar el diámetro del brote ((Figura 3), aunque no significativamente, debido a retraso en los efectos de fertilizante, causados por posible retención de los nutrimentos en el xilema de los brinzales, como lo explican López-López y Flores-Nieves (2016), o bien, debido a que la deficiencia del nutrimento en el sitio es demasiado aguda al tiempo que la demanda de nutrimento por el árbol es muy alta, como se sustentó anteriormente; todo esto, aunado a la elevada variabilidad de las unidades experimentales al inicio del experimento, según lo demuestra el efecto del DM15 (Tabla 5).
Tabla 5 - Análisis de covarianza para el diámetro del brote principal en el experimento de fertilización de Pseudotsuga menziesii en el ejido "Palo Bendito", Huayacocotla, Veracruz
| Fuente | DF | Suma de cuadrados | Cuadrado de la media | F-Valor | Pr > F |
| Modelo | 16 | 67.9788981 | 4.2486811 | 1.33 | 0.2121 |
| P*K | 1 | 12.96649066 | 12.96649066 | 4.05 | 0.0488 |
| DM15 | 1 | 15.29947337 | 15.29947337 | 4.78 | 0.0328 |
| Error | 59 | 188.9155966 | 3.2019593 | ||
| Total corregido | 75 | 256.8944947 |
P=fósforo, K=potasio, DM15= diámetro basal 2015
En cuanto al Peso Seco (PS) de 100 acículas, al utilizar el diámetro basal 2015 como covariable, se identifica que esta covariable no influyó en el comportamiento del peso seco de 100 acículas, de acuerdo con el ANCOVA (Tabla 6). Esto significa que esta variable no es afectada por el tamaño de los árboles, lo que la hace ser más confiable que otras para evaluar los efectos de fertilización en P. menziezii. En el caso de esta variable, los efectos significativos encontrados son ejercidos por el K, siendo destacables también las interacciones P*Mi; y P*K*Mi. La prueba de Tukey (á=0.05) confirma lo anterior (Figura 4). De acuerdo con este resultado, es posible afirmar que el K es un elemento deficiente en el área de estudio. Weetman (1989) menciona que los árboles con una buena nutrición tienden a crecer más rápidamente que los árboles con deficiencias nutrimentales y que, por lo regular, el crecimiento se presenta en altura y diámetro basal o a la altura del pecho, pero también se presenta un crecimiento en el tamaño de las acículas.
Tabla 6 - Análisis de covarianza para el Peso Seco (PS) de acículas en el experimento de fertilización de Pseudotsuga menziesii en el ejido "Palo Bendito", Huayacocotla, Veracruz
| Fuente | DF | Suma de cuadrados | Cuadrado de la media | F-Valor | Pr > F |
| Modelo | 16 | 0.29771736 | 0.01860734 | 2.17 | 0.0317 |
| K | 1 | 0.06293008 | 0.06293008 | 7.33 | 0.0109 |
| P*Mi | 1 | 0.06006675 | 0.06006675 | 7.00 | 0.0127 |
| P*K*Mi | 1 | 0.04048408 | 0.04048408 | 4.71 | 0.0377 |
| DM15 | 1 | 0.00894011 | 0.00894011 | 1.04 | 0.3155 |
| Error | 31 | 0.26618855 | 0.00858673 | ||
| Total corregido | 47 | 0.56390592 |
P=fósforo, K=potasio, Mi= micronutrimentos, DM15= diámetro basal 2015
Variables fisiológicas
Retención de follaje
El Análisis de Varianza (ANOVA), realizado para la retención de acículas, indica que hay efectos significativos entre los tratamientos probados (Tabla 7); (Figura 5). El tratamiento cuatro, que consistió en la aplicación de P y el tratamiento uno (tratamiento testigo sin fertilizante) fueron los tratamientos que menor proporción de follaje retuvieron. Se esperaba que el tratamiento testigo fuera uno de los que menor cantidad de follaje retuvieran debido a que no recibió ninguno de los nutrimentos probados en este estudio. Los resultados indican que el P, por sí solo, no ayuda a la retención de follaje, pero en combinación con el K, constituye el tratamiento que ayuda a retener mayor proporción de follaje. El N y el K, por sí solos, promovieron la retención de una mayor cantidad de follaje, ya que son estadísticamente iguales al tratamiento siete, el cual retuvo la mayor cantidad de follaje.
Tabla 7 - Análisis de varianza para la retención de acículas en el experimento de fertilización de Pseudotsuga menziesii en el ejido "Palo Bendito", Huayacocotla, Veracruz
| Fuente | DF | Suma de cuadrados | Cuadrado de la media | F-Valor | Pr > F |
| Modelo | 15 | 3796.876637 | 253.125109 | 5.37 | <.0001 |
| Error | 32 | 1508.839576 | 47.151237 | ||
| Total corregido | 47 | 5305.716213 |
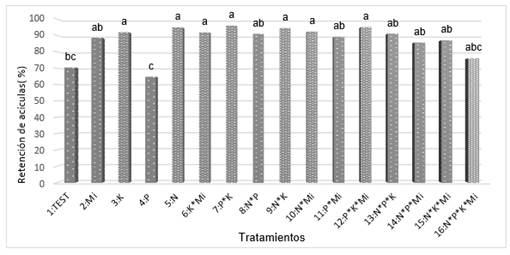
Fig. 5 - Retención de acículas en el experimento de fertilización de Pseudotsuga menziesii en el ejido "Palo Bendito", Huayacocotla, Veracruz
Este resultado es importante dado que, en el caso de árboles de navidad, la retención de follaje es una característica altamente deseable desde el punto de vista comercial (Chastagner y Riley, 2003) y el mercado prefiere que los árboles retengan el follaje por tres semanas o más. De acuerdo con esta variable, se recomienda aplicar en el sitio materiales fosfatados y potásicos para mejorar la retención de follaje. Este resultado es congruente con lo indicado por la mayoría de variables morfológicas, las cuales, aunque no mostraron efectos significativos (excepto el peso seco de acículas) de los tratamientos, sí indicaron posibles deficiencias de K y P en las tendencias de crecimiento.
Estado nutrimental
Todos los tratamientos generaron un mayor peso seco de acículas que el tratamiento testigo (Figura 6). Asimismo, casi todos los tratamientos, excepto el tratamiento nueve, mostraron concentraciones bajas de N, de acuerdo con las concentraciones críticas (1.45 %) establecidas para Pseudotsuga menziesii por Ballard y Carter (1986).

Fig. 6 - Diagrama de Timmer para el análisis de N en el experimento de fertilización de Pseudotsuga menziesii en el ejido "Palo Bendito", Huayacocotla, Veracruz
En los análisis de suelos, se puede observar claramente que la concentración de N es muy baja, comparada con el valor adecuado según la norma 021-RECNAT-2000 (SEMARNAT, 2000). Debido a que la concentración foliar de N en casi todos los tratamientos es inferior a la concentración crítica, es probable que exista una deficiencia de este nutrimento en el sitio de estudio, aun cuando la aplicación de este nutrimento no arrojó valores significativamente superiores en las variables morfológicas. (Tabla 8)
Tabla 8 - Análisis de suelos del sitio experimental, interpretados de acuerdo con la NOM-021-RECNAT-2000
| Parámetro | Muestra | Alto | Bajo | Diagnóstico |
| CE(dS/m) | 0.39 | 2 | 1.1 | Bajo |
| pH | 4.10 | 6.5 | 5.1 | Bajo |
| CIC(meq/100 g) | 22.19 | 25 | 15 | Adecuado |
| N(%) | 0.08 | 0.8 | 0.3 | Bajo |
| P Bray (ppm) | 0.53 | 30 | 15 | Bajo |
| K (meq/100 g) | 0.001 | 0.6 | 0.3 | Bajo |
| Ca(meq/100 g) | 3.00 | 10 | 5 | Bajo |
| Mg(meq/100 g) | 1.50 | 3 | 1.3 | Adecuado |
| Fe(ppm) | 13.26 | 4.5 | 4.5 | Elevado |
| Cu(ppm) | 0.11 | 0.2 | 0.2 | Bajo |
| Zn(ppm) | 0.18 | 1 | 1 | Bajo |
| Mn(ppm) | 3.95 | 1 | 1 | Elevado |
| B(ppm) | 0.00 | 1 | 1 | Bajo |
CE=conductividad eléctrica, pH=potencial hidrógeno, CIC=capacidad de intercambio catiónico, N=nitrógeno, P=fósforo, K=potasio, Ca=calcio, Mg=magnesio, Fe=hierro, Cu=cobre, Zn=zinc, Mn=manganeso, B=boro
La falta de respuestas a la aplicación de N se debió, de acuerdo con la figura #6, a que las deficiencias de P y K, en el sitio experimental, son probablemente más agudas que las de N. En efecto, los tratamientos con N, ya sea solo o combinado con P (N*P) o K (N*K), generaron las mayores concentraciones de N, lo que indica que internamente el N es excesivo con respecto a los otros nutrimentos (López y Alvarado, 2010). La figura 6 muestra también que los cuatro tratamientos que generaron las mayores biomasas de acículas incluyeron la aplicación de K, mientras que los dos más productivos incluyeron P, lo que confirma que estos nutrimentos (K y P) son los más deficientes en el área experimental.
Con respecto al fósforo (Figura 7), todas las concentraciones foliares de los tratamientos aplicados fueron bajas en comparación con las concentraciones críticas (0.15 %) para esta especie (Ballard y Carter, 1986). Esto concuerda con lo indicado por los análisis de suelos (Tabla 8), lo que muestran que existe deficiencia de P, de acuerdo con la norma 021-RECNAT-2000 (SEMARNAT, 2000).

Fig. 7 - Diagrama de Timmer para el análisis de P en el experimento de fertilización de Pseudotsuga menziesii en el ejido "Palo Bendito", Huayacocotla, Veracruz
De acuerdo con la figura #7, el hecho de que el tratamiento con suministro solo de P haya generado mayor biomasa foliar y concentraciones más altas de este nutrimento en los tejidos, en comparación con el tratamiento testigo, indica que el nutrimento es deficiente en el sitio, pero la dosis utilizada de P fue suficiente para eliminar la deficiencia, mientras que otro u otros factores limitaron el crecimiento (López y Alvarado, 2010). En este caso, es posible que ese factor limitante haya sido el K. La deficiencia de P se confirma porque de los siete tratamientos que generaron mayor peso seco de acículas, cinco de ellos recibieron alguna dosis de P (Figura 7), lo que de acuerdo con López y Alvarado (2010) corresponde a vectores de (Timmer y Stone, 1978), del tipo +++ o bien, +-+ (López y Alvarado, 2010), los que indican deficiencia del nutrimento antes de su aplicación.

Fig. 8 - Diagrama de Timmer para el análisis de K en el experimento de fertilización de Pseudotsuga menziesii en el ejido "Palo Bendito", Huayacocotla, Veracruz
Para el caso del K, todos los tratamientos que recibieron alguna dosis de este nutrimento, excepto cuando se suministró como nutrimento e único, generaron concentraciones más bajas que la concentración crítica de K (0.80 %) de esta especie (Ballard y Carter, 1986); (Van den Driessche, 1989). En la mayoría de los tratamientos se generó dilución del K foliar por crecimiento [vectores tipo +-+, o bien, +-; López y Alvarado (2010) ], pero se identifica claramente que la aplicación de N limitó la absorción (menores contenidos) de K (tratamientos 5, 8, 9 y 10). Lo anterior confirma que se trata de un nutrimento deficiente. (Figura 8) La concentración foliar alta de K en el tratamiento que solo recibió K se debió a que no recibió N, pues es bien conocida la relación de antagonismo entre estos dos nutrimentos en el suelo (Fageria, 2001). De acuerdo con Van den Driessche (1991), este vector corresponde al vector de Timmer del tipo +++ (Park et al., 2015), el cual indica que este nutrimento es muy poco abundante en el área de estudio (Ballard y Carter, 1986). Por otro lado, de los cuatro tratamientos que generaron mayor biomasa de acículas, tres recibieron K, lo que nuevamente indica posible deficiencia de K en el sitio. Esta deficiencia se confirma, además, con la prueba de Tukey (á=0.05) para el peso seco de 100 acículas (Figura 4), que muestra los efectos positivos del potasio. Los análisis de suelo indicaron que el K es un nutrimento deficiente en el área de estudio de acuerdo con la norma 021-RECNAT-2000. Cabe mencionar que en una de las tres muestras de suelo enviadas al laboratorio no fue detectado el K, y en los dos restantes la concentración determinada fue muy baja (Tabla 8).

Fig. 9 - Diagrama de Timmer para el análisis de micronutrimentos en el experimento de fertilización de Pseudotsuga menziesii en el ejido "Palo Bendito", Huayacocotla, Veracruz
En los casos de Fe y B todos los tratamientos tienen mayores concentraciones que los respectivos tratamientos testigos (Figura 9), aunque los micronutrimentos no ocasionaron un crecimiento significativo de las acículas. De acuerdo con las concentraciones críticas de Fe (>50 ppm) y B (>20 ppm), determinadas por Ballard y Carter (1986) y Van den Driessche (1989), estos nutrimentos no son deficientes en el área de estudio. De acuerdo con los análisis de suelo, efectivamente, el Fe no es deficiente en el sitio; sin embargo, el B sí presenta deficiencia (Tabla 8). En el caso del Zn todos los tratamientos (excepto uno) generaron una mayor concentración que el tratamiento testigo. Las concentraciones críticas de Zn (15 ppm), establecidas por Ballard y Carter (1986) y Van den Driessche (1989), indican que este nutrimento posiblemente es no es eficiente, ya que las concentraciones de Zn en las plantas de cinco de los 16 tratamientos son menores a 15 ppm. De hecho, los análisis de suelo indican que el Zn es deficiente en el sitio experimental. El Cu posiblemente también lo es en las plantas, de acuerdo con las concentraciones críticas Cu>3ppm; Ballard y Carter, (1986), ya que todos los tratamientos tienen menos de tres partes por millón, mientras que los análisis de suelo también indican deficiencia de este nutrimento.
De acuerdo con los resultados obtenidos en la presente investigación, se puede sugerir la aplicación de dosis iguales o mayores que las proporcionadas de P y K, ya que estos nutrimentos son deficientes en el sitio. También se sugiere aplicar una dosis media de N, pues su deficiencia es menos aguda que las de P y K. Se recomienda una dosis de micronutrimentos igual o mayor que la utilizada en este estudio, debido a que, aunque no tuvieron efecto positivo en el crecimiento, sí propiciaron un aumento de sus concentraciones foliares, por lo que al remediarse las deficiencias de macronutrimentos, se evitará el desarrollo de deficiencias de estos elementos menores.
El P es un nutrimento deficiente en el sitio experimental y tiende a aumentar el diámetro basal de los brinzales de Pseudotsuga menziesii.
El K es el nutrimento más deficiente en el sitio y su aplicación a los árboles genera incrementos significativos de algunas variables morfológicas.
El N es un nutrimento ligeramente deficiente en relación con los demás macronutrimentos en el área de estudio.
Los análisis de suelo en laboratorio son probablemente más eficientes que los ensayos de fertilización para detectar deficiencias de N en la especie estudiada.
La aplicación de micronutrimentos en el área no afecta positivamente las variables morfológicas de los árboles de P. menziesii; sin embargo, aumenta las concentraciones foliares de algunos de ellos, lo que puede prevenir deficiencias de micronutrimentos después de la enmienda de las deficiencias de macronutrimentos.
La aplicación de una mezcla de N, P y K promueve mayor retención de acículas en los árboles, en el área experimental.
Las tasas de crecimiento de los árboles responden positivamente a la fertilización química, siempre y cuando se apliquen los nutrimentos deficientes en el sitio.
La retención de follaje postcosecha de brinzales de Pseudotsuga menziesii se puede mejorar significativamente mediante el manejo nutrimental de las plantaciones.

