










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkCultivos Tropicales
versión On-line ISSN 1819-4087
cultrop vol.39 no.3 La Habana jul.-set. 2018
ARTÍCULO ORIGINAL
Efectos de la temperatua en el metabolismo de proteínas y carbohidrratos y el índice de vegetación en trigo (Triticum durum L.)
Temperature effects on protein and carbohydrates metabolism and the vegetation index in wheat (Triticum durum L.)
Leandris Argentel-Martínez,I-II Jaime Garatuza-Payán,II Enrico A. Yépez,II Francisco J. Salazar-Huerta,I Tulio ArredondoIII
I Instituto Tecnológico del Valle del Yaqui, calle 600, Block 611, Bácum, 85275 San Ignacio Río Muerto, Sonora, México
II Instituto Tecnológico de Sonora. Dirección de Recursos Naturales, 5 de febrero, 818 Sur, Col. Centro, Cd. Obregón, Sonora. México. CP: 85000
III Instituto Potosino de investigación Científica y Tecnológica. San Luis Potosí, SLP, México. CP: 78216
RESUMEN
Se estableció un experimento en condiciones de campo en el Valle del Yaqui, Sonora, México, que consistió en elevar la temperatura del trigo en 2 °C por encima de la temperatura ambiente del dosel durante la fenología del cultivo y determinar su efecto en los contenidos de proteínas (PST) y carbohidratos solubles totales (CTS) en tallos y en grano, la tasa de movilización de los carbohidratos (TMC) del tallo, y el índice de vegetación de diferencia normalizada (NDVI). Se utilizó como modelo experimental la variedad de trigo cristalino CIRNO C2008. El contenido de PST, por efecto del calor disminuyó significativamente en la fenofase de espigamiento respecto al tratamiento de Control, lo que denota una senescencia temprana. El contenido CST en el tallo fue mayor en el tratamiento de Calor cuando se determinó 10 días después de la floración. En la maduración no existieron diferencias significativas. El calor impuesto no afectó la TMC. En el grano, el calor no afectó el contenido de PST, pero sí se incrementó el contenido de CST. El NDVI, debido al calor, disminuyó a partir de la fenofase de elongación del tallo hasta la maduración. El máximo valor de NDVI ocurrió, en los tratamientos en la floración, demostrando tolerancia de la variedad al calor para la ocurrencia de esta fenofase. Tales resultados sugieren la factibilidad del uso de CIRNO C2008, como posible progenitor para programas de mejora genética ante el estrés térmico.
Palabras clave: calor, fenología, cambio climático.
ABSTRACT
An experiment, under field conditions, in the Valle del Yaqui, Sonora, Mexico, was established which consisted of raising the temperature of the wheat at 2 °C above the ambient canopy temperature during the crop phenology to determinate warming effect on total soluble proteins (PST) and carbohydrates (CST) in stems and grain and the total soluble carbohydrates mobilization rate within the stem (TMC), and the NDVI. CIRNO C2008, a crystalline wheat variety was used as an experimental model. The PST, due to warming effect, decreased significantly in the heading phenophase in more than half of the Control treatment, aspect that denotes an early senescence. However, the CST content in the stem was higher in the heat treatment when it was determined 10 days after flowering. In the maturity there were no significant differences. The imposed heat did not affect the TMC. In the grain, heat did not affect the PST content but the CST content increased. The NDVI, due to heat effect, decreased from the stem elongation to maturity. The maximum value occurred, in the treatments, in flowering, demonstrating warming tolerance of the used variety to the occurrence of this phenophase. Such results suggest the feasibility of using CIRNO C2008 as a possible progenitor for breading in front heat stress.
Key words: heat, phenology, climate change.
INTRODUCCIÓN
Los escenarios de cambio climático pronosticados para los últimos años, según informes del IPCC, constituyen una amenaza potencial para el desarrollo de la agricultura en diversas regiones del mundo (1).
México, por su situación geográfica, posee gran diversidad de climas y ecosistemas (2); sin embargo, los climas áridos y semiáridos son los que mayormente predominan, cubriendo casi el 50 % de su extensión territorial total (3). En tales condiciones climáticas el desarrollo de la vegetación, la actividad microbiana del suelo y el ecosistema se dificulta por la incidencia negativa de factores abióticos, donde tiene marcada incidencia el incremento de la temperatura (4).
La actual vulnerabilidad de la agricultura en estas regiones semiáridas de México, podría exacerbarse por los efectos que se esperan de los escenarios de cambio climático, que incluyen principalmente la disminución de la cantidad de horas frío de muchos cultivos y la amplia y significativa variabilidad de las temperaturas, lo cual probablemente afectará el desempeño fisiológico de los cultivos, cuya relación temperatura del dosel y el rendimiento es muy estrecha (5).
El valle del Yaqui en el estado de Sonora, es una de las regiones de mayor contribución a la producción nacional de trigo en México (6). Esta región será impactada por escenarios del cambio climático, donde se pronostican incrementos de temperatura de hasta 2 °C para los próximos 10 años (7). Existen algunos reportes, basados en modelos predictivos parametrizados que concluyen que el trigo será uno de los cultivos de mayor grado de afectaciones. Sin embargo, a nivel de planta, teniendo en cuenta la interacción genotipo-ambiente, la variabilidad del desempeño fisiológico, bioquímico y agronómico de esta especie, en esta región, no está del todo descrita. Su estudio detallado, en condiciones de campo, contribuirá al entendimiento de mecanismos fisiológicos y bioquímicos durante su adaptación al cambio climático (8).
La morfología de las plantas es el reflejo de complejos procesos fisiológicos y bioquímicos que tienen lugar a nivel de órganos y tejidos, los cuales varían en función de las condiciones edafoclimáticas existentes en los ecosistemas de cultivo (9). Leves variaciones de las condiciones climáticas pueden ocasionar significativa reducción de la tasa de desarrollo de sus órganos, principalmente, el área foliar y el rendimiento en las plantas (10). El estrés térmico pudiera entonces afectar el contenido de clorofilas y modificar el metabolismo glucídico y proteico de las plantas, así como la calidad del grano (11). La superficie foliar de la planta refleja intensamente la energía en el rango del infrarrojo cercano (12). La reflectancia es determinada por las propiedades de los tejidos foliares; es decir, por su estructura celular y las interfaces entre la pared celular exterior, el protoplasto y el cloroplasto, lo cual es indicativo de su estado fisiológico (9).
Estas características anatómicas de las plantas una vez afectadas por factores estresantes, como el calor, pudiera ocasionar variaciones en los valores del índice de vegetación de diferencia normalizada (13), por lo cual su evaluación in situ ofrece información confiable del estado nutricional de la planta (14), que se puede corroborar con las variaciones en los contenidos de proteínas solubles totales, como evidencia de la nutrición mineral nitrogenada durante el desarrollo y en el grano y el contenido de carbohidratos solubles totales en el tallo y en el grano, como evidencia de la eficiencia de la actividad fotosintética ante el posible efecto adverso del calor. En tal sentido se realizó la presente investigación con el objetivo de evaluar el efecto del calentamiento a base de 2 °C por encima de la temperatura ambiente del dosel del cultivo, en los contenidos de proteínas (PST) y carbohidratos solubles totales (CTS) en tallos y en grano, la tasa de movilización de los carbohidratos (TMC) del tallo, y el índice de vegetación de diferencia normalizada (NDVI).
MATERIALES Y MÉTODOS
Ubicación de área experimental
El experimento se desarrolló durante la campaña de cultivo de diciembre 2016 hasta abril 2017, en condiciones de campo, en el Centro Experimental de Transferencia de Tecnología (CETT-910) del Instituto Tecnológico de Sonora (ITSON), ubicado en el Valle del Yaqui a los 27° 22’0.4’’ N y 109° 54’50.6’’ W (UTM: 607393.24 m E; 3027508.34 m N).
Tratamientos y control de temperatura
Se establecieron dos variantes experimentales, T1: incremento de 2 °C a la temperatura ambiente del dosel del cultivo en las parcelas y T2: temperatura ambiente del dosel en las parcelas de control (tratamiento de control). Tales tratamientos fueron distribuidos siguiendo un diseño experimental completamente aleatorizado con 12 repeticiones.
Para elevar la temperatura del dosel del cultivo, a partir de los 15 días posteriores a la germinación, se utilizaron seis radiadores térmicos por parcela (modelo FTE-1000, 1000 W, 240 V, 245 mm de largo x 60 mm de ancho, construidos por Mor Electric Company Heating Association Inc. Comstock Park, MI, EE. UU.), los que fueron ubicados sobre estructuras triangulares equiláteras de 5,2 m de lado (Figura 1).
Se instalaron dos radiadores en cada lado de las estructuras triangulares formando un hexágono regular, que emitían calor hasta elevar la temperatura 2 ºC por encima de la temperatura ambiente del dosel.
La superficie de cálculo de cada parcela fue un círculo de 3 m de diámetro circunscrito en el hexágono regular formado por los seis radiadores (1). Para el control térmico, se instalaron sensores de temperatura infrarrojo (ITSR Apogee Instruments Inc., Logan, UT, EE. UU.) dirigidos al centro de cada superficie de cálculo, con un ángulo de inclinación de 45° desde la superficie horizontal del suelo. Los sensores ITSR se acoplaron a un registrador de datos (CR1000 Campbell Sci, Inc. Logan, UT, EE. UU.) que envía una señal de voltaje a una interfaz (MAI-05V, Instrumentos Avatar) que, a su vez, traduce la señal de voltaje a miliamperios y la envía a un regulador (Atenuador A1P-24-30-S05, Instrumentos Avatar). Este regulador controla la corriente enviada a los calentadores, de modo que la cantidad de calor emitido aumenta o disminuye en función de la diferencia de temperatura entre las parcelas de Calor y de Control, a través de la rutina proporcional, integrativa y derivativa descrita (15).
Variables agroclimáticas
Durante el ciclo biológico del cultivo, la temperatura media mensual se mantuvo entre 17 y 24 °C (promedio 18,6 ºC). La precipitación mensual fue menor de 15,0 mm y la humedad relativa varió de 50 a 63 % (Figura 2).

Variedad utilizada
La variedad de trigo CIRNO C2008, clasifica como trigo cristalino o duro (Triticum durum L.). Se originó a partir de la selección en poblaciones segregantes de mestizaje SOOTY-9 / RASCON-37 // CAMAYO, llevado a cabo en el Centro Internacional para la Mejora del Maíz y el Trigo (CIMMYT).
Esta variedad fue liberada para el cultivo desde 2008, siendo ampliamente utilizada en México y particularmente en Sonora. El rendimiento de grano cuando la variedad fue liberada en Sonora y alcanzó, 5,6 t ha-1 y 6,3 t ha-1 con dos y tres riegos, respectivamente (16).
Siembra, fertilización y riegos
La siembra se realizó de manera mecanizada con una sembradora (SUB-24) el 8 de diciembre de 2016, en un suelo vertisol (17), a tres hileras en los surcos y una densidad de siembra de 170 kg ha-1. La fertilización de fondo se realizó a base de 250 kg ha-1 de urea + 100 kg ha-1 de fosfato monoamónico (MAP), 11-52-00, durante el primer riego (fenofase de desarrollo).
Se aplicó fertilizante nitrogenado a una dosis de 50 kg ha-1 de urea en cada riego de auxilio aplicado. En los tres riegos de auxilio se aplicaron con una lámina de agua promedio de 14 cm por cada riego y con un intervalo de riego de 25 días.
Control de plagas, enfermedades y malezas
Durante el desarrollo del cultivo se encontró presencia ligera de áfido foliar (Schizaphis graminum) y se aplicó el plaguicida Muralla Max (ia. Imidacloprid + Betaciflutrin) a razón de 0,20 L ha-1 en la periferia de las parcelas a una distancia de 3 m de la superficie de cálculo. Asimismo, se observó una ligera presencia de malas hierbas de hoja ancha y se controlaron manualmente antes de los riegos.
Variables evaluadas
Contenido de proteínas solubles totales en las fenofases de macollamiento y floración
Una muestra de 1,0 g de material fresco foliar se depositó en un mortero y se maceró. La extracción se realizó con 40,0 mL de una solución buffer de fosfato a pH 6,86 preparado a partir de fosfato de potasio, solución que se adicionó lentamente mientras se efectuaba la extracción (18). El contenido de proteínas solubles totales fue cuantificado mediante fotometría de absorción en un espectrofotómetro Hewlett Packard 8452, a una longitud de onda de 750 nm.
El contenido de carbohidratos, se determinó en base a masa seca de los dos últimos entrenudos superiores del tallo principal (Ent2 y Ent3), de plantas de cada tratamiento cosechadas 10 días después de iniciada la fenofase de floración (10 DDF) y en la fenofase de la maduración (previo a la cosecha). Cada uno de los entrenudos se dividió en segmentos de 1-2 mm con un bisturí. Se procesaron tres repeticiones por entrenudo, las cuales se promediaron para obtener una muestra final por cada tratamiento. Para este ensayo se tomaron tres plantas por repetición de cada tratamiento.
Para la extracción de los carbohidratos (19) se realizaron tres extracciones con 5 mL de etanol (80 %) a 0,05 g de masa seca de los segmentos tallo anteriormente descritos (10 DDF y en la maduración). El contenido de carbohidratos solubles totales (mg g MS-1) (MS: materia seca), se determinó por el método de la antrona (20). La absorbancia se midió a una longitud de onda de 625 nm en un espectrofotómetro (JENWAY 6405 UV/Vis) del Instituto Tecnológico de Sonora.
Después de cosechadas las muestras y evaluado su rendimiento de biomasa y de grano, se tomó una muestra aleatoria formada por 100 granos de cada repetición en ambos tratamientos, la que fue molida hasta obtener una harina y se determinaron nuevamente los contenidos de proteínas y carbohidratos solubles totales en el grano siguiendo las mismas metodologías anteriormente descritas.
La tasa de movilización de los carbohidratos totales en el tallo (TMC) (%), a los 10 DDF y la maduración, se calculó siguiendo la siguiente fórmula (21):
TMC= [(A-B) (A)-1] * 100
donde: A y B representa la concentración de carbohidratos totales (mg g MS-1) a los 10 DDF y en la maduración, respectivamente.
Índice de vegetación de diferencia normalizada (NDVI)
El índice de vegetación de diferencia normalizada se midió con un sensor portátil (Green Seeker, marca Trimble, desde 15 días después de la germinación, en cada fenofase hasta la cosecha para un total de 11 mediciones. Para esta medición se introdujo una variante experimental sin aplicar fertilizante nitrogenado conforme a las especificaciones del sensor. Se tomaron 10 mediciones en cada repetición de tratamiento, a una altura de 0,60 m del dosel del cultivo, conforme a la referencia del sensor. La escala de evaluación utilizada fue la recomendada por el fabricante: -1<NDVI>1, mayores valores del NDVI representan un mejor estado nutritivo (22).
Análisis estadísticos
Se comprobó el cumplimiento de los supuestos teóricos de homogeneidad de varianza (23) y se determinaron las medias, sus respectivas desviaciones y el error estándar. Los contenidos de proteínas y carbohidratos solubles totales y la removilización de carbohidratos en un total de seis repeticiones por tratamiento, así como, los contenidos determinados en el grano se compararon mediante una distribución teórica de probabilidades para variables cuantitativas continuas de t-Student para un nivel de significación del 1 %.
Para el procesamiento de los datos de NDVI obtenidos en cada fenofase, se compararon mediante un análisis de varianza de clasificación simple basado en un modelo lineal de efectos fijos (24) y cuando existieron diferencias entre las medias estas fueron comparadas por la prueba de comparación múltiple de Tukey para niveles de significación del 1 % (25). Se utilizó para estos análisis el paquete estadístico profesional ESTATISTICA, versión 8.4 para WINDOWS (26).
RESULTADOS Y DISCUSIÓN
Proteínas solubles totales en las fenofases de macollamiento y espigamiento
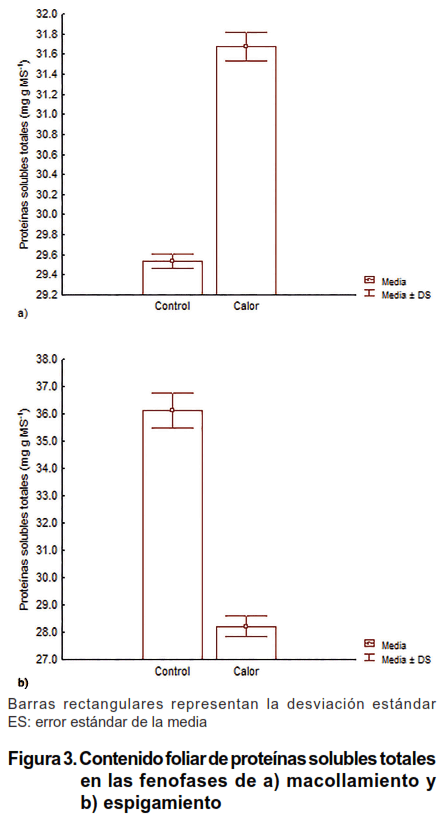
El contenido de proteínas solubles totales se incrementó significativamente en el tratamiento de Control en la fenofase de espigamiento; sin embargo, en el tratamiento de Calor se observó una respuesta contraria y su disminución en la fenofase de espigamiento, con respecto a la de macollamiento fue del 11,4 % de concentración existente en la fenofase de macollamiento (Figura 3).
El incremento de proteínas solubles totales encontrado en el tratamiento de Control, en la fenofase de espigamiento, indicó la capacidad de la variedad para acumular reservas metabólicas para la etapa reproductiva, donde el consumo energético y el dinamismo celular son elevados (26). Por su parte, lo observado en el tratamiento de Calor denotó su efecto adverso en la acumulación de proteínas solubles totales, viéndose favorecido quizás el catabolismo proteico. En la fenofase de espigamiento la integración metabólica entre glúcidos y proteínas es alta para asegurar fundamentalmente la calidad del polen (27). Por lo tanto, las disminuciones de los valores del indicador en el tratamiento de Calor fueron debido a la hidrólisis acelerada de las proteínas hasta sus precursores para elevar la concentración de osmolitos libres en el citosol y así disminuir su potencial osmótico e hídrico (28).
En condiciones de estrés abiótico, fundamentalmente por salinidad y sequía, en muchas especies vegetales, primeramente, se produce un incremento del contenido de proteínas solubles totales (29), pero estos valores van descendiendo, en dependencia de la intensidad del estrés y del grado de tolerancia de la variedad (30).
Carbohidratos solubles totales en las fenofases floración y en la maduración
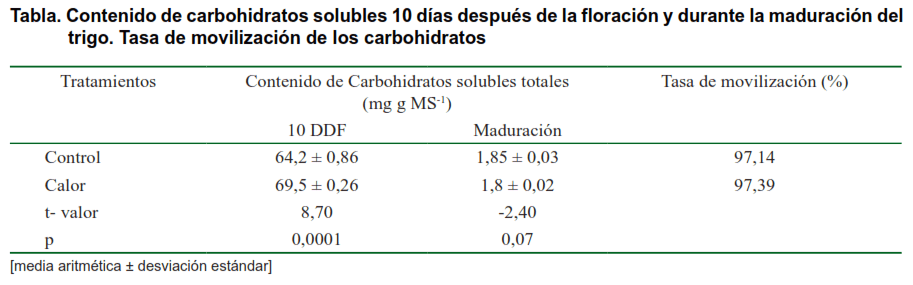
El contenido de carbohidratos totales en el tallo de las plantas fue mayor en el tratamiento de Calor cuando se determinó 10 DDF; sin embargo, en la fenofase de maduración no existieron diferencias significativas, pero si se encontró una reducción de su contenido entre las fenofases en que se evaluó (Tabla). El resultado encontrado sugiere un efecto positivo del calor impuesto sobre la acumulación de carbohidratos en la variedad usada como modelo experimental. Por otra parte, no existieron diferencias en la tasa de movilización demostrando que el calor no ofreció ningún impedimento bioquímico para este proceso.
Existen reportes sobre el efecto adverso del calor en la movilización de carbohidratos en otros cereales (31) y en trigo (32). El presente estudio constituye el primer reporte en el Valle del Yaqui sobre la estabilidad bioquímica del metabolismo de carbohidratos ante el incremento de la temperatura del dosel del cultivo en trigo.
Contenido de proteínas solubles totales en el grano
El contenido de proteínas solubles totales en el grano no presentó variación significativa por efecto del calor impuesto, aunque, se observó una tendencia a la disminución de sus valores en el tratamiento de Calor (Figura 4). Tal resultado demostró la posibilidad de que la variedad CIRNO C2008, transporte fuentes proteicas y precursores de proteínas hasta el fruto en condiciones de estrés.

Tal resultado tuvo lugar debido a un incremento de la actividad de las enzimas nitrato reductasa y glutamina sintasa, favoreciendo la acumulación y metabolismo de nitrógeno en el tratamiento de Calor (33). La concentración proteica en el grano de trigo (Triticum estivum L.) es una de las variables de mayor importancia para el rendimiento agrícola e industrial, debido a que es un elemento determinante del precio del grano debido a su valor nutricional (34).
La proteína en el grano de trigo es producto de la acumulación del nitrógeno absorbido por la planta durante su etapa vegetativa hasta el llenado del grano (35), y el porcentaje final, es una relación directa del nitrógeno disponible por la planta y el rendimiento del cultivo (relación proteína/carbohidratos) (36). Durante las primeras fases del llenado, ocurre el momento de mayor acumulación de proteínas en el grano y la velocidad de ésta dependerá del genotipo, de la disponibilidad simplástica de nitrógeno y de la temperatura ambiente (37).
Contenido de carbohidratos solubles totales en el grano
El mayor contenido de carbohidratos solubles totales se encontró en el tratamiento de Calor y fue significativamente superior al tratamiento de Control (Figura 5). Este resultado fue debido un incremento de actividad fotosintética del cultivo. La capacidad de algunas especies y variedades de plantas de acumular en condiciones adversas significativas cantidades de carbohidratos solubles en el grano es un mecanismo de protección del gluten para la posterior germinación (38).
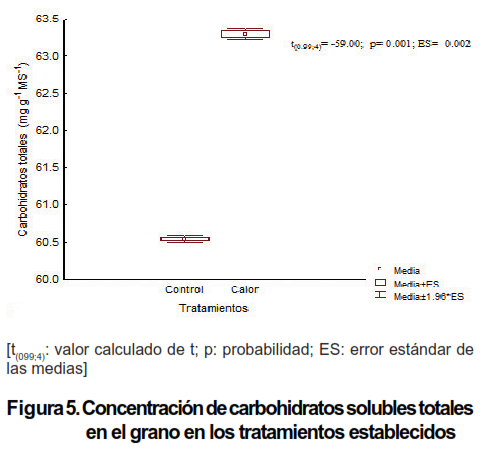
Un aspecto importante en la evaluación de la calidad del grano es la concentración de carbohidratos solubles, dado su efecto durante el procesamiento para la confección de pastas y harina (35). Las cosechas que han sido afectadas por el estrés abiótico, fundamentalmente el hídrico, merman su rendimiento y disminuyen la calidad del grano (39,40). En dicha reducción del rendimiento la disminución del contenido de carbohidratos en el grano ha sido más significativa que la movilización de proteínas solubles totales (41,42). Sin embargo, en el presente estudio, aunque se trata de estrés térmico, no se afectó la calidad y sí se incrementó el contenido de carbohidratos.
NDVI
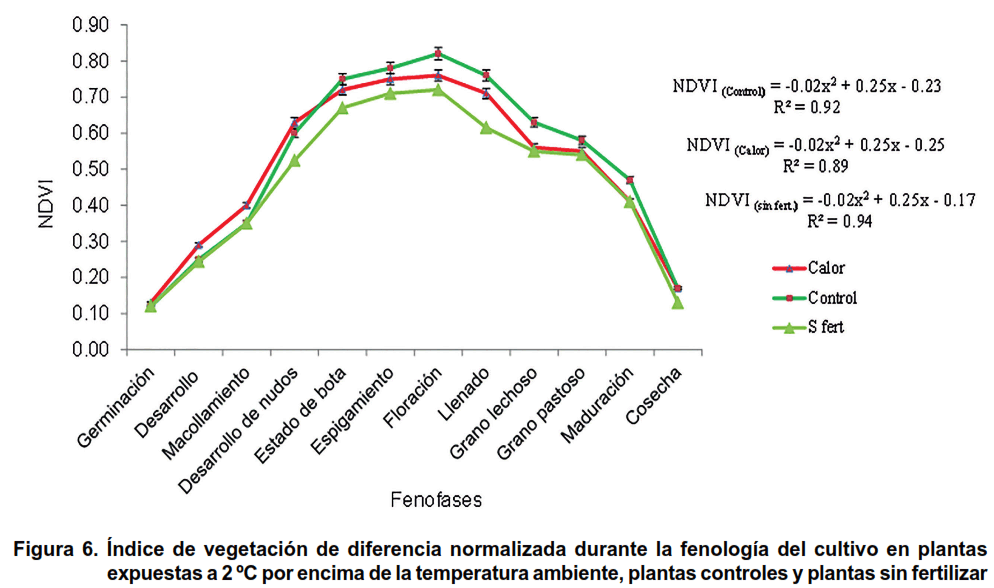
El NDVI en el tratamiento de Calor fue significativamente superior respecto al Control y a la variante sin fertilizante, desde la germinación hasta el macollamiento. Sin embargo, a partir de esta fenofase y hasta el desarrollo de los nudos, el indicador no evidenció diferencias significativas entre tratamientos. A partir de esta fenofase el NDVI en tratamiento Control superó estadísticamente al tratamiento de Calor hasta el final de la fenología del cultivo (Figura 6).
El valor máximo de la curva polinómica descrita por los valores de NDVI (parábola convexa de coeficiente cuadrático negativo) ocurrió en ambos tratamientos en la fenofase de floración. Este resultado demuestra la estabilidad genética de la variedad evaluada para la ocurrencia de esta fenofase (43), incluso ante condiciones de deficiencia de fertilizante nitrogenado y ante un posible estrés por calor. El hecho de haberse obtenido en esta fenofase el mayor valor de NDVI en el tratamiento de Control, evidenció el buen estado fisiológico y nutritivo de la planta y su condición favorable para el inicio de la etapa reproductiva.
El estudio del NDVI, que ha sido correlacionado con el contenido de nitrógeno y la clorofila foliar en el cultivo de maíz (Zea mays) (44) y el trigo (45), donde se evidenció que, aun cuando se produjo una disminución de la reflectancia espectral del calor, la variedad utilizada como modelo experimental alcanzó sus máximos valores en las fenofases de espigamiento, floración y llenado del grano en los tratamientos, donde se encontraron altos valores contenido de proteínas y carbohidratos solubles totales.
El NDVI ha sido propuesto como un indicador que predictivo de la actividad fotosintética de la planta, ya que esta es determinada principalmente por el contenido y la actividad de las clorofilas, lo que contribuye a la nutrición carbonada y conduce a la obtención de un alto contenido de carbohidratos en el grano (46).
Los análisis bioquímicos desarrollados en la variedad de trigo CIRNO C2008 sometida al incremento de la temperatura del dosel del cultivo en 2 °C a partir de los 15 días posteriores a la germinación, en condiciones de campo, demostraron que el calor no ejerce afectaciones significativas sobre la acumulación de carbohidratos.
Este resultado indicó afectaciones menores en el aparato fotosintético y en el metabolismo nitrogenado, aunque en el grano se observó una tendencia a disminuir el contenido de proteínas solubles totales. Tal resultado en el grano; sin embargo, sugiere una posible disminución de la calidad de la semilla y la calidad industrial para la confección de harina y pastas por efecto del incremento de la temperatura durante la fenología del cultivo.
CONCLUSIONES
- El contenido de PST, por efecto del calor disminuye en la fenofase de espigamiento en el tallo y no se afecta en los granos.
- El calor impuesto incrementa el contenido de CST en el tallo a los 10 días después de la floración y durante la maduración disminuye como evidencia de la ocurrencia de su movilización hacia los granos, a la vez que incrementa el contenido de CST en los granos.
- El NDVI, por efecto del calor, disminuye a partir de la fenofase de desarrollo de los nudos hasta la maduración. El máximo valor ocurre en esta variedad de trigo, en los tratamientos establecidos, durante la floración y los valores describen una función cuadrática convexa que solo difiere en el vértice.
BIBLIOGRAFÍA
1. Garatuza J, Argentel L, Yepez EA, Arredondo T. Initial response of phenology and yield components of wheat (Triticum durum L., CIRNO C2008) under experimental warming field conditions in the Yaqui Valley. PeerJ. 2018;6:e5064. doi:10.7717/peerj.5064
2. Moosavi SS, Abad HHS, Mohamadi GN, Imani AA. Evaluation of yield gap in various cultivars of wheat under climatic conditions of Ardabil region, Iran. Crop Research. 2017;52(1–3):14–20.
3. Millar N, Urrea A, Kahmark K, Shcherbak I, Robertson GP, Ortiz-Monasterio I. Nitrous oxide (N2O) flux responds exponentially to nitrogen fertilizer in irrigated wheat in the Yaqui Valley, Mexico. Agriculture, Ecosystems & Environment. 2018;261:125–32. doi:10.1016/j.agee.2018.04.003
4. Turner NC. Turgor maintenance by osmotic adjustment, an adaptive mechanism for coping with plant water deficits. Plant, Cell & Environment. 2017;40(1):1–3. doi:10.1111/pce.12839
5. Argentel L, Garatuza J, Yépez E, Vega M, Rivera M, Garibaldi J. Eficiencia de uso de agua en trigo (Triticum durum L.) en el Valle del Yaqui en un escenario de cambio climático. Revista Latino americana de Recursos Naturales. 2017;13(2):58-65.
6. Lares MF, Robles A, Yepez EA, Handler RM. Global warming potential of intensive wheat production in the Yaqui Valley, Mexico: a resource for the design of localized mitigation strategies. Journal of Cleaner Production. 2016;127:522–32. doi:10.1016/j.jclepro.2016.03.128
7. Cavazos T, Arriaga S. Downscaled Climate Change Scenarios for Baja California and the North American Monsoon during the Twenty-First Century. Journal of Climate. 2012;25(17):5904–15. doi:10.1175/JCLI-D-11-00425.1
8. Argentel L, Garatuza J, Armendáriz MM, Yepez EA, Arredondo T, González J. Estrés térmico en cultivo del trigo. Implicaciones fisiológicas, bioquímicas y agronómicas. Cultivos Tropicales. 2017;38(1):57-67.
9. Azcón Bieto J, Talón M. Fundamentos de fisiología vegetal [Internet]. 2.a edición. Barcelona, España: Ed. McGrow-Hill-Interamericana de España; 2008 [cited 2018 Jul 6]. 656 p. Available from: http://www.publicacions.ub.edu/ficha.aspx?cod=06985
10. Argentel L, Garatuza J, Yepez EA, de los Santos- S. Evaluación de la tolerancia de variedades mexicanas de trigo a la salinidad, a través de indicadores fisiológicos, bioquímicos y agronómicos, cultivadas en Cuba en condiciones de campo. Cultivos Tropicales. 2016;37(1):91-101.
11. Chaves NF, Gutiérrez MV. Respuestas al estrés por calor en los cultivos. I. Aspectos moleculares, bioquímicos y fisiológicos. Agronomía Mesoamericana. 2017;28(1):237-53. doi:10.15517/am.v28i1.21903
12. Kumar R, Silva L. Light ray tracing through a leaf cross section. Applied Optics. 1973;12(12):2950–2054. doi:10.1364/AO.12.002950
13. Ma BL, Dwyer LM, Costa C, Cober ER, Morrison MJ. Early prediction of soybean yield from canopy reflectance measurements. Agronomy Journal. 2001;93(6):1227–34. doi:10.2134/agronj2001.1227
14. Raun WR, Solie JB, Martin KL, Freeman KW, Stone ML, Johnson GV, et al. Growth stage, development, and spatial variability in corn evaluated using optical sensor readings. Journal of Plant Nutrition. 2005;28(1):173–82. doi:10.1081/PLN-200042277
15. Kimball BA. Using canopy resistance for infrared heater control when warming open-field plots. Agronomy Journal. 2015;107(3):1105. doi:10.2134/agronj14.0418
16. Figueroa P, Félix JL, Fuentes G, Valenzuela V, Chávez G, Mendoza JA. CIRNO C2008, nueva variedad de trigo cristalino con alto rendimiento potencial para el estado de Sonora. Revista mexicana de ciencias agrícolas. 2010;1(5):739-44.
17. Bockheim JG, Gennadiyev AN, Hartemink AE, Brevik EC. Soil-forming factors and Soil Taxonomy. Geoderma. 2014;226–227:231–7. doi:10.1016/j.geoderma.2014.02.016
18. Prakash L, Prathapasenan G. Effect of NaCl salinity and putrescine on shoot growth, tissue ion concentration and yield of rice (Oryza sativa L. var. GR-3). Journal of Agronomy and Crop Science. 1988;160(5):325–34. doi:10.1111/j.1439-037X.1988.tb00630.x
19. Yoshida S, Forno DA, Cock JH, Gomez KA. Laboratory manual for physiological studies of rice. [Internet]. Third Edition. Manila, Philippines: The International Rice Research Institute; 1971 [cited 2018 Jul 6]. 83 p. Available from: https://www.cabdirect.org/cabdirect/abstract/19721703488
20. McCready RM, Guggolz J, Silviera V, Owens HS. Determination of starch and amylose in vegetables. Analytical Chemistry. 1950;22(9):1156–8. doi:10.1021/ac60045a016
21. Yang J, Zhang J, Liu L, Wang Z, Zhu Q. Carbon remobilization and grain filling in japonica/indica hybrid rice subjected to postanthesis water deficits. Agronomy Journal. 2002;94(1):102–9. doi:10.2134/agronj2002.1020
22. Inman D, Khosla R, Mayfield T. On-the-go active remote sensing for efficient crop nitrogen management. Connolly C, editor. Sensor Review. 2005;25(3):209–14. doi:10.1108/02602280510606499
23. Kolmogorov A. Basic Concepts of Probability Theory. Berlin: Julius Springer; 1933. 62 p.
24. Fisher S Ronald Aylmer. The design of experiments [Internet]. 2nd ed. Londres, Inglaterra: Edinburgh : Oliver and Boyd; 1937 [cited 2018 Jul 6]. 260 p. Available from: https://trove.nla.gov.au/version/19867660
25. Tukey JW. A survey of sampling from contaminated distributions. In: Contribution to probability and statistics: Essays in honor to Harold Hotelling. Stanford University Press, Estados Unidos: Ingram Olkin, ed.; 1960. p. 448–85. (STRG Technical report ; 33).
26. STATISTICA Software. StatSoft (data analysis software system) [Internet]. Version 12. Tulsa, USA; 2013 [cited 2018 Jul 9]. Available from: https://statisticasoftware.wordpress.com/2013/05/15/statsoft-releases-version-12-of-statistica-software/
27. Minguet EG, Alabadí D, Blázquez MA. Gibberellin implication in plant growth and stress responses. In: Tran L-SP, Pal S, editors. Phytohormones: A Window to Metabolism, Signaling and Biotechnological Applications [Internet]. New York, NY: Springer New York; 2014 [cited 2018 Jul 6]. p. 119–61. doi:10.1007/978-1-4939-0491-4_5
28. Bita CE, Gerats T. Plant tolerance to high temperature in a changing environment: scientific fundamentals and production of heat stress-tolerant crops. Frontiers in Plant Science. 2013;4:273. doi:10.3389/fpls.2013.00273
29. Dwivedi SK, Basu S, Kumar S, Kumar G, Prakash V, Kumar S, et al. Heat stress induced impairment of starch mobilisation regulates pollen viability and grain yield in wheat: Study in Eastern Indo-Gangetic Plains. Field Crops Research. 2017;206:106–14. doi:10.1016/j.fcr.2017.03.006
30. Galili G, Amir R, Fernie AR. The regulation of essential amino acid synthesis and accumulation in plants. Annual Review of Plant Biology. 2016;67(1):153–78. doi:10.1146/annurev-arplant-043015-112213
31. Kusvuran S, Kiran S, Ellialtioglu SS. Antioxidant enzyme activities and abiotic stress tolerance relationship in vegetable crops. In: Shanker AK, Shanker C, editors. Abiotic and biotic stress in plants - recent advances and future perspectives [Internet]. 1st ed. Croatia: InTech; 2016 [cited 2018 Jul 9]. p. 481–503. doi:10.5772/62235
32. González LM. Apuntes sobre la fisiología de las plantas cultivadas bajo estrés de salinidad. Cultivos Tropicales. 2013;23(4):47–57. doi:10.1234/ct.v23i4.645
33. Tofiño A, Romero HM, Ceballos H. Effect of abiotic stress on starch synthesis and degradation. A review. Agronomía Colombiana. 2007;25(2):245-54.
34. Dai Z, Yin Y, Li Y, Cao L, Wang Z. Grain position in spike of wheat (Triticum aestivum L.) affects glutenin macropolymer particles distribution. Agrociencia. 2014;48(3):295–306.
35. Sperotto RA, Vasconcelos MW, Grusak MA, Fett JP. Whole-plant mineral partitioning during the reproductive development of rice (Oryza sativa L.). Spanish Journal of Agricultural Research. 2017;15(2):e0802-11. doi:10.5424/sjar/2017152-10332
36. H. RR, M. KM, R. AD, R. PB, H. S, N. HG. Biochemical and physiological constituents and their correlation in wheat (Triticum aestivum L.) genotypes under high temperature at different development stages. International Journal of Plant Physiology and Biochemistry. 2017;9(1):1–8. doi:10.5897/IJPPB2015.0240
37. Vicente R, Martínez-Carrasco R, Pérez P, Morcuende R. New insights into the impacts of elevated CO2, nitrogen, and temperature levels on the regulation of C and N metabolism in durum wheat using network analysis. New Biotechnology. 2018;40((Part B)):192–9. doi:10.1016/j.nbt.2017.08.003
38. Vishwakarma MK, Arun B, Mishra VK, Yadav PS, Kumar H, Joshi AK. Marker-assisted improvement of grain protein content and grain weight in Indian bread wheat. Euphytica. 2016;208(2):313–21. doi:10.1007/s10681-015-1598-6
39. Bernard SM, Habash DZ. The importance of cytosolic glutamine synthetase in nitrogen assimilation and recycling. New Phytologist. 2009;182(3):608–20. doi:10.1111/j.1469-8137.2009.02823.x
40. Barlow KM, Christy BP, O’Leary GJ, Riffkin PA, Nuttall JG. Simulating the impact of extreme heat and frost events on wheat crop production: A review. Field Crops Research. 2015;171:109–19. doi:10.1016/j.fcr.2014.11.010
41. Shirdelmoghanloo H, Cozzolino D, Lohraseb I, Collins NC. Truncation of grain filling in wheat (Triticum aestivum) triggered by brief heat stress during early grain filling: association with senescence responses and reductions in stem reserves. Functional Plant Biology. 2016;43(10):919–30. doi:10.1071/FP15384
42. Maich RH, Steffolani ME, Di Rienzo JA, León AE. Association between grain yield, grain quality and morpho-physiological traits along ten cycles of recurrent selection in bread wheat (Triticum aestivum L.). Cereal Research Communications. 2017;45(1):146–53. doi:10.1556/0806.44.2016.036
43. Argentel L, Garatuza J, Ontiveros MMA, Yépez EA, Garibaldi JM, Ortiz JE, et al. Caracteres fisiológicos y agronómicos de la variedad de trigo cristalino CIRNO C2008 confirman su estabilidad genética. Agrociencia. 2018;52(3):419–35.
44. Dwivedi R, Prasad S, Jaiswal B, Kumar A, Tiwari A, Patel S, et al. Evaluation of wheat genotypes (Triticum aestivum L.) at grain filling stage for heat tolerance. International Journal of Pure & Applied Bioscience. 2017;5(2):971–5. doi:10.18782/2320-7051.2614
45. Abdelrahman M, El-Sayed M, Jogaiah S, Burritt DJ, Tran L-SP. The “STAY-GREEN” trait and phytohormone signaling networks in plants under heat stress. Plant Cell Reports. 2017;36(7):1009–25. doi:10.1007/s00299-017-2119-y
46. Duan T, Chapman SC, Guo Y, Zheng B. Dynamic monitoring of NDVI in wheat agronomy and breeding trials using an unmanned aerial vehicle. Field Crops Research. 2017;210:71–80. doi:10.1016/j.fcr.2017.05.025
Recibido: 07/05/2018
Aceptado: 05/07/2018
Jaime Garatuza-Payán. Instituto Tecnológico de Sonora. Dirección de Recursos Naturales, 5 de febrero, 818 Sur, Col. Centro, Cd. Obregón, Sonora. México. CP: 85000.Email: garatuza1@gmail.com


{kind=link}
{kind=link}
{kind=link}
{kind=link}