










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkBiotecnología Aplicada
versión On-line ISSN 1027-2852
Biotecnol Apl vol.34 no.3 La Habana jul.-set. 2017
REPORT
New methodology for Vip3A monitoring in corn resistant to Spodoptera frugiperda (Lepidoptera: Noctuidae)
Nueva metodología para el monitoreo de Vip3A en plantas de maíz resistentes a Spodoptera frugiperda (Lepidoptera: Noctuidae)
Daily Hernández, Hasel Aragón, Marcos González, Lianet Rodríguez-Cabrera, Milagros Ponce, Andrés Tamayo, Pilar Téllez, Camilo Ayra-Pardo, Rodolfo Valdés
Centro de Ingeniería Genética y Biotecnología, Ave 31 e/ 158 y 190, Cubanacán, Playa, CP 11600, La Habana, Cuba.
ABSTRACT
Here we report the main research results demonstrating the expression of the insecticidal toxin Vip3Aa in transgenic Bt-Vip3Aa corn hybrids, at ‘high dose’ according to the Environmental Protection Agency of the USA. It was also developed a quantitative ELISA assay named TOXIVip, specific against the Vip3Aa protein. The Vip3Aa toxin was cloned and expressed in Escherichia coli (Ec-Vip3Aa), purified by Immobilized Metal Affinity Chromatography (IMAC) and used for the generation of monoclonal antibodies (mAbs). The mAbs were able to recognize the native Vip3Aa protein expressed in transgenic corn plants and were used in the development of the TOXIVip method. Specifically, the TOXIVip method allowed quantifying the expression of the toxin in Bt-Vip3Aa corn hybrids, and aided on corroborating them as complying with the ‘high dose’ criterion. So far, the TOXIVip method was the first quantitative system ever reported for selecting the best Bt-Vip3Aa corn plants showing insecticidal activity according to the highest levels of the Vip3Aa toxin expressed. Advantageously, the TOXIVip method provides a fast and specific procedure to determine the Vip3Aa protein in transgenic plants, particularly in Bt-Vip3Aa corn plants, and also in the environment and as part of the screening tests in the food stock chain. Such system could be determinant to establish the ‘high dose’ criterion during the introduction of new lines and hybrids of Bt-Vip3A corn, also helping in genetic introgression analyses while developing new corn genotypes. This research granted the 2016 Award of the Cuban National Academy of Sciences.
Keywords: Vip3A ELISA, Bt-Vip3A hybrids, high dose, corn.
RESUMEN
Se presentan los principales resultados de investigación que demostraron la producción de la toxina insecticida Vip3A en plantas transgénicas de maíz híbrido Bt-Vip3A, a altas dosis según los requerimientos de la Agencia de Protección Ambiental de EE.UU. También se desarrolló un ensayo de ELISA específico contra la proteína Vip3A, denominado método TOXIVip. Para ello, la Vip3A se clonó y expresó en Escherichia coli (Ec-Vip3Aa), se purificó mediante cromatografía de afinidad por quelatos metálicos y se uso para la obtención de anticuerpos monoclonales (mAbs). Los anticuerpos fueron capaces de reconocer a la toxina Vip3A nativa, expresada en las plantas transgénicas y con ellos se desarrolló el método TOXIVip. Este ensayo permitió cuantificar la expresión de la toxina en plantas híbridas Bt-Vip3Aa, así como el cumplimiento del criterio de altas dosis. Hasta el momento, el método TOXIVip es el primer sistema conocido que permita la selección de las mejores plantas Bt-Vip3Aa con actividad insecticida, según los altos niveles de toxina Vip3Aa expresada. Ventajosamente, el método TOXIVip proporciona un procedimiento rápido y específico para determinar la proteína Vip3Aa en plantas transgénicas, especialmente en plantas Bt-Vip3Aa de maíz, así como en el medioambiente y en la cadena de productos alimentarios. Tal sistema puede ser determinante para establecer el criterio de alta dosis durante la introducción de nuevas líneas e híbridos de maíz Bt-Vip3A, y de ayuda en los análisis de introgresión genética durante el desarrollo de nuevos genotipos de maíz. Este trabajo mereció el Premio Anual de la Academia de Ciencias de Cuba para el año 2016.
Palabras clave: Vip3A ELISA, híbridos Bt-Vip3A, alta dosis, maíz.
INTRODUCTION
The indiscriminate and long-term use of corn plants that express Bacillus thuringiensis toxins (plants-Bt), have been the causes of the emergence of resistant insects [1]. In this sense, for the management of insects resistant to Bt toxins, several strategies have been drawn up, among which are combining several Bt toxins in the same plant (Cry1/Vip3A), and development of self-pesticide plants, with ‘high dose’ of expression of one of the Bt toxins [2].
Therefore, determining the ‘high dose’ criterion in Bt plants plays a key role in resistance management by insect pests. In this regard, the criterion ‘high dose’ was described for the first time by the Environmental Protection Agency of the USA (EPA) and refers to the concentration of the insecticidal protein (Vip3Aa) expressed in the Bt/Vip3Aa corn plants must be equal to or greater than 25 times the required concentration of the insecticidal protein to eliminate 99.99 % of the population of susceptible insects present in the field [3]. To do this, lyophilized Bt-Vip3Aa corn leaves have been used, diluting the powder at 1:25 in the artificial diet of susceptible insects. The main limitation of this strategy is the possible synergistic effect of the toxin with secondary metabolites of the leaves generated during lyophilization. Moreover, it takes a relatively long period of 7-10 days for obtaining the final result, and it requires an artificial colony of insects that could differ genetically from insects found in the field in terms of susceptibility [4].
The quantification of the Vip3Aa toxin in lines and hybrids of Bt-Vip3Aa corn was a critical point in this study because the technology available in the world market is destined only for the detection of the Vip3Aa toxin in corn plants. Therefore, a new methodology called TOXIVip was developed to detect, quantify and select corn plants that express the Vip3Aa toxin in ‘high doses’. In the other hand, the Vip3Aa toxin was cloned into an expression vector in Escherichia coli (Ec-Vip3Aa), purified and used more for the generation of mAbs. These specific mAbs to the Ec-Vip3Aa toxin were used to develop the TOXIVip method aimed at the quantification of the Vip3Aa toxin and selection of Bt-Vip3Aa corn plants that meet the ‘high dose’ criterion.
RESULTS
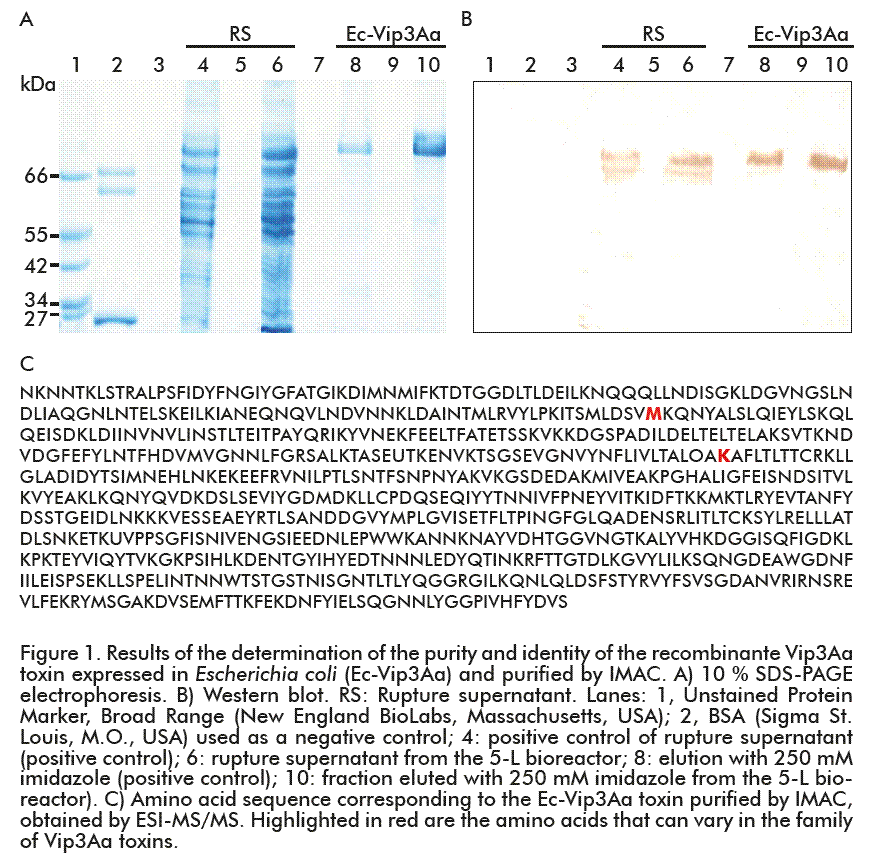
The Ec-Vip3Aa toxin was cloned into the pQE-30 expression vector in E. coli strain JM-109, under the control of the strong pTaq promoter [5]. Protein expression was induced by adding IPTG, and the Ec-Vip3Aa toxin was fused to a histidine leader. This last facilitated the isolation of the Ec-Vip3Aa toxin from the rupture supernatant (RS), mediated by the interaction that was established between the groups imidazole of the histidines and the metal fixed in the IMAC chromatography matrix used (Figure 1). Therefore, after cell rupture and centrifugation, the Ec-Vip3Aa toxin was purified from RS, 97.2 ± 2.8 % pure and for a final recovery of 50.4 ± 4.0 %. These parameters were sufficient to generate the mAb required for the development of the TOXIVip method with the aim of detecting and quantifying the Vip3Aa toxin in Bt-Vip3Aa corn plants.
The Ec-Vip3Aa toxin was sequenced for identity analysis, the sequence further confirmed by mass spectrometry (Figure 1) and further compared to those reported for the Vip3Aa toxin in the Bacillus thuringiensis (Bt) database [6]. It was demonstrated that both polypeptide sequences were similar, the Ec-Vip3Aa toxin with 789 amino acids [5, 7-9].
It was also checked whether the Ec-Vip3Aa toxin is biologically active against S. frugiperda larvae [10-12]. Toxicity is defined as the lethal concentration of the Ec-Vip3Aa toxin that causes death in 99 % (LC99) of the susceptible and exposed insect population. Data obtained from the bioassay were analyzed using the Probit statistical method and it was shown that the Ec-Vip3Aa toxin was biologically active against larvae of the corn moth, with a LC99 of 6.680 ppm (Table 1).
The LC99 value measured in the biological activity assay of the Ec-Vip3Aa toxin will allow the subsequent selection of Bt-Vip3Aa corn plants that meet the ‘high dose’ criterion. The strategy proposed in this study for the selection of Bt-Vip3Aa corn plants expressing the Vip3Aa toxin at high doses was divided into two fundamental steps: 1) determination of the LC99 of the Ec-Vip3Aa toxin, 2) direct quantification of the Vip3Aa toxin in Bt-Vip3Aa corn plants using the TOXIVip method.
For the development of the method it was necessary to generate specific mAbs to the Ec-Vip3Aa toxin, so it was used as an immunogen. Up to 50 μg of the Ec- Vip3Aa toxin were administered in three subcutaneous immunizations at 15-day intervals, the first was completed with 100 μL of complete Freund’s adjuvant and the remaining two in incomplete Freund’s adjuvant [13, 14]. A booster dose was applied intraperitoneally, 72 h before the extraction of the B cells and fusion with the myeloma cells (1: 1 v/v) to generate the respective hybridomas. Cells producing the anti-Ec-Vip3Aa antibodies were subjected to several cloning steps using the limiting dilution technique, whereas hybridomas were selected by a qualitative DAS-ELISA. The hybrid cells were inoculated intraperitoneally in Balb/c mice and the ascites was collected ten days after the inoculation [13, 15]. The anti-Ec-Vip3Aa antibodies were purified by affinity chromatography, using a Sepharose Fast-Flow Protein A matrix (GE Healthcare, Uppsala, Sweden) (Table 2).
The specific mAbs to the Ec-Vip3Aa toxin showed selectivity, specificity and affinity for the Ec-Vip3Aa toxin (Table 2), and they were used for the development of the TOXIVip method [7]. The quantification of the Vip3Aa toxin in Bt-Vip3Aa corn plants is limited as there are no quantification systems available on the market that allow direct quantification of this Bt toxin. The immunoassays are fast and effective methods of high precision and specificity for the desired analyte [16, 17].
In the other hand, in this study it is proposed for the selection of corn plants that meet the ‘high dose’ criterion, to combine the dose/response bioassay with the TOXIVip method developed to quantify the Vip3Aa toxin. The advantage of the combined use of the TOXIVip bioassay for the selection of Bt-Vip3Aa corn plants with high dose expression of the Vip3Aa toxin lies in the non-interference of the secondary metabolites generated during the corn leaves lyophilization process [18]. It is performed in a shorter period and does not need specialized equipment. The TOXIVip method could also be useful to assess possible changes in Vip3Aa toxin levels in Bt-Vip3Aa corn plants exposed to different abiotic stress agents, to assist corn breeders during the process of introgression of new Bt-Vip3Aa corn plants, as well as on assessing the impact on the environment of the dissemination of transgenic characters [19, 20].
The larvae of S. frugiperda are characterized by consuming foliar tissue in plants [21]. However, the newborn larvae feed on the same mass of eggs to which they belonged, and then consume the foliar tissue on one side, without perforating it, leaving the epidermal layer of the leaf bundle intact [22]. From the second or third stage, the feeding seems more voracious, leaving a trail of perforations in the leaves. The last stages can cause complete defoliation. In Cuba, S. frugiperda is the main pest of corn cultivation. The life cycle of the corn goes through the vegetative growth stage (V), which is divided into sub-stages according to the number of leaves of the plant that goes from V1-V16 and the reproductive phase (VT/R) [23]. The monitoring of variations on the level of expression of the Vip3Aa toxin in Bt-Vip3Aa corn plants, meeting the ‘high dose’ criterion, becomes a necessity for the control of insects resistant to Bt-Vip3Aa corn plants.
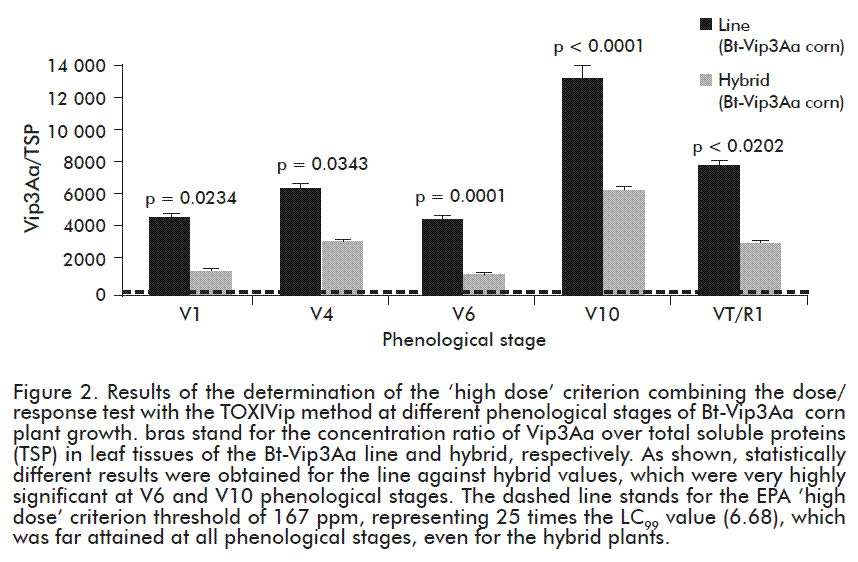
The quantification of the Vip3Aa toxin was performed in the foliar tissue of the Bt-Vip3Aa corn plants using the TOXIVip method. Significantly higher differences were found in the expression of Vip3Aa in leaves between the line and the corn hybrid Bt-Vip3A (p < 0.05) at phonological stages included from V1, V4, V6 and V10 to VT/R1 (Figure 2). Such differences could be derived from changes in the copy number of the transgene between both cultivars (two in the line against one in the hybrid), or due to particular genomic influences in the resulting proteome [24, 25].
For the management of resistant insects in autopesticide crops, EPA establishes that the expression of the toxin in the Bt-Vip3A plant must be 25 times higher or equal to the toxin concentration value obtained during the bioassay that eliminates 99 % of the population of exposed insects, known as LC99. In our experimental setting, the 25 × LC99 value (25 × 6.680 ppm) was 167 ppm, while the lowest Vip3Aa expression was attained in the evaluated hybrids at 829 ppm in the V4 stage of culture (Figure 2). This confirmed that the Bt-Vip3Aa corn line and hybrids are capable of expressing the Vip3Aa toxin at the levels required by international regulatory agencies and therefore meeting the ‘high dose’ criteria.
SCIENTIFIC AND ECONOMIC RELEVANCE
In summary, the usefulness of the TOXIVip method was demonstrated for the first time, for the rapid and specific quantification of the Vip3Aa toxin. In addition, a dose-response bioassay was combined for the first time with the TOXIVip method, which allowed the selection of the best hybrids. of corn Bt-Vip3Aa plants in terms of expression levels of the Vip3Aa toxin. Therefore, it is proposed to include this methodology as a tool to:
1) The direct and short-term evaluation of the ‘high dose’ criterion in Bt-Vip3Aa corn plants to guarantee the control of resistant insects;
2) Assist corn breeders in the process of genetic introgression of Bt-Vip3Aa corn for the development of new maize genotypes;
3) Assist the regulators in the detection and quantification of Vip3Aa toxin in the environment, grain imports and in the food chain;
4) For the quality evaluation of seeds and plant varieties;
5) Analyze environmental risks when using Bt-Vip3A corn plants.
Noteworthy, the TOXIVip method developed is not commercially available yet.
This work was awarded in different phases of development, on 2013 at CIGB as Scientific-Technical Breakthrough of the Year, for the development of immunochemical tools to detect the MIR162 event in transgenic maize resistant to Spodoptera frugiperda. It was also presented in seven specialized international conferences and published in reputed international journals as well as celebrated by international experts on this field of research.
ACKNOWLEDGEMENTS
The authors thank the collaboration of the following specialists from the CIGB: Pilar Téllez, Ivis Morán, Claudia Rodríguez and Albis Riverón, from the Laboratory of Enviromental and Biotechnology; Miguel Castillo and Leonardo Gómez from the Deparment of Monoclonal Antibody Production; Victoria María Lugo from the Deparment of Technological Development, and Yassel Ramos from the Direction of Biomedical Research.
REFERENCES
1. Moar W, Roush R, Shelton A, Ferre J, MacIntosh S, Leonard BR, et al. Field-evolved resistance to Bt toxins. Nat Biotechnol. 2008;26(10):1072-4.
2. Huang F, Andow DA, Buschman LL. Success of the high dose/refuge resistance management strategy after 15 years of Bt crop use in North America. Entomol Exp Appl. 2011;140:1-16.
3. Bernardi O, Bernardi D, Ribeiro RS, Okuma DM, Salmeron E, Fatoretto J, et al. Frequency of resistance to Vip3Aa20 toxin from Bacillus thuringiensis in Spodoptera frugiperda (Lepidoptera: Noctuidae) populations in Brazil. Crop Protection. 2015;76:7-14.
4. Tabashnik BE, Carriere Y. Surge in insect resistance to transgenic crops and prospects for sustainability. Nat Biotechnol. 2017;35(10):926-35.
5. Hernández D, Rodríguez-Cabrera L, Valdés R, Morán I, Téllez P, Ayra-Pardo. Bacillus thuringiensis Vip3Aa1 Expression and Purification from E. Coli to be determined in seeds and leaves of genetically-modified corn plants. J Agron. 2013;12(4):153-67.
6. Crickmore N. The Vip nomenclature. 2016 [cited 2016 Oct 14]. Available from: http://www.lifesci.sussex.ac.uk/home/Neil_Crickmore/Bt/vip.html
7. Estruch JJ, Yu CG. Plant pest control. US patent 6,291,156 B1, 2001.
8. Lee MK, Walters FS, Hart H, Palekar N, Chen JS. The mode of action of the Bacillus thuringiensis vegetative insecticidal protein Vip3A differs from that of Cry1Ab delta-endotoxin. Appl Environ Microbiol. 2003;69(8):4648-57.
9. Syngenta Report. Solicitud de liberación experimental de maíz con la tecnología BT11 x MIR162 x MIR604 x GA21 x (SYN-BTO11-1 x SYN-IR162-4 x SYN-IR604-5 x MON-00021-9) en la región agrícola del Estado de Chiguagua durante los ciclos PV 2013 y Pv 2014. Ciudad de México: Syngenta Agro S.A. de C.V.; 2012.
10. Pardo-Lopez L, Munoz-Garay C, Porta H, Rodriguez-Almazan C, Soberon M, Bravo A. Strategies to improve the insecticidal activity of Cry toxins from Bacillus thuringiensis. Peptides. 2009;30(3):589-95.
11. Hernandez-Martinez P, Hernandez- Rodriguez CS, Rie JV, Escriche B, Ferre J. Insecticidal activity of Vip3Aa, Vip3Ad, Vip3Ae, and Vip3Af from Bacillus thuringiensis against lepidopteran corn pests. J Invertebr Pathol. 2013;113(1):78-81.
12. Palma L, Munoz D, Berry C, Murillo J, Caballero P. Bacillus thuringiensis toxins: an overview of their biocidal activity. Toxins (Basel). 2014;6(12):3296-325.
13. Gavilondo JV. Anticuerpos monoclonales. La Habana: Elfos Scientiae;1995
14. Ben Hamadou-Charfi D, Sauer AJ, Abdelkafi-Mesrati L, Jaoua S, Stephan D. Production of polyclonal and monoclonal antibodies against the Bacillus thuringiensis vegetative insecticidal protein Vip3Aa16. Appl Biochem Biotechnol. 2015;175(5):2357-65.
15. Hernández D, Aragón H, González M, González G, Tamayo A, Morán I, et al. Monoclonal antibody generation and characterization for Vip3Aa20 quantification in transgenic corn plants. BioProcessing J. 2016;15(3):30-43.
16. Grothaus GD, Bandla M, Currier T, Giroux R, Jenkins GR, Lipp M, et al. Immunoassay as an analytical tool in agricultural biotechnology. J AOAC Int. 2006;89(4):913-28.
17. Yang L, Wang C, Holst-Jensen A, Morisset D, Lin Y, Zhang D. Characterization of GM events by insert knowledge adapted re-sequencing approaches. Sci Rep. 2013;3:2839.
18. Fabrick JA, Unnithan GC, Yelich AJ, DeGain B, Masson L, Zhang J, et al. Multi-toxin resistance enables pink bollworm survival on pyramided Bt cotton. Sci Rep. 2015;5:16554.
19. Amanullah, Kakar KM, Khan A, Khan I, Shah Z, Hussain Z. Growth and yield response ofmaize (Zea mays L.) to foliar NPK-fertilizers undermoisture stress condition. Soil Environ. 2014;33(2):116-23.
20. Kachhawa, D.. (2017). Microorganisms as a biopesticides. J Entomol Zool Studies. 2017;5(3):468-73.
21. Lemes ARN, Figueiredo CS, Sebastiao I, Marques da Silva L, da Costa Alves R, de Siqueira HAA, et al. Cry1Ac and Vip3Aa proteins from Bacillus thuringiensis targeting Cry toxin resistance in Diatraea flavipennella and Elasmopalpus lignosellus from sugarcane. PeerJ. 2017;5:e2866.
22. Silva DM, Bueno AF, Andrade K, Stecca CS, Oliveira PM, Oliveira MCN. Biology and nutrition of Spodoptera frugiperda (Lepidoptera: Noctuidae) fed on different food sources. Scientia Agricola. 2017;74(1):18-31.
23. Munkvold GP, White DG. Compendium of Corn Diseases. Fourth Edition. St. Paul, MN: American Phytopathological Society; 2016.
24. Herrero S, Bel Y, Hernandez-Martinez P, Ferre J. Susceptibility, mechanisms of response and resistance to Bacillus thuringiensis toxins in Spodoptera spp. Curr Opin Insect Sci. 2016;15:89-96.
25. Marques LH, Castro BA, Rossetto J, Silva OA, Moscardini VF, Zobiole LH, et al. Efficacy of Soybean’s Event DAS-81419-2 Expressing Cry1F and Cry1Ac to Manage Key Tropical Lepidopteran Pests Under Field Conditions in Brazil. J Econ Entomol. 2016;109(4):1922-8.
Received in October, 2016.
Accepted in March, 2017.
Daily Hernández. Centro de Ingeniería Genética y Biotecnología, Ave 31 e/ 158 y 190, Cubanacán, Playa, CP 11600, La Habana, Cuba. E-mail: daily.hernandez@cigb.edu.cu.


{kind=link}
{kind=link}
{kind=link}
{kind=link}