










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Farmacia
versión On-line ISSN 1561-2988
Rev Cubana Farm v.44 n.3 Ciudad de la Habana jul.-sep. 2010
ARTÍCULOS DE REVISIÓN
Función del peróxido de hidrógeno en el vitíligo
Function of hydrogen peroxide in cases of vitiligo
Nubia Fernández Hernández;I Gregorio Martínez Sánchez;II Manuel Enrique Cedeño PalenzuelaIII
IMaestra en Ciencias. Investigadora Agregada. Centro de Estudios para las Investigaciones y Evaluaciones Biológicas. Instituto de Farmacia y Alimentos. (CEIEB-IFAL). Universidad de La Habana. La Habana, Cuba.
IIDoctor en Ciencias Farmacéuticas. Investigador Titular. CEIEB-IFAL. Universidad de La Habana. La Habana, Cuba.
IIIMaestro en Ciencias. Investigador Agregado. CEIEB-IFAL. Universidad de La Habana. La Habana, Cuba.
RESUMEN
El vitíligo es un desorden de la pigmentación que tiene una incidencia entre el 0,5 y 3 % en la población mundial. Se caracteriza por una despigmentación de la piel provocada por la presencia de melanocitos afuncionales en la epidermis. Cuando ocurren eventos desencadenantes del estrés oxidativo, las concentraciones de peróxido de hidrógeno se incrementan ostensiblemente en melanocitos y queratinocitos. Los queratinocitos actúan como fuente de especies reactivas de oxígeno y transfieren a los melanocitos grandes cantidades de peróxido de hidrógeno. Evidencias experimentales han demostrado que como consecuencias de las altas concentraciones de peróxido de hidrógeno en la epidermis de pacientes con vitíligo, se afectan las concentraciones de butirilcolinesterasa y acetilcolineterasa, la actividad enzimática en el ciclo de las 6HB4, los péptidos derivados de proopiomelanocortinas y las proteínas del estrés entre otros muchos eventos. Todos estos eventos, tras una situación precipitante sobre un individuo genéticamente predispuesto, podrían promover cambios en la pigmentación de la piel que lo conducen, en última instancia, al vitíligo.
Palabras clave: Peróxido de hidrógeno, vitíligo, estrés oxidativo.
ABSTRACT
Vitiligo is a pigmentation disorder with an incidence between the 0.5 and the 3 % at worldwide. It is characterized by a skin depigmentation provoked by presence of dysfunctional melanocytes in epidermis. When oxidative stress triggering events are present, hydrogen peroxide concentrations clearly increase in the melanocytes and keratinocytes. The keratinocytes act as a source of oxygen and transfer to melanocytes high amounts of hydrogen peroxide. Experimental evidences have demonstrated that due to the high hydrogen peroxide concentrations in epidermis of vitiligo patients affecting the concentrations of butyryl-cholinesterase and the acetylcholinesterase, the enzymatic activity in 6HB4 cycle, propiomelanocortins and stress protein among many events. All these events after a hasty situation on a genetically predisposing subject could to promote changes in skin pigmentation that as a last resort leading to vitiligo.
Key words: Hydrogen peroxide, vitiligo, oxidative stress.
INTRODUCCIÓN
El vitíligo afecta aproximadamente al 2 % de la población mundial. Es una enfermedad asintomática que no tiene distinción de raza ni edad aunque frecuentemente aparece antes de los 20 años. El vitíligo es una hipomelanosis circunscrita caracterizada por la aparición de manchas acrómicas. Esta hipomelanosis se extiende a cualquier parte del cuerpo aunque prevalece en zonas periorificiales. Se clasifica fundamentalmente de 2 formas: generalizado o segmentario. Es una enfermedad de fácil diagnóstico con luz de Wood. Desde el punto de vista histológico se plantea que hay una ausencia de melanocitos en la epidermis de individuos enfermos, sin embargo, parece que los melanocitos están presentes pero en un estadio afuncional.1
El agente causal de la enfermedad es tema de estudio desde hace más de 3 décadas. Diferentes grupos de investigadores proponen varias teorías para explicar la o las causas que le dan origen. Cada grupo, aferrado a sus resultados y con argumentos sólidos, explica el posible origen de esta enfermedad pero resulta necesaria la integración de todos los hallazgos científicos para lograr claridad en el tema.
De manera general se postulan varias teorías: teoría autoinmune, teoría citotóxica, teoría neural, teoría genética y teoría de la convergencia. En algunos estudios se concluye que la muerte del melanocito es debida a una destrucción autoinmune, otros sugieren que es la excesiva formación de precursores de melanina los que podrían destruir al melanocito o la presencia de un mediador neuroquímico podría inhibir la producción de melanina. Los que defienden la causa genética no han podido demostrar con claridad este evento. Recientemente se plantea que la destrucción del melanocito ocurre por la sucesión de eventos a partir de una situación precipitante sobre un individuo genéticamente predispuesto.
El propósito de este trabajo es la integración de varios eventos ya probados a un denominador común: la demostrada presencia de peróxido de hidrógeno (H2O2) en concentraciones incrementadas en la epidermis de pacientes con vitíligo y las concentraciones disminuidas de catalasa tanto en epidermis como en sangre.
La búsqueda y localización de la información, incluyó una revisión de artículos científicos en la Base de Datos MEDLINE, entre 2000-2009, para lo cual se utilizaron en lo fundamental los descriptores siguientes: melanocitos, vitíligo, hipopigmentación, radicales libres, peróxido de hidrógeno. Se localizaron las fuentes de información primaria (artículos originales). La búsqueda bibliográfica incluyó artículos científicos de revisión y de resultados experimentales.
MELANOCITOS-MELANOGÉNESIS. GENERALIDADES
Los melanocitos son células dendríticas que migran hacia la capa basal de la epidermis de la piel, el folículo piloso, el iris de los ojos, el oído interno y las leptomeninges. Su origen, común al de las células nerviosas, es el ectoderma de la cresta neural.2 Se caracterizan por su núcleo ovoide y en condiciones normales los melanocitos se disponen contactando con los queratinocitos por medio de sus dendritas.
La unidad melánica-epidérmica, llamada así desde 1963 por Fitzpatrick y Breathnach,3 describe un melanocito cuyas dendritas se expanden y contactan con 36 queratinocitos aproximadamente donde se descarga la melanina. Esta proporción se mantiene constante independientemente de la raza. La principal función de los melanocitos es la melanogénesis, es decir, la producción de melanina, pigmento que determina el color de la piel humana, los ojos y el pelo. La síntesis de melanina en el ser humano determina la pigmentación cutánea constitutiva o color de piel y la pigmentación facultativa o capacidad para broncearse y tiene además una función fotoprotectora muy importante, aunque esta potencia fotoprotectora es pequeña, estimándose similar a la de un factor de protección solar de 1,5-2, lo que supone una neutralización del 40-50 % de los rayos ultravioletas (UV).
Las melaninas pueden ser eumelaninas (color marrón parduzco) o feomelaninas (pigmentos rojo amarillento) en dependencia de la interacción del receptor 1 de la melanocortina (MC1R) presente en la membrana de los melanocitos y la hormona estimulante del melanocito alfa (MSH-a). La MSH-a controla intracelularmente la formación de L-tirosina a partir de la L-fenilalanina tanto en melanocitos como en queratinocitos y la actividad tirosinasa en melanocitos.4
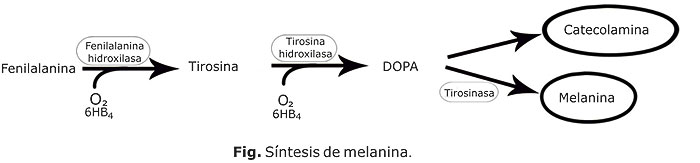
La síntesis de la melanina consta de una serie de reacciones enzimáticas que tienen lugar en el interior de organelos especializados llamados melanosomas. La tirosinasa (T), con dos isoformas, la proteína 1 y la 2 relacionada con tirosinasa (TRP-1 y la TRP-2), son enzimas que intervienen en la melanogénesis. En los melanocitos, la tirosina hidroxilasa transforma la tirosina en DOPA lo que activa la T; posteriormente la DOPA se puede convertir en dopaquinona, a partir de la cual se generan las melaninas. (figura) La TRP-1 y TRP-2
intervienen en la síntesis de eumelaninas, mientras que las feomelaninas se obtiene mediante la incorporación de derivados sulfatados por una vía anabólica alternativa.5
A medida que se va produciendo la maduración de los gránulos en los melanosomas, estos serán transportados hacia el extremo distal de las dendritas melanocíticas, desde donde serán transferidos a los queratinocitos.
La pigmentación melánica está predeterminada genéticamente y es regulada por la radiación UV o por diversos factores químicos. La radiación UV es el principal estímulo para la melanogénesis ya sea de forma directa sobre los melanocitos o de forma indirecta mediante la inducción de la secreción de factores activadores de la melanogénesis por parte de los queratinocitos y los melanocitos. Estos activadores de la melanogénesis son la MSH-a, la hormona adrenocorticotropa (ACTH) y el óxido nítrico (NO).
QUERATINOCITOS COMO FUENTE DE H2O2
En 1991 se evaluaron las concentraciones basales de H2O2 en queratinocitos de células epidérmicas normales y las compararon con las concentraciones de H2O2 en melanocitos en iguales condiciones. Los resultados mostraron que los valores de H2O2 en los queratinocitos eran superiores a los encontrados en melanocitos.6 Los melanocitos son particularmente sensibles al estrés oxidativo y aunque ellos son capaces de utilizar las especies reactivas de oxígeno durante la melanogénesis, la modulación del H2O2 parece ser determinante en este proceso, por lo que se requiere un balance redox preciso.7,8
En las células epidérmicas se incrementan las concentraciones de H2O2 en respuesta al estrés oxidativo provocado por radiaciones, procesos inflamatorios, procedimientos quirúrgicos, trauma mecánico, entre otros factores. Los mecanismos antioxidantes endógenos pueden no ser suficientes para contrarrestar el efecto deletéreo que provocan las sustancias oxidantes y dicho estrés puede conducir a la disrupción de la función celular.
En 1987 se demostró que la molécula de H2O2 es capaz de permear a través de la membrana celular, incluso por difusión pasiva.9 Dada la relación entre la cantidad de queratinocitos y la de melanocitos en la epidermis humana (unidad melánica epidérmica: 36:1), las concentraciones de H2O2 en ambos tipos de células fueron comparadas y luego fue medida la transferencia de esta molécula desde los queratinocitos a los melanocitos.10
Los resultados de estos estudios demostraron que los valores basales de H2O2 en queratinocitos son superiores al de los melanocitos y que estos podrían constituir la fuente de especies reactivas de oxígeno de los melanocitos transfiriéndose a estos desde los propios queratinocitos.11 Esto podría estar involucrado en la destrucción o empeoramiento de la función del melanocito en pacientes con vitíligo, pues los melanocitos de las lesiones de vitíligo podrían tener mayor sensibilidad a los valores normales de H2O2.10
ENZIMAS ANTIOXIDANTES EN EL VITÍLIGO
A nivel celular ocurren procesos biológicos endógenos que dan como resultado sustancias oxidantes. El organismo posee sistemas enzimáticos y no enzimáticos que protegen a las células del daño que pudiera provocar un desbalance de este equilibrio redox. En pacientes con vitíligo se han estudiado enzimas antioxidantes como catalasa (CAT), NADPH oxidasa, MAO, NO sintasa, superóxido dismutasa (SOD) y glutatión peroxidasa (GPx), tanto en eritrocitos como en tejido (cuadro). En estos pacientes la actividad de SOD se ha encontrado significativamente incrementada y la actividad de la GPx disminuida.12-14 La CAT y la GPx son las enzimas que remueven el H2O2, por lo que resultan importantes reguladoras del estrés oxidativo. Su actividad disminuida provoca acumulación de H2O2. En 1991, se reportó en pacientes con vitíligo, una reducción en la actividad de CAT a nivel epidérmico.15 Otros estudios realizados en 2004, no encontraron actividad de esta enzima en eritrocitos. 16
INFLUENCIA DEL INCREMENTO DE LAS CONCENTRACIONES DE H2O2 SOBRE LA EPIDERMIS
Estudios donde se ha examinado la unión de calcio a la calmodulina en presencia de H2O2 han revelado que la calmodulina oxidada pierde la capacidad para activar la ATPasa dependiente de calcio, lo cual implica una captación disminuida de L-fenilalanina en la epidermis. Estos estudios que se realizaron en pacientes con vitíligo agudo demostraron que la actividad y expresión de la catalasa epidérmica están disminuidas y por tanto la homeostasis del calcio de queratinocitos y melanocitos está alterada.17
En 2004, se comprobó que en la epidermis de pacientes con vitíligo se acumulan concentraciones de H2O2 en un intervalo de 10-3M unido a una disminución de la actividad y la expresión de la enzima catalasa debido a la desactivación de su sitio activo.18 La disminución de la catalasa incrementa las concentraciones de H2O2 en la epidermis. El H2O2 es un inhibidor reversible de la tirosinasa humana y juega un importante papel en la regulación (concentración-dependiente) de señales colinérgicas epidérmica.19
H2O2 y colinesterasas
Las señales colinérgicas en células no neuronales es comparable con la neurotransmisión. La disfunción del sistema colinérgico no neuronal está involucrado en la patogénesis de enfermedades inflamatorias.20
La acetilcolina (Ach) y sus receptores están presentes en una gran variedad de células entre las que se encuentran melanocitos, queratinocitos, fibroblastos, células endoteliales y células del sistema inmune. Por esta razón están involucradas en funciones básicas de la piel como son la diferenciación de queratinocitos, la formación de la barrera epidérmica, la producción de sebo, la circulación sanguínea, la respuesta inmune, la angiogénesis, entre otras y por tanto pueden estar modificados en enfermedades como el vitíligo, psoriasis, dermatitis atópica.21,22
En el vitíligo se demostró, por espectroscopia FT-Raman (Fourier-Transform), que la cascada colinérgica está severamente afectada debido a la acumulación de H2O2 en el orden de los mM, lo que oxida los residuos de aminoácidos de proteínas y péptidos y afecta su función.23 En 2006, se demostró la presencia de butirilcolinestrasa (BchE) en la epidermis humana. En este compartimento la actividad de esta enzima es superior a la de la acetilcolinesterasa (AchE) y también está sujeta a la regulación del H2O2 de manera concentración dependiente. Se ha comprobado que bajas concentraciones de H2O2 (10-6 M) activan esta enzima por incremento de la Vmáx. mientras que mayores concentraciones (10-3 M) inhiben su actividad con una significativa disminución de la Vmáx.18
En pacientes con vitíligo agudo, los residuos de metionina y triptófano tanto de la AchE como la BchE de la epidermis, son blanco del H2O2, lo que conduce a la acumulación de Ach en la epidermis de dichos pacientes; la colinoacetiltransferasa (chAT) no se afecta. Este proceso fue revertido con un tratamiento que incluía pseudocatalasa.23 Las colinesterasas inactivadas contribuyen al mantenimiento y promoción del estrés oxidativo.
H2O2 y tetrahidrobiopterinas
Es sabido que la L-fenilalanina en el citosol de los melanocitos epidérmicos promueve la formación de L-tirosina a partir de la fenilalanina hidroxilasa. Esta tirosina es el sustrato precursor de la melanina o la adrenalina. El cofactor 6-tetrahidrobiopterina (6BH4) es producido tanto en melanocitos como en queratinocitos para regular la actividad de la tirosina hidroxilasa (TH), fenilalanina hidroxilasa (PAH) y T.24
Estudios realizados en 2004 demostraron la función del 6HB4 en la regulación de la pigmentación humana.24 Concentraciones de 6HB4 en un intervalo de 10-6 M inhibieron la actividad de la T después de ser activada con L-tirosina; este mecanismo produjo H2O2 a partir del O2. Sin embargo, en ausencia del sustrato, la 6HB4 se oxidó directamente a 7,8 dihidrobiopterina por acción de la T la que es activada a bajas concentraciones de H2O2 (< 0,3 x 10-3 M). La 6BH4 oxidada (6-biocterina) es citotóxica en melanocitos epidérmicos.25,26 La inhibición de T por exceso de 6HB4 es revertida con MSH-a por formación de un complejo 6HB4-MSH-a.24
El H2O2 derivado de cualquier fuente podría regular todos los procesos dependientes de 6BH4 entre los que se encuentran la hidroxilación de la L-fenilalanina, L-tirosina, L-triptófano, la producción de NO y varios procesos inmunomoduladores.27 Desde 1994, se reportó que en pacientes con vitíligo había una excesiva síntesis de novo de 6HB4, lo que provocaba incremento de noradrenalina tanto en plasma como en orina y acumulación de 7-tetrahidrobiopterina (7BH4) en epidermis; esto último podría bloquear el suplemento de L-tirosina de la L-fenilalanina por inhibición competitiva con la fenilalanina hidoxilasa.28 Tanto la 6HB4 como la 7BH4 favorecen el estrés oxidativo en pacientes con vitíligo. Estos pacientes acumulan concentraciones en el orden de los milimolares de H2O2 tanto en epidermis como en linfocitos y monocitos sanguíneos. El H2O2 afecta numerosas enzimas entre las que se encuentra la dihidropteridina reductasa (DHPR). Concentraciones menores de 30 µM de H2O2 incrementan la actividad de esta enzima mientras que mayores concentraciones la desactivan.29 Estos resultados se pusieron de manifiesto cuando se administró luz UVB/pseudocatalasa a pacientes con vitíligo y las concentraciones de DHPR retornaron a los valores normales luego de inhibirse el exceso de H2O2 de la epidermis.
H2O2 y proopiomelanocortinas
La MSH-a, al igual que la ACTH, son hormonas polipeptídicas derivadas de la división de la proopiomelanocortina (POMC). La piel humana tiene la capacidad para sintetizar, de forma autocrina, los péptidos derivados de la POMC. Se ha demostrado la presencia y la funcionabilidad de MSH-a y -b, ACTH, y -b endorfinas en la regulación de pigmentación de la piel.30,31 Se sabe además que los melanocitos expresan mayores concentraciones de POMC comparado con los queratinocitos.32
Los residuos de metionina de los péptidos derivados de POMC, a través de su actividad proteolítica dependiente de Ca2+, son los blancos de la oxidación provocada por la alta concentración de H2O2 presente en la epidermis de pacientes con vitíligo.31,32 Esto contribuye a la reducción epidérmica de la expresión de dichos péptidos derivados de POMC, lo que se ha podido comprobar en pacientes con un estadio temprano de vitíligo.32 En dichos pacientes quedó evidenciado que la b-endorfina oxidada pierde su función en la melanogénesis.31
La MSH-a aumenta la melanogénesis estimulando la actividad de las enzimas TRP-1 y TRP-2, mediante su unión al MC1R, principal receptor de melacortinas en la epidermis humana y al que se le ha reconocido recientemente su función como antioxidante. Tanto la MSH-a como la ACTH de melanocitos humanos poseen efectos mitogénicos y melanogénicos similares. Estos efectos están mediados por la unión a su receptor específico de MC1R; ambos poseen similar afinidad.33 La activación de MC1R es importante en la respuesta del melanocito humano a las radiaciones UV.34 Esta respuesta, que está mediada fundamentalmente por la MSH-a, es independiente de la síntesis de melanina y ocurre por la inhibición de la apoptosis del melanocito inducida por las radiaciones UV.35 La supervivencia de la MSH-a es debida a la reducción del daño al DNA inducido por dichas radiaciones.
La expresión del MC1R, así como de las enzimas involucradas en la síntesis de melanina, están reguladas por el factor de transcripción asociado a microftalmia (MITF).36 La unión de MSH a su receptor, activa la adenilato ciclasa incrementándose la concentración intracelular de AMPc lo que conduce a la activación del MITF.37 En melanocitos normales el estrés oxidativo reduce la expresión de MITF, sin embargo, esta reducción podría proteger a las células un estrés oxidativo más severo. La expresión reducida de MITF, por tanto, podría disminuir la expresión de TRP1.38
En el vitíligo los melanocitos están en un continuo estado de estrés oxidativo y la muerte celular es el resultado de fallas en el sistema antioxidante o de fallas en la regulación de la TRP-1.38 La muerte celular temprana de melanocitos en el vitíligo podría estar relacionada con el incremento de la sensibilidad al estrés oxidativo, lo cual puede estar dado por problemas en la síntesis de TRP-1 y su interacción con calnexinas.39
La reducción del MITF podría disminuir también la expresión de genes que promueven la apoptosis durante el estrés oxidativo.38 Las concentraciones de MITF podrían tener un significativo impacto en la sobrevida e iniciación de la apoptosis en el melanocito.40 Trabajos recientes demuestran que los melanocitos expresan constitutivamente Bcl-2, Bax y otras proteínas implicadas en fenómenos de apoptosis celular.41
La proteína p altera la disponibilidad del glutatión, lo que reduce la capacidad del melanocito para remover las quimiotoxinas y combatir el estrés oxidativo.38 La inhibición de la melanogénesis podría ser el resultado de la regulación negativa del MITF inducida por H2O2, lo que en última instancia podría provocar la muerte del melanocito observada en el vitíligo.42
H2O2 y proteínas del estrés
Las células estresadas por cualquier causa, se caracterizan por una elevada expresión de proteínas del estrés (PS). Se sabe que estas, en medio intracelular, tienen actividad citoprotectora pero una vez liberadas al medio extracelular, pueden inducir la respuesta inmune. Estas células sirven como antígeno en algunas enfermedades autoinmunes o pueden inducir fagocitosis y procesamiento de antígenos "chaperones" mediante células dendríticas (CD).43 Las CD ejercen su efecto mediante la expresión en su superficie, de miembros de la familia del factor de necrosis tumoral (TNF), que van a reconocer a su respectivos receptores en las células estresadas.44
En el melanocito estresado se expresa el ligando inductor de apoptosis relacionado con el TNF (TRAIL), por lo que podría comportarse como una célula tumoral y podría iniciarse una respuesta citotóxica a dicho melanocito evidenciándose, en última instancia, la despigmentación observada en el vitíligo.45 La despigmentación en el vitíligo está acompañada por la infiltración de células T a la piel dañada dada por la activación de células dendríticas las que son activadas como consecuencias de la expresión de las PS en la epidermis lesionada.46,47 Se sabe además que las PS son blancos de las quinonas. Su unión inhibe el adecuado balance redox y promueve la actividad de la p53, por lo que podría iniciarse el proceso apoptótico potenciándose la actividad quimiotóxica e inducirse un estado de estrés oxidativo.48 Las concentraciones incrementadas de H2O2 encontradas en la piel de pacientes con vitíligo podrían contribuir a la activación de las proteínas del estrés y las posibles consecuencias que de ello se derivan.
Teniendo en cuenta muchos de los hallazgos hasta ahora encontrados, la regulación del estrés oxidativo podría contribuir a la mejoría en los pacientes con vitíligo, pues los estudios más recientes enfocan esta enfermedad mediada por especies reactivas de oxígeno donde el equilibrio redox-H2O2 intracelular, controla la función del melanocito de manera concentración dependiente.49 En este ámbito la función del H2O2 desempeña una función crucial, por lo cual las terapias encaminadas a controlar sus concentraciones pudieran conducir al éxito terapéutico.
El vitíligo es el resultado de la compleja interacción entre factores genéticos, ambientales e inmunológicos, lo cual, en última instancia, conduce a la despigmentación de la piel.50 No obstante, independientemente de la causa que lo origine, existen rutas metabólicas comunes que dan como resultado la manifestación clínica observada en estos pacientes.
REFERENCIAS BIBLIOGRÁFICAS
1. Atlas de Dermatología. Desórdenes de los melanocitos. En línea. (Acceso 6 de junio de 2009). Disponible en:
2. Melanocito. En línea. (Acceso 10 de junio de 2009). Disponible en: http://es.wikipedia.org/wiki/Melanocito
3. Mina Y, Barbara AG. Melanocyte Biology: Before, During, and After the Fitzpatrick Era. J Invest Dermatol. 2004;122:215-26.
4. Schallreuter KU, Moore J, Tobin DJ, Gibbons NJ, Marshall HS, Jenner T, et al. Alpha-MSH can control the essential cofactor 6-tetrahydrobiopterin in melanogenesis (abstract). Ann N Y Acad Sci. 1999;885:329-41. (Acceso 10 de junio de 2009). Disponible en: http://www.ncbi.nlm.nih.gov/pubmed/10816664
5. Avilés JA, Lázaro P. Genetic predisposition in cutaneous melanma. Actas Dermosifiliogr. 2006;97:229-40.
6. Yohn JJ, Norris DA, Yrastorza DG, Buno IJ, Leff JA, Hake SS, et al. Disparate antioxidant enzyme activities in cultured human cutaneous fibroblasts, keratinocytes, and melanocytes (abstract). J Invest Dermatol. 1991;97:405-9. (Acceso 10 de junio de 2009). Disponible en: http://www.ncbi.nlm.nih.gov/pubmed/1875041
7. Jimbow K, Chen H, Park JS, Thomas PD. Increased sensitivity of melanocytes to oxidative stress and abnormal expression of tyrosinase-related protein in vitiligo. Br J Dermatol. 2001;144:55-65.
8. Nappi AJ, Vass E. Hydrogen peroxide generation associated with the oxidations of the eumelanin precursors 5,6-dihydroxyindole and 5,6-dihydroxyindole-2-carboxylic acid (abstract). Melanoma Res 1996; 6: 341-9. (Acceso 10 de junio de 2009). Disponible en: http://www.ncbi.nlm.nih.gov/pubmed/8908594
9. Frenkel K, Chrzan K, Ryan CA, Wiesner R, Troll W. Chymotrypsin-specific protease inhibitors decrease H2O2 formation by activated human polymorphonuclear leukocytes (abstract). Carcinogenesis 1987; 8: 1207-12. (Acceso 10 de junio de 2009). Disponible en: http://www.ncbi.nlm.nih.gov/pubmed/3621459
10. Pelle E, Mammone T, Maes D, Frenkel K. Keratinocytes Act as a Source of Reactive Oxygen Species by Transferring Hydrogen Peroxide to Melanocytes. J Invest Dermatol. 2005;124:793-7.
11. Pelle E, Huang X, Mammone T, Marenus K, Maes D, Frenkel K. Ultraviolet-B-induced oxidative DNA base damage in primary normal human epidermal keratinocytes and inhibition by a hydroxyl radical scavenger. J Invest Dermatol. 2003;121:177-83.
12. Yildrim M, Baysal V, Inaloz HS, Can M. The role of oxidants and antioxidants in generalized vitiligo at tissue level. JEADV. 2004;18:683-6.
13. Hazneci E, Karabulut AB, Oztürk C, Batçioðlu K, Doðan G, Eþrefoðlu M. A comparative study of superoxide dismutase, catalase, and glutathione peroxidase activities and nitrate levels in vitiligo patients. Int J Dermatol. 2005;44:636-40.
14. Dammak I, Boudaya MR, Ben GA, Marrekchi ES, Turki H. A comparative study of oxidant-antioxidant status in stable and active vitiligo patients. Arch Dermatol Res. 2006;298:147-52.
15. Schallreuter KU, Wood JM, Berger J. Low catalase levels in the epidermis of patients with vitiligo (abstract). J Invest Dermatol. 1991;97:1081-5. (Acceso 10 de junio de 2009). Disponible en: http://www.ncbi.nlm.nih.gov/pubmed/1748819
16. Deepali A, Shajil EM, Marfatia YS, Rasheedunnisa B. Study on the antioxidant status of vitiligo patients of different age groups in Baroda. Pigment Cell Res. 2004;17:289-94.
17. Schallreuter KU, Gobbons NC, Zothner C, About Ellof MM, Wood JM. Hydrogen peroxide-mediated oxidative stress disrupts calcium binding on calmodulin: more evidence for oxidative stress in vitiligo. Biochem Biophys Res Commun. 2007;360: 70-5.
18. Schallreuter KU, Elwary SM, Gibbons NC, Rokos H, Wood JM. Activation/deactivation of acetylcholinesterase by H2O2: more evidence for oxidative stress in vitiligo. Biochem Biophys Res Commun. 2004;315(2):502-8.
19. Schallreuter KU, Gibbons NC, Zothner C, Elwary SM, Rokos H, Wood JM. Butyrylcholinesterase is present in the human epidermis and is regulated by H2O2: more evidence for oxidative stress in vitiligo. Biochem Biophys Res Commun. 2006;349:931-8.
20. Wessler I, Kirkpatrick CJ. Acetylcholine beyond neurons: the non-neuronal cholinergic system in humans. Br J Pharmacol. 2008;154(8):1558-71.
21. Kurzen H, Wessler I, Kirkpatrick CJ, Kawashima K, Grando SA. The non-neuronal cholinergic system of human skin. Horm Metab Res. 2007;39(2):125-35.
22. Grando SA, Pittelkow MR, Schallreuter KU. Adrenergic and cholinergic control in the biology of epidermis: physiological and clinical significance. J Invest Dermatol. 2006;126(9):1948-65.
23. Schallreuter KU, Elwary S. Hydrogen peroxide regulates the cholinergic signal in a concentration dependent manner. Life Sci. 2007;80(24-25):2221-6.
24. Wood JM, Chavan B, Hafeez I, Schallreuter KU. Regulation of tyrosinase by tetrahydropteridines and H2O2 (abstract). Biochem Biophys Res Commun 2004; 325(4): 1412-7. (Acceso 10 de junio de 2009). Disponible en: http://www.ncbi.nlm.nih.gov/pubmed/15752727
25. Schallreuter KU, Moore J, Wood JM, Beazley WD, Gaze DC, Tobin DJ, et al. In vivo and in vitro evidence for hydrogen peroxide (H2O2) accumulation in the epidermis of patients with vitiligo and its successful removal by a UVB-activated pseudocatalase. J Investig Dermatol Symp Proc. 1999;4(1):91-6.
26. Schallreuter KU, Wood JM. Thioredoxin reductase - its role in epidermal redox status. J Photochem Photobiol B 2001;64(2-3):179-84.
27. Schallreuter KU, Moore J, Wood JM, Beazley WD, Peters EM, Marles LK, et al. Epidermal H(2)O(2) accumulation alters tetrahydrobiopterin (6BH4) recycling in vitiligo: identification of a general mechanism in regulation of all 6BH4-dependent processes? J Invest Dermatol. 2001;116(1):167-74.
28. Schallreuter KU, Wood JM, Lemke KR, Pittelkow MR, Lindsey NJ, et al. Defective tetrahydrobiopterin and catecholamine biosynthesis in the depigmentation disorder vitiligo (abstract). Biochim Biophys Acta 1994; 1226(2): 181-92. (Acceso 10 de junio de 2009). Disponible en: http://www.ncbi.nlm.nih.gov/pubmed/8204666
29. Hasse S, Gibbons NC, Rokos H, Marles LK, Schallreuter KU. Perturbed 6-tetrahydrobiopterin recycling via decreased dihydropteridine reductase in vitiligo: more evidence for H2O2 stress. J Invest Dermatol. 2004;122(2):307-13.
30. Dell'anna ML, Ottaviani M, Albanesi VA, Vidolin P, Leone G, Ferrano CJ. Membrane lipid alterations as a possible basis for melanocyte degeneration in vitiligo. J Invest Dermatol. 2007;127:1226-33.
31. Spencer JD, Gibbons NC, Rokos H, Peters EM, Wood JM; Schallreuter KU. Oxidative stress via hydrogen peroxide affects proopiomelanocortin peptides directly in the epidermis of patients with vitiligo. J Invest Dermatol. 2007;127(2):411-20.
32. Spencer JD, Gibbons NC, Böhm JM, Schallreuter KU. The Ca2+-binding capacity of epidermal furin is disrupted by H2O2-mediated oxidation in vitiligo (abstract). Endocrinology 2008; 149(4): 1638-45. Fecha de revisión: junio de 2009. (Acceso 10 de junio de 2009). Disponible en: http://www.curehunter.com/public/pubmed18174282.do
33. Abdel-Malek Z, Suzuki I, Tada A, Im S, Akcali C. The melanocortin-1 receptor and human pigmentation (abstract). Ann N Y Acad Sci 1999; 885: 117-33. (Acceso 10 de junio de 2009). Disponible en: http://www.ncbi.nlm.nih.gov/pubmed/10816645
34. Abdel-Malek Z, Scott MC, Suzuki I, Tada A, Im S, Lamoreux L, et al. The melanocortin-1 receptor is a key regulator of human cutaneous pigmentation. Pigment Cell Res. 2000;13(Suppl 8):156-62.
35. Kadekaro AL, Kanto H, Kavanagh R, Abdel-Malek ZA. Significance of the melanocortin 1 receptor in regulating human melanocyte pigmentation, proliferation, and survival. Ann N Y Acad Sci. 2003;994:359-65.
36. Aoki H, Moro O. Involvement of microphthalmia-associated transcription factor (MITF) in expression of human melanocortin-1 receptor (MC1R). Life Sci. 2002;71(18):2171-9.
37. Widlund HR, Fisher DE. Microphthalamia-associated transcription factor: a critical regulator of pigment cell development and survival. Oncogene. 2003;22:3035-3041. (Acceso 10 de junio de 2009). Disponible en: http://www.pubmedcentral.nih.gov/articlerender.fcgi?artid=1636837
38. Manga P, Sheyn D, Yang F, Sarangarajan R; Boissy RE. A role for tyrosinase-related protein 1 in 4-ter-butylphenol-induced toxicity in melanocytes: Implications for vitiligo. Am J Pathol. 2006;169(5):1652-62.
39. Jimbow K, Chen H, Park JS, Thomas PD. Increased sensitivity of melanocytes to oxidative stress and abnormal expression of tyrosinase-related protein in vitiligo. Br J Dermatol. 2001;144(1):55-65.
40. Larribere L, Hilmi C, Khaled M, Gaggioli C, Bille K, Auberger P, et al. The cleavage of microphthalmia-associated transcription factor, MITF, by caspases plays an essential role in melanocyte and melanoma cell apoptosis. Genes Dev. 2005;19:1980-1985.
41. Dell'anna ML, Picardo M. A review and a new hypothesis for non-immunological pathogenetic mechanisms in vitiligo. Pigment Cell Res. 2006;19:406-11.
42. Jiménez C, Martínez M, Pérez C, Daum N, Solano F, García JC. Inhibition of melanogenesis in response to oxidative stress: transient downregulation of melanocyte differentiation markers and possible involvement of microphthalmia transcription factor. J Cell Sci. 2001;114:2335-44.
43. Noessner E, Gastpar R, Milani V, Brandl A, Hutzler PJ, Kuppner MC, et al. Tumor-derived heat shock protein 70 peptide complexes are cross-presented by human dendritic cells. J Immunol. 2002;169:5424-32.
44. Lu G, Janjic BM, Janjic J, Whiteside TL, Storkus WJ, Vujanovic NL. Innate direct anticancer effector function of human immature dendritic cells. II. Role of TNF, lymphotoxin-alpha(1)beta(2), Fas ligand, and TNF-related apoptosis-inducing ligand. J Immunol. 2002;168:1831-1839.
45. Kroll T, Bommiasamy H, Boissy RE, Hernández C, Nickoloff BJ, Mestril R, et al. 4-Tertiary Butyl Phenol Exposure Sensitizes Human Melanocytes to Dendritic Cell-Mediated Killing: Relevance to Vitiligo. J Invest Dermatol. 2005;124:798-806.
46. Le Poole IC, Wañkowicz-Kaliñska A, van den Wijngaard RM, Nickoloff BJ, Das PK. Autoimmune aspects of depigmentation in vitiligo. J Investig Dermatol Symp Proc. 2004;9:68-72.
47. Ongenae K, Van Geel N, Naeyaert JM. Evidence for an autoimmune pathogenesis of vitiligo. Pigment Cell Res. 2003;16:90-100.
48. Manga P, Sheyn D, Yang F, Sarangarajan R, Boissy RE. A Role for Tyrosinase-Related Protein 1 in 4-tert-Butylphenol-Induced Toxicity in Melanocytes. Am J Pathol. 2006;169(5):1652-62.
49. Schallreuter KU. Advances in melanocyte basic science research. Dermatol Clin. 2007;25(3):283-91.
50. Spritz RA. The genetics of generalized vitiligo and associated autoimmune diseases. J Dermatol Sci. 2006;41:3-10.
Recibido: 8 de abril de 2010.
Aprobado: 17 de mayo de 2010.
M. C. Nubia Fernández Hernández. Centro de Estudios para las Investigaciones y Evaluaciones Biológicas. Instituto de Farmacia y Alimentos. (CEIEB-IFAL). Universidad de La Habana. Avenida 23 No. 21425 e/ 214 y 222, La Coronela, La Lisa, La Habana, Cuba. Correo electrónico: nubiafh@uh.cu


{kind=link}
{kind=link}