










Mi SciELO
 Servicios personalizados
Servicios personalizadosServicios Personalizados
Articulo
 Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por emailIndicadores
-
 Citado por SciELO
Citado por SciELO
Links relacionados
-
 Similares en
SciELO
Similares en
SciELO
Compartir

 Permalink
PermalinkRevista Cubana de Medicina Tropical
versión On-line ISSN 1561-3054
Rev Cubana Med Trop vol.70 no.3 Ciudad de la Habana set.-dic. 2018
ARTÍCULO ORIGINAL
Prevalencia genotípica de cagA y vacA en aislamientos de Helicobacter pylori de pacientes colombianos
Genotypic prevalence of cagA and vacA in isolations of Helicobacter pylori in Colombian patients
Paula Tatiana Uribe Echeverry, María Alejandra Acosta Cerquera, Brenda Lucia Arturo Arias, María Del Socorro Jaramillo Arredondo, Jhon Fredy Betancur Pérez, Juan Manuel Pérez Agudelo
Universidad de Manizales. Manizales, Caldas, Colombia.
RESUMEN
Introducción: El gen de la toxina vacuolizante (vacA) y el gen asociado a citotoxina (cagA) son considerados los principales factores de virulencia de Helicobacter pylori, los cuales generan mayor prevalencia e incidencia de patogenicidad.
Objetivo: Determinar la prevalencia del gen cagA y las variantes alélicas del gen vacA en cepas de H. pylori aisladas de pacientes colombianos.
Métodos: Se incluyeron 72 pacientes que fueron remitidos a endoscopia a la Clínica San Marcel de la ciudad de Manizales, Colombia, durante el segundo semestre del año 2015, a quienes se les tomó una biopsia de antro y una de cuerpo gástrico para cultivo microbiológico y posterior estudio molecular. La extracción de ADN bacteriano se realizó por el método descrito por Valentine. La identificación de los factores de virulencia cagA y vacA se efectuó por reacción en cadena de la polimerasa convencional, empleando iniciadores específicos para cada uno de ellos.
Resultados: Se obtuvieron aislamientos de Helicobacter pylori de 16 pacientes, de los cuales se identificaron cepas cagA positivas y las variantes alélicas s1 y s2 asociadas al gen vacA. No se encontró asociación estadísticamente significativa entre el diagnóstico endoscópico y los factores de virulencia de los genes cagA y vacA.
Conclusiones: Hubo una prevalencia del 50 % de cagA+ y una frecuencia 68,8 % para las variantes alélicas VacAs1m1 en los aislamientos de H. pylori obtenidos de las biopsias gástricas, lo que denota que factores diferentes y que complementan la virulencia bacteriana se asocian a la presentación de condiciones gastrointestinales, con implicaciones diagnósticas, pronósticas y terapéuticas.
Palabras clave: Helicobacter pylori; aislamiento; citotoxicidad;cagA; vacA; PCR.
ABSTRACT
Introduction: The vacuolated toxin´s gene (vacA) and the cytotoxin associated gene (cagA) are the main virulence factors of Helicobacter pylori, which generate more prevalence and incidence of patogenicity.
Objective: To determine the prevalence of cagA gene and allelic variants of vacA gene in strains of H. pylori isolated from Colombian patients.
Methods: There were 72 patients included who had been referred to endoscopy at San Marcel Clinic in Manizales city, during the second semester of the year 2015. They had antrum and gastric body's biopsy for microbiological culture and subsequent molecular study. The extraction of bacterial DNA was performed by the method described by Valentine. Identification of the virulence factors cagA and vacA genes was performed by conventional PCR, using specific primers for each of them.
Results: Isolates of H. pylori were obtained in sixteen patients, and in these were found cagA positive strains and the allelic variants associated with the vacA gene. A statistically significant association between endoscopic diagnosis and virulence factors cagA and vacA was not found.
Conclusions: It was found a prevalence of 50 % of cagA+ positive strains and a frequency of 68.8 % of the allelic variant vacAs1m1 in the isolations of H. pylori obtained from gastric biopsies, denoting different factors and that complements of bacterial virulence are associated with the presentation of gastrointestinal conditions, with diagnostic, prognostic and therapeutic implications.
Keywords: Helicobacter pylori; isolation; cytotoxicity; cagA; vacA; PCR.
INTRODUCCIÓN
Helicobacter pylori (H. pylori) es una bacteria gramnegativa, microaerofílica en forma de espiral, que posee de cinco a seis flagelos que permiten su movilización y penetración en el epitelio gástrico.1 La infección se adquiere durante la infancia y la prevalencia aumenta gradualmente con la edad, alcanzando el 50 % en los países en desarrollo; la transmisión se produce por contacto persona a persona por vía fecal/oral, oral/oral o gástrica/oral.2 Este microorganismo es considerado el principal agente causal en el desarrollo de úlcera gástrica y duodenal, gastritis crónica no autoinmune, linfoma MALT (Mucosa Associated Lymphoid Tissue) y cáncer gástrico.3,4
H. pylori posee factores de virulencia que inducen mayor prevalencia e incidencia de patogenicidad, como el gen vacA (toxina vacuolizante) y el gen cagA (asociado a citotoxina).5 El gen vacA se encuentra presente en todas las cepas,6,7 posee tres regiones polimórficas denominadas: región señal s1( subtipos s1a, s1b y s1c) y s2; región intermedia (i1 e i2) y región media (m1 y m2).8,9 Por las combinaciones alélicas de las regiones señal y media se presenta una variación considerable en la actividad vacuolizante de las diferentes cepas de H. pylori.10 Los aislamientos que presentan las combinaciones s1m1 y s1m2 producen niveles altos y moderados de citotoxina respectivamente, mientras que los aislamientos con la combinación s2m2 producen muy poca citotoxina o no la producen, razón por la cual las variaciones alélicas de vacA pueden desencadenar una acción más persistente de la bacteria y una mayor progresión clínica de la lesión gástrica como factor de riesgo para cáncer gástrico.11
El gen asociado a la citotoxina A (cag A) es uno de los 32 genes que constituyen la isla de patogenicidad Cag-PAI (principal determinante de virulencia de H. pylori); el gen cagA codifica para una proteína altamente inmunogénica, que ingresa directamente en las células del epitelio gástrico por medio del sistema de secreción tipo IV de la bacteria, interactuando con efectores celulares que conducen a la activación de vías de señalización celular promoviendo respuestas proinflamatorias y mitogénicas, alteración en las uniones intercelulares y pérdida de polaridad celular.12 Las cepas de H. pylori cagA positivas (cagA+) están relacionadas con mayor riesgo de gastritis atrófica, úlcera péptica y el incremento de desarrollar cáncer gástrico.4
Dada la importancia de los factores de virulencia vacA y cagA de H. pylori en el desarrollo de enfermedades gastroduodenales y la prevalencia de la infección informada en la ciudad Manizales (85,5 %),13 el objetivo de este estudio fue determinar la prevalencia del gen cagA y las variantes alélicas del gen vacA en cepas de H. pylori aisladas de pacientes colombianos.
MÉTODOS
Se llevó a cabo un estudio de diseño observacional, transversal y de corte analítico.
Se consideraron como criterios de inclusión: pacientes mayores de 18 años de ambos sexos, residentes en el departamento de Caldas (Colombia) con diagnóstico asociado a enfermedad ácido péptica, gastritis y úlcera gástrica o duodenal y que fueron remitidos a endoscopia digestiva alta en la Clínica San Marcel de la ciudad Manizales, Colombia. Como criterio de exclusión se consideró el consumo de inhibidores de bomba de protones, antagonistas del receptor H2, antiácidos, antibióticos y/o aspirina o AINES cuatro semanas previas al examen diagnóstico.
El periodo de inclusión de los pacientes fue el segundo semestre del 2015. La muestra fue no probabilística basada en criterios y se completó de manera consecutiva con 72 casos.
La endoscopia fue realizada por un gastroenterólogo. Durante el procedimiento se tomaron dos biopsias gástricas (una del antro y una del cuerpo del estómago), que fueron empleadas para cultivo microbiológico y posterior análisis molecular. Las biopsias se almacenaron en tubos eppendorf de 1,5 mL con caldo infusión cerebro corazón (BHI), se refrigeraron en nevera portátil, y fueron transportadas en un lapso inferior a 6 h al laboratorio de microbiología de la Universidad de Manizales para su procesamiento.
Este estudio considerado de riesgo mínimo de acuerdo con la resolución 8430 de 1993 del Ministerio de la Protección Social de Colombia, fue aprobado por el comité de ética de la Universidad de Manizales.
Cultivo microbiológico. Los medios de cultivo empleados para el aislamiento bacteriano fueron: agar TSA (Tripticasa Soya Agar-BBL™), enriquecido con sangre de cordero al 7 %, y suplemento selectivo DENT (Oxoid Limited) 4 mL/L y Agar TSA suplementado con sangre de cordero al 7 %, isovitalex 0,5 % (BBL™) y antibióticos de uso comercial: Anfotericina B x 50 mg (Bristol Myers Squibb) 3,5 mg/L, Polimixina B x 500 000 unidades (APP Pharmaceuticals, LLC) 0,66 mg/L, Vancomicina 500 mg (Vitalis) 10 mg/L y Trimetroprim sulfa 40/200 mg/5 mL MK® 5 mg/L. Los cultivos se incubaron a 37 °C por un periodo de 7 a 10 días, en cámara con generador de anaerobiosis Gas PakTM EZ Anaerobe Container system with Indicator (Becton and Dickinson, Company, USA). A las colonias obtenidas después del periodo de incubación se les realizó coloración de Gram y las pruebas de catalasa, ureasa y oxidasa para confirmar el fenotipo de la bacteria.
Extracción de ADN genómico. La extracción del ADN genómico de los aislados de H. pylori se realizó de acuerdo con el protocolo descrito por Valentine.14 Las bacterias fueron resuspendidas en buffer TE 1X (10 mM Tris-HCl pH 7,5; 1 mM EDTA pH 8,0), 0,5 % SDS (Dodecil sulfato de sodio), y Proteinasa K (20 mg/mL), después fueron incubadas 1 h a 37 °C. Posteriormente fueron tratadas con NaCl 5 M y solución 10 % CTAB (bromuro de hexadeciltrimetilamonio)/NaCl 0,7 M por 15 min a 65 °C; por último el material genético (ADN) fue purificado con cloroformo/alcohol isoamílico (24:1), y resuspendido en 100 µL de buffer TE 1X.
Para verificar la calidad del material genético extraído se realizó electroforesis en gel de agarosa al 0,8 % (p/v) teñidos con Gel RedTM15 y visualizados en un transiluminador de luz UV ENDURO™ GDS (Labnet).
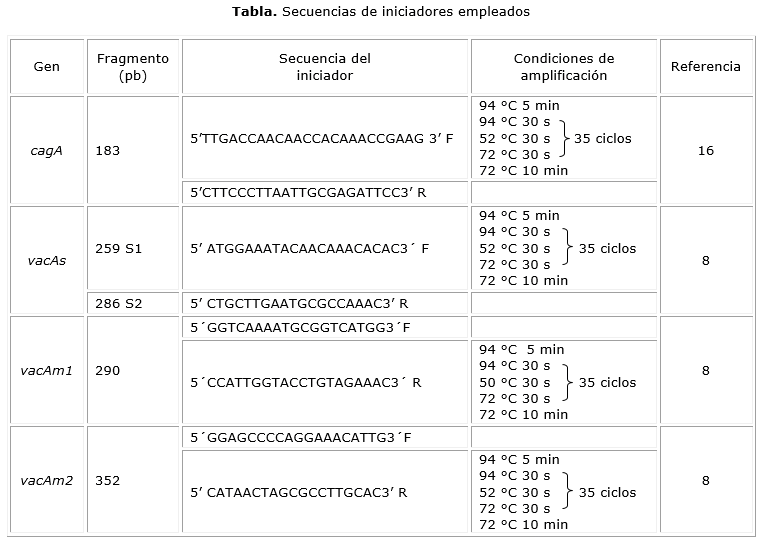
Reacción en cadena de la polimerasa (PCR). El ADN genómico obtenido fue empeado para la amplificación de fragmentos asociados a los genes citotóxicos vacA y cagA (tabla). Se incluyó como control positivo muestras de cada uno de los genes en estudio que previamente fueron amplificadas por PCR, secuenciadas, analizadas y verificadas para cada uno de los alelos. La mezcla de reacción contenía: 1X buffer (Bioline), 2 mM MgCl2 (Bioline), 2,5 mM dNTP (Bioline), 2,5 µM de cada uno de los iniciadores, 0,5 U de la enzima Taq polimerasa (Bioline) y 50-100 ng de ADN. La amplificación se realizó en el termociclador Multigene (LabnetTM). Los productos obtenidos fueron corridos en geles de agarosa al 1,5 % (p/v) teñido con Gel RedTM y visualizados en el transiluminador de luz UV ENDURO™ GDS (Labnet).
Los datos fueron sometidos a procedimientos estadísticos descriptivos y relacionales. La plataforma utilizada para el análisis de datos fue el programa SPSS (Statistical Package for the Social Sciences) versión 24, de la empresa IBM. Se consideró estadísticamente significativo un valor p (α)< 0,05. Los estadígrafos utilizados fueron chi cuadrado con corrección de continuidad de Yates y chi cuadrado de independencia para la comparación de la prevalencia observada frente a la esperada. Se desarrollaron análisis complementarios de sensibilidad y especificidad mediante tablas de contingencia.
RESULTADOS
De los 72 pacientes incluidos en el estudio, 28 pacientes (38,9 %) correspondieron a hombres y 44 pacientes (61,1 %) a mujeres, con un promedio de edad de 43,2 años (rango 18-73 años).
A todos los pacientes se les realizó cultivo microbiológico de biopsias obtenidas del antro y cuerpo del estómago, se aisló H. pylori en 16 pacientes de los cuales siete presentaron crecimiento en antro, cuatro en cuerpo y cinco en antro y cuerpo del estómago.
Basados en los diagnósticos endoscópicos de los pacientes con aislamiento de H. pylori, se encontró una frecuencia correspondiente a: 3/16 (18,85 %) para gastritis crónica antral; 2/16 (12,5 %) para gastritis erosiva antral y gastritis nodular antral y 1/16 (6,3 %) para cada uno de los siguientes diagnósticos: gastritis atrófica con metaplasia intestinal, gastritis verrucosa antral, gastritis atrófica corporoantral y gastropatía antral.
Para el gen cagA y las variantes alélicas del gen vacA (Fig.), el tamaño de los fragmentos amplificados correspondió a aquellos previamente informados por otros autores.8,16 Con la identificación molecular se encontraron las siguientes frecuencias: cagA+ 8/16 (50 %), cagA- 8/16 (50 %), vacAs1 12/16 (75 %), vacAs2 4/16 (25 %), vacAm1 14/16 (87,5 %), vacAm2 2/16 (12,5 %).
Las variantes alélicas identificadas para vacA fueron: vacAs1m1 11/16 (68,8 %), vacAs1m2 1/16 (6,3 %), vacAs2m1 3/16 (18,8 %), vacAs2m2 1/16 (6,3 %). La asociación entre el genotipo cagA positivo o negativo con las variantes alélicas de vacA no mostró relación estadísticamente significativa (p> 0,05). De manera similar, no se encontró relación estadísticamente significativa entre el diagnóstico endoscópico, las variantes alélicas de vacA (p> 0,05) y el genotipo de cagA (p > 0,05).
DISCUSIÓN
El desarrollo clínico de la infección por H. pylori está asociado a factores de virulencia, ambientales y del hospedero,17,18 razón por la cual el diagnóstico y el tratamiento de la infección juegan un papel fundamental en la erradicación del patógeno.19
Los métodos de diagnóstico de H. pylori como las pruebas histológicas, cultivo microbiológico, reacción en cadena de la polimerasa y prueba rápida de ureasa, requieren la toma de biopsias por endoscopia y por lo tanto son consideradas como métodos invasivos.20 El cultivo es considerado el método más específico para la detección de H. pylori, es empleado en la práctica clínica para confirmar sensibilidad frente a los antibióticos cuando se presenta falla del tratamiento y en investigación los aislamientos de H. pylori son usados para la identificación de factores de virulencia y para estudios de epidemiología.20 Es este estudio el cultivo microbiológico se realizó para la identificación molecular de los genotipos cagA y variantes alélicas de vacA; se consiguieron aislamientos de H. pylori en 16 pacientes, la baja proporción de aislamientos obtenidos pudo estar asociado al nivel socioeconómico y la procedencia de los pacientes, fundamentalmente debido a que la mayoría residían en la zona urbana del municipio Manizales y pertenecían a estratos socioeconómicos medio y medio alto, donde la calidad de vida está relacionada entre otros factores, con la calidad del agua a la vez que se garantiza optimas condiciones de higiene, pues se ha informado un mayor riesgo de infección por H. pylori en personas que habitan en el área rural donde normalmente los estratos socioeconómicos son bajos, las condiciones sanitarias no son adecuadas y el agua de consumo humano es de baja calidad.21,22
A nivel molecular, la reacción en cadena de la polimerasa, con una sensibilidad del 75 % y una especificidad del 90 %, es considerada la prueba estándar para el diagnóstico de la infección, basada en el uso de iniciadores específicos permite la identificación de factores de virulencia y los cambios génicos que conducen a la resistencia a antibióticos.7,9,19,23-25
Para las cepas cagA+ se han informado prevalencias de 74,8 % en Ghana, 90 % en Nigeria, 95 % en Sudáfrica,24 100 % en Japón25 y 47,8 % en Brasil26,28 entre otros. En el caso de Colombia la prevalencia se encuentra entre el 43 % y el 90,5 %.9,25,27,29 En este estudio se encontró una prevalencia de cagA+ del 50 %, ubicándose en el rango informado en los estudios colombianos (p> 0,05); sin embargo, y en concordancia con otras investigaciones, no se encontró una asociación significativa entre las cepas cagA+ y el diagnóstico establecido de enfermedad gastroduodenal.30,31
Con respecto al factor de virulencia vacA, los resultados mostraron mayor predominio de vacAs1 (75 %) y vacAm1 (87,5 %), estos resultados son consistentes con lo informado por varios autores.4,9,29,32 En la combinación de los genotipos de la región s y m, la variante vacAs1m1 fue identificada en el 68,8 % de los aislamientos, guardando relación con las frecuencias entre el 24 % y el 84 % indicadas en diferentes poblaciones a nivel mundial.30
Los resultados no mostraron asociación significativa entre el genotipo cagA+ y las combinaciones alélicas de vacA y el diagnóstico endoscópico, lo que podría indicar que factores de virulencia diferentes a los analizados, así como los factores ambientales y propios del hospedero podrían estar relacionados con el desarrollo de la enfermedad gastroduodenal.17,18
Hubo una prevalencia del 50 % de cagA+ y una frecuencia 68,8 % para las variantes alélicas vacAs1m1 en los aislamientos de H. pylori obtenidos de las biopsias gástricas de pacientes que fueron remitidos a la clínica San Marcel, lo que denota que factores diferentes como los de tipo medioambiental o del hospedero, complementan la virulencia bacteriana, asociándose a la presentación de condiciones gastrointestinales con implicaciones diagnósticas, pronósticas y terapéuticas.
Es necesario realizar estudios que permitan el aislamiento de un número mayor de cepas de H. pylori para determinar la influencia de diferentes factores de virulencia con el desarrollo clínico de la enfermedad gastroduodenal.
Agradecimientos
Los investigadores agradecen a la dirección de investigaciones de la Universidad de Manizales por la financiación del proyecto Análisis clínico, microbiológico y molecular de Helicobacter pylori en pacientes con lesiones gástricas en el departamento de Caldas. Código E0601X0204.
Conflicto de intereses
Los autores declaran que no presentan conflicto de interés.
REFERENCIAS BIBLIOGRÁFICAS
1. Khalifehgholi M, Shamsipour F, Ajhdarkosh H, Daryani NE, Pourmand MR, Hosseini M, et al. Comparison of five diagnostic methods for Helicobacter pylori. Iran J Microbiol. 2013;5(4):396-401
2. Arévalo A, Trespalacios A, Otero W. The importance of CagA protein in Helicobacter Pylori infection. Rev Colomb Gastroenterol. 2009;24(4):388-95.
3. Dabiri H, Jafari F, Baghaei K, Shokrzadeh L, Abdi S, Pourhoseingholi MA, et al. Prevalence of Helicobacter pylori vacA, cagA, cagE, oipA, iceA, babA2 and babB genotypes in Iranian dyspeptic patients. Microb Pathog. 2017;105:226-30.
4. González-Vázquez R, Córdova-Espinoza MG, Escamilla-Gutiérrez A, Morales-Méndez I, Ochoa-Pérez SA, Armendáriz-Toledano F, et al. Frequency of virulence genes in mixed infections with Helicobacter pylori strains from a Mexican population. Rev Gastroenterol Méx. 2016;81(1):11-20.
5. Salimzadeh L, Bagheri N, Zamanzad B, Azadegan-Dehkordi F, Rahimian G, Hashemzadeh-Chaleshtori M, et al. Frequency of virulence factors in Helicobacter pylori-infected patients with gastritis. Microb Pathog. 2015;80:67-72.
6. Audibert C, Burucoa C, Janvier B, Fauchere JL. Implication of the Structure of the Helicobacter pylori cag Pathogenicity Island in Induction of Interleukin-8 Secretion. Infect Immun. 2001;69(3):1625-1629.
7. Breurec S, Michel R, Seck A, Brisse S, Come D, Dieye F, et al. Clinical relevance of cagA and vacA gene polymorphisms in Helicobacter pylori isolates from Senegalese patients. Clin Microbiol Infec. 2012;18(29):153-9.
9. Arévalo-Galvis A, Trespalacios-Rangel AA, Otero W, Mercado-Reyes MM, Poutou-Piñales RA. Prevalence of cagA, vacA, babA2 and iceA Genes in H. pylori Strains Isolated from Colombian Patients with Functional Dyspepsia. Pol J Microbiol. 2012;61(1):33-40.
10. Mnich E, Kowalewicz-Kulbat M, Sicińska P, Hinc K, Obuchowski M, Gajewski A, et al. Impact of Helicobacter pylori on the healing process of the gastric barrier. World J Gastroenterol. 2016;22(33):7536.
11. Backert S, Blaser M. The Role of CagA in the Gastric Biology of Helicobacter pylori. Cancer Res. 2016;76(14):4028-31.
12. Correa P. Gastric Cancer: Overview. Gastroenterol Clin N Am. 2013;42:211-7.
13. Bravo LE, Cortes A, Carrascal E, Jaramillo R, Garcia LS, Bravo PE, et al. Helicobacter pylori: patología y prevalencia en biopsias gástricas en Colombia. Colomb Med. 2003;34:124-31.
14. Valentine Jl, Arthur RR, Mobley HL, Dick J. Detection of Helicobacter pylori by using the polymerasa chain reaction. J Clin Microbiol. 1991 Apr;29(4):689-95.
15. Uribe P, Herrera J, Orozco N, Betancur J. Uso alternativo del colorante gelred en la tinción de ácidos nucleicos. Archivos de Medicina. 2013;13(2):160-6.
16. Trespalacios A, Otero R, Mercado M. Resistencia de Helicobacter pylori a metronidazol, claritromicina y amoxicilina en pacientes colombianos. Rev Colomb Gastroenterol. 2010;25(1):31-8.
17. Shiota S, Suzuki R, Matsuo Y, Miftahussurur M, Tran TT, Binh T, et al. Helicobacter pylori from Gastric Cancer and Duodenal Ulcer Show Same Phylogeographic Origin in the Andean Region in Colombia. Plos One. 2014;9(8):105392.
18. Trang TH, Thanh BT, Yamaoka Y. Relationship between vacA Types and Development of Gastroduodenal Diseases. Toxins. 2016;8(6):182.
19. Montealegre C, Jaramillo CA, Montealegre G, Parra G, Echeverry MM, Delgado MP. Detección histológica y molecular de Helicobacter pylori y genotipificación con base en los genes de virulencia babA2 e iceA en pacientes con patología gástrica benigna. Rev Chil. Infectol. 2010;27(2):112-8.
20. Garza-González E, Perez-Perez GI, Maldonado-Garza H, Bosques-Padilla FJ. A review of Helicobacter pylori diagnosis, treatment, and methods to detect eradication. World J Gastroenterol. 2014;20(6):1438-49.
21. Chen H, Ming-Jen C, Shou-Chuan S. Socioeconomic status, personal habits, and prevalence of Helicobacter pylori infection in the inhabitants of Lanyu. J Formos Med Assoc. 2014;113(5): 278-83.
22. Correa S, Cardona AF, Correa T, Correa LA, García HI, Estrada S. Prevalencia de Helicobacter pylori y características histopatológicas en biopsias gástricas de pacientes con síntomas dispépticos en un centro de referencia de Medellín. Rev Col Gastroenterol. 2016;31(1):9-15.
23. Moncayo JI, Álvarez A, Santacruz JJ, Santacoloma M, Arturo B, et al. Evaluación de diferentes pruebas para el diagnóstico de Helicobacter pylori. Investigaciones Andina 2011;13(23):297-311.
24. Patel S. Diagnosis of Helicobacter pylori: What should be the gold standard? World J Gastroenterol. 2014;20(36):12847.
25. Molina AG, Jaramillo CA, Delgado MP, Bohórquez ME, Ámezquita A. Detección y genotipificación de Helicobacter pylori sobre la base de los genes ADNr 16S y el gen asociado a citotoxina (cagA) y posible asociación con enfermedades gastrointestinales. Rev Cubana Med Trop. 2008;60(2):105-10.
26. Archampong T, Asmah RH, Aidoo EK, Wiredu EK, Gyasi RK, Adjei DN, et al. Helicobacter pylori cagA and vacA genes in dyspeptic Ghanaian patients. BMC Res Notes. 2017;10:231.
27. Watada M, Shiota S, Matsunari O, Suzuki R, Murakami K, Fujioka T, et al. Association between Helicobacter pylori cagA-related genes and clinical outcomes in Colombia and Japan. BMC Gastroenterol. 2011;11:141.
28. Pereira WN, Avante M, Munhoz L, De Labio RW, Aparecido W, Bianchi JP, et al. Association among H. pylori virulence markers dupA, cagA and vacA in Brazilian patients. J Venom Anim Toxins Incl Trop Dis. 2014;20:1
29. Moncayo JI, Santacruz JJ. Deteccion del gen CagA y tipificación del gen VacA en cepas de Helicobacter pylori aisladas de pacientes con enfermedad ulcero péptica en Risaralda. MedUNAB. 2000;3(8):69-75.
30. Zhou W, Yamazaki S, Yamakawa A, Ohtani M, Ito Y, Keida Y. The diversity of vacA and cagA genes of Helicobacter pylori in East Asia. FEMS Immunol Med Microbiol. 2004;40:81-7.
31. Pajavand H, Alvandi A, Mohajeri P, Bakhtyari S, Bashiri H, Kalali B, et al. High Frequency of vacA s1m2 Genotypes Among Helicobacter pylori Isolates From Patients With Gastroduodenal Disorders in Kermanshah, Iran. Jundishapur J Microbiol. 2015;8(11):e25425.
32. Mendoza-Elizalde S, Cortés-Márqueza AC, Giono-Cerezo S, Zuñiga G, Consuelo-Sánchezd A, Valencia-Mayoral P, et al. Analysis of the genotypic diversity of strains of Helicobacter pylori isolated from pediatric patients in Mexico. Infect Genet Evol. 20015;29:68-74.
Recibido: 2 de octubre de 2017.
Aprobado: 23 deagosto de 2018.
Paula Tatiana Uribe Echeverry. Grupo de investigación Médica. Universidad de Manizales. Manizales, Caldas, Colombia. Correo electrónico: puribe@umanizales.edu.co


{kind=link}
{kind=link}