










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
Las especies exóticas invasoras constituyen una amenaza para la biodiversidad de los ecosistemas naturales y para la agricultura mundial. El incremento del comercio internacional, el turismo y el deterioro progresivo del clima han traído como consecuencia la diseminación de diferentes agentes nocivos desde su centro de origen hacia otras latitudes, eludiendo las barreras naturales establecidas como parte del control natural. Dentro de las ventajas que muestran las mismas sobre las especies nativas están la alta supervivencia, el rápido crecimiento poblacional, la adaptación a las nuevas condiciones climáticas, los mecanismos de dispersión más efectivos y la ausencia de sus enemigos naturales, los que pudieran suprimir el desarrollo de sus poblaciones cuando invaden nuevos territorios (1).
El ácaro rojo del cocotero (Raoiella indica Hirst) es una especie exótica invasora que se encuentra comúnmente en muchas especies de arecáceas (2,3). En Cuba ha mostrado preferencia por Cocos nucifera L. (4), sobre el cual alcanza altas densidades poblacionales (5).
Los estudios de los parámetros biológicos y poblacionales de R. indica efectuados en varios países han mostrado que el crecimiento de los parámetros poblacionales, así como su tasa de desarrollo, longevidad y fecundidad, dependen de las condiciones ambientales y las plantas hospedantes (5,6,7). Estos resultados imponen la necesidad de conocer su comportamiento biológico en las condiciones ambientales imperantes en nuestro país, las cuales son idóneas para el desarrollo de dicho ácaro. A partir de lo antes expuesto, el objetivo de nuestro trabajo fue determinar la biología y la tabla de vida de R. indica sobre hojas de C. nucifera.
MATERIALES Y MÉTODOS
El experimento se efectuó en el Laboratorio de Fitopatología del Centro de Estudio de Tecnologías Agropecuaria (CETA) perteneciente a la Facultad Agroforestal (FAF), de la Universidad de Guantánamo (UG). Durante la realización del experimento la temperatura promedio fue de 30,8 ± 1,5 ºC y la humedad relativa de 56,7 ± 8,2 %, registradas con un Termohigrómetro digital (Testo 608-H2).
A partir de poblaciones de R. indica establecidas sobre hojas de C. nucifera cv. ´Criollo` (Indio verde) en condiciones de campo, se estableció una cría en laboratorio por el método de supervivencia de hojas en placas Petri de 10,5 cm de diámetro.
Para iniciar el estudio de duración del desarrollo, se tomaron cinco hembras grávidas de la cría establecida en laboratorio con un pincel 00 y se individualizaron en unidades experimentales constituidas por porciones de hojas de cocotero cv. ´Criollo` (Indio verde) de 3 cm2 colocadas en placas Petri, cuyos bordes se cubrieron con algodón humedecido para evitar que los ácaros se escapen y mantener la turgencia. Las hembras se observaron cada 12 horas y cuando se registró la presencia de huevos, las mismas se retiraron y se dejó solo un huevo por unidad experimental.
Las unidades experimentales se revisaron cada 12 horas y se registraron los cambios de fase y la duración de los mismos hasta la emergencia de los adultos. Cada vez que se produjo un cambio de fase, se realizó un cambio de sustrato. Con los datos obtenidos se calculó la duración media del desarrollo, la desviación estándar y los valores máximos y mínimos absolutos, por fase y de huevo hasta adulto, en días, para ambos sexos y total.
Las hembras obtenidas del estudio del desarrollo se mantuvieron individualizadas en las unidades experimentales y se les colocó un macho de la cría durante 24 horas para garantizar la primera cópula. Una vez que se confirmó el apareamiento, se observaron diariamente hasta la muerte de las hembras. Se registró el número de huevos puestos y la viabilidad de los mismos; se consideraron no viables aquellos huevos que no eclosionaron en un término de 15 días.
A partir de estos datos se determinó la duración promedio de los periodos de preoviposición, oviposición, posoviposición y la longevidad de las hembras. Además, se determinó la curva de supervivencia (% de hembras vivas.día-1), la fecundidad promedio y la tasa de oviposición diaria (huevos.hembra.día-1).
Para la determinación de los parámetros de la tabla de vida de R. indica se utilizaron los datos recopilados del estudio de la biología. Sobre la base del procedimiento descrito por Birch (8), se calcularon los siguientes parámetros poblacionales:
Tasa neta de reproducción (R 0 ). Refleja el número promedio de progenie hembra que es capaz de producir cada hembra de la población durante toda su vida.

Cuando el valor R 0 es igual a 1, entonces la población se reemplazará a sí misma de manera precisa, de generación en generación; si R 0 > 1 la población estará en un estado de crecimiento, si R 0 < 1 la población estará decreciendo (9).
Tiempo medio de generación (t). Representa el tiempo promedio entre dos generaciones sucesivas.
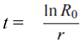
Tasa intrínseca de incremento (r m ). Número de progenie hembra por hembra por unidad de tiempo.
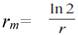
Tasa finita de incremento (λ). La tasa intrínseca de incremento natural se puede convertir a una tasa finita de multiplicación (por individuo, por unidad de tiempo) y se interpreta como el número de individuos que se agrega a la población por individuo

Tiempo de duplicación de la población (TD). Número de días requeridos por la población para duplicar su número.
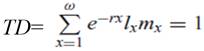
RESULTADOS Y DISCUSIÓN
El ácaro rojo del cocotero tiene cuatro fases de desarrollo (huevo, larva, dos estados ninfales y adulto). Los cambios de fases después de la eclosión del huevo están dados por la presencia de la exuvia o muda. Los huevos, por lo general, se encuentran en grupos y en pocas ocasiones pueden verse de forma aislada. Tienen una coloración anaranjada que se torna blanco opaca cuando están próximos a la eclosión.
Las larvas poseen tres pares de patas, son de forma ovalada con una coloración rojiza, presentan tamaño pequeño y movimientos lentos. Por lo general, permanecen la mayor parte del tiempo alimentándose hasta llegar al estado quiescente, que puede ser de 1-2 días. Acto seguido a la muda, emerge la protoninfa con cuatro pares de patas y un tamaño mayor a la larva. Los movimientos de la protoninfa son más rápidos que los de la fase anterior; se alimenta hasta entrar en el estado quiescente que demora de 1-2,5 días. La deutoninfa es similar a la protoninfa, pero con mayor tamaño.
La hembra tiene el cuerpo redondeado, de color rojizo, el tamaño es mayor al de los machos. Presenta manchas oscuras en la parte dorsal debido a la alimentación. Los machos son más pequeños que las hembras con la parte del opistosoma de forma aguzada.
En todas las fases los ácaros se liberan del exoesqueleto por movimientos que realizan hasta romperlos de formas transversal y quedar libre del anterior exoesqueleto. En los extremos de las setas transversales se pudo observar la presencia de secreciones en forma de gota.
La duración del ciclo de desarrollo fue aproximadamente de 17 días; las hembras necesitaron más tiempo que los machos para alcanzar la adultez (Tabla 1). La fase de larva demoró más para ambos sexos, seguida de los huevos, mientras que la menor duración fue para la protoninfa. Las fases inmaduras tuvieron una baja mortalidad, se encontró 13 % para las larvas, 10,2 % en las protoninfas y solo 2,27 % en las deutoninfas, para un total de 24,98 %. Esto indica que esta plaga puede desarrollarse con facilidad sobre este hospedante y lograr una alta supervivencia. El cociente sexual fue 67 % favorable a las hembras.
TABLA 1 Duración del desarrollo (en días) para todas las fases de Raoiella indica sobre hojas de Cocos nucifera./ Duration (in days) for all the development stages of Raoiella indica on leaves of Cocos nucifera. (Mín: mínimo; Máx: máximo; DE: desviación estándar)
| Fases | Hembra | Macho | Total | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Mín | Máx | Media±DE | Mín | Máx | Media±DE | Mín | Máx | Media±DE | |
| Huevo | 3 | 6,5 | 4,73±0,99 | 3 | 6,5 | 4,50±1,10 | 3 | 6,5 | 4,64±1,03 |
| Larva | 4 | 7,5 | 5,18±1,06 | 4 | 6 | 5,00±0,92 | 4 | 7,5 | 5,12±1,01 |
| Protoninfa | 2,5 | 6,5 | 3,54±1,11 | 2 | 4 | 2,96±0,67 | 2 | 6,5 | 3,34±1,03 |
| Deutoninfa | 3 | 6,5 | 4,55±1,42 | 3 | 5 | 3,71±0,74 | 3 | 6,5 | 4,27±1,30 |
| Adulto | 13 | 23,5 | 18,00±2,92 | 14,5 | 19 | 16,40±1,42 | 13 | 23,5 | 17,38±2,65 |
| N | 28 | 14 | 42 | ||||||
La duración del desarrollo de R. indica obtenida en este estudio está dentro del rango de duración informada para esta especie en condiciones climáticas similares (29 ± 1,0 °C y 60±10 % de humedad relativa) sobre cuatro cultivares de cocotero: ´Jamaican Tall´, ´MYD x JT´, ´Niu Leka´ y ´Malayan Yellow Dwarf´. En este estudio, la menor duración se observó también para la fase de protoninfa, mientras que la mayor duración se registró para la fase de huevo (10). Sin embargo, fue inferior a la informada para los cultivares ´Salak Dwarf´, ´Local Sumenep Tall´ e ´Hybrid´, aunque en este caso con condiciones ambientales más frescas (26-28 oC y 75-85 % de humedad relativa (11).
Los estudios de duración del desarrollo, parámetros reproductivos y su relación con los factores bióticos y abióticos son elementos fundamentales en el conocimiento de una especie y, por consiguiente, en la toma de decisión para su control, pero con mayor significación cuando se trata de especies exóticas invasoras en el país donde se introdujo (12).
La duración del ciclo de desarrollo en R. indica puede estar influenciada por la calidad del sustrato utilizado y por la especie de planta (13). Otro factor que puede influir también son los cultivares de cocotero utilizadas, en particular sobre el desarrollo de las fases inmaduras (14).
Al analizar los periodos de preoviposición, oviposición y posoviposición (Tabla 2), se observa que la preoviposición tuvo una corta duración; en este periodo las hembras están maduras sexualmente y realizan la cópula con los machos. La oviposición se inicia a partir del segundo día después que emerge la hembra y puede durar hasta los 26 días, con una media de 12,77 días.
TABLA 2 Duración (en días) de los periodos de preoviposición, oviposición, posoviposición y longevidad de Raoiella indica sobre hojas de Cocos nucifera./ Duration (in days) of preoviposition, oviposition, and postoviposition periods and longevity of Raoiella indica on leaves of Cocos nucifera (Mín: mínimo; Máx: máximo; DE: desviación estándar)
| Periodos | Mín | Máx | Media±DE |
|---|---|---|---|
| Preoviposición | 2 | 5 | 2,93±0,86 |
| Oviposición | 2 | 26,5 | 12,77±6,34 |
| Posoviposición | 0,5 | 11,5 | 3,48±2,72 |
| Longevidad | 6 | 36 | 19,09±8,32 |
La longevidad de las hembras llegó hasta los 36 días, con una media de 19,09 días. Estos resultados son similares a lo informado previamente para cocotero por Hastie (15) para Cuba; superiores a los encontrados por Vásquez (12), que solo alcanzó los 24,1 días y Gómez-Moya et al. (13), donde llegaron a los 27,1 días. Esto puede deberse a diferencias en la preferencia del ácaro por los cultivares de cocotero utilizados.
El pico máximo de puesta de huevos ocurre a los cinco días, a partir de este momento disminuye progresivamente hasta dejar de ovipositar a partir del día 35 después del inicio de la puesta (Fig. 1). Con respecto a la media de huevos por hembra por día, se evidencia que en los primeros días es donde se alcanzan los mayores valores y disminuyen según las hembras van envejeciendo.
La curva de supervivencia se corresponde a una curva intermedia de tipo I y II, siguiendo los criterios de Vera (16), con una mortalidad que afecta fundamentalmente en las primeras etapas de vida de la población; se produce posteriormente una mortalidad progresiva, con un máximo aproximado en la mitad de su vida. La mayor cantidad de hembras viven durante el periodo de máxima oviposición y van muriendo progresivamente, con un incremento de la mortalidad después de pasar esta fase.
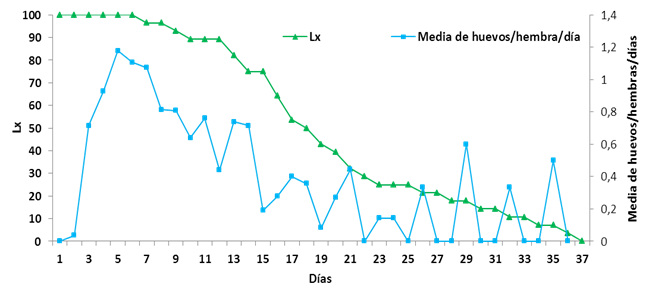
FIGURA 1 Supervivencia (Lx) y media de huevo por hembra por día de Raoiella indica sobre hojas de Cocos nucifera. / Daily oviposition rate and survival of Raoiella indica on leaves of Cocos nucifera
La fecundidad de las hembras varió de 3 a 18 huevos, mínimo y máximo respectivamente, con un promedio de 10,64±3,46 huevos. Esta fecundidad fue inferior a la obtenida por Vásquez (12). Se observó que el mayor promedio de huevos por hembra por día se alcanzó desde el tercer día hasta el día 14 y el pico máximo de oviposición ocurrió al cuarto día con 1,2 huevos por hembra, con una media general de huevos por hembra por días de 0,57, inferior a la informada por Vásquez (12), quien informó una media de 2,4.
De acuerdo con Tabashnik (14), las variaciones en la tasa de oviposición de un organismo pudieran ser parcialmente explicadas por el efecto, tanto de la planta hospedante como por la genética del herbívoro. Sin embargo, estas variaciones han sido comprobadas principalmente entre poblaciones; queda aún sin explicar cómo la genética actúa para explicar las variaciones intrapoblacionales de la oviposición (17).
Como en el estudio solo se colocó un macho por hembra, podría ser este un factor para que la fecundidad sea inferior a la informada por otros autores, si se tiene en cuenta que se conoce que varios machos por unidad experimental pueden garantizar múltiples cópulas y esto garantiza una mayor oviposición en muchas especies de ácaros. Además, la presencia de varios individuos proporciona un efecto de grupo que puede favorecer la oviposición (18), aunque el uso de un solo macho por hembra es muy común en estudios biológicos de ácaros. La eclosión de los huevos alcanzó el 95,3 %.
La curva de mortalidad (dx) logra el valor máximo a los 15 días de emergidas las hembras; este valor se conoce como edad modal para las muertes (Fig. 2). A medida que las causas de mortalidad aumentan, la edad modal disminuye; este es el propósito principal de las acciones de control, según los criterios de Baños (19). Ello implicaría la adopción de alguna medida de manejo o control, con anterioridad a los 15 días, con vista a disminuir la edad modal para la muerte de las hembras.
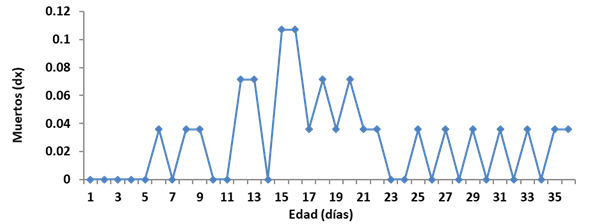
FIGURA 2 Tasa de mortalidad (dx) de las hembras de Raoiella indica sobre hojas de Cocos nucifera. / Mortality rate (d x ) of females of Raoiella indica on leaves of Cocos nucifera
A partir de la estimación de los parámetros poblacionales de R. indica, se pudo observar que la tasa neta de reproducción (R 0 ) fue de 14,03; lo anterior quiere decir que la población está en crecimiento, ya que el valor es superior a uno y que cada hembra puede agregar 14,03 individuos a la población; esto evidencia la importancia que puede llegar a alcanzar esta especie exótica invasora en este cultivo debido a su capacidad reproductiva.
La tasa intrínseca de incremento (r m ) fue de 0,41, por tanto, en condiciones ambientales ideales, esta especie potencialmente puede multiplicar su población de manera acelerada. El valor de r m fue superior al informado por Vásquez (12) y Gómez-Moya et al. (13), con una temperatura media de 29 °C. Según Reyes y Mesa (20), la tasa intrínseca de crecimiento natural es la capacidad de multiplicación de una especie en un momento dado.
La r m es un parámetro genéticamente determinado, que refleja una capacidad potencial de multiplicación poblacional. Aunque esta tasa se calcula en condiciones de laboratorio, es útil para predecir lo que puede pasar en condiciones de campo si estas le son favorables a la población. Este resultado evidencia que R. indica posee el potencial biológico necesario para incrementar rápidamente sus poblaciones sobre cocotero, lo cual resulta de interés para los encargados de la protección fitosanitaria del cultivo.
El número de individuos que se agregaría por hembra por día a la población serían 1,51, según la tasa finita de crecimiento (λ). Esta puede ser una de las razones del porqué en los campos se presentan ataques de esta especie con grandes cantidades de individuos.
El tiempo que media entre dos generaciones sucesivas (t) fue de 12,36 días, con un tiempo de duplicación de la población (TD) de 1,69. Los parámetros poblacionales calculados evidencian la capacidad de duplicación y de agregar hembras de manera permanente que tiene R. indica, lo que permite su persistencia en el cultivo y la multiplicación de su población en condiciones ambientales favorables.
Las tasas netas de reproducción, intrínsecas de incremento y finitas de incremento determinadas para R. indica fueron superiores a las obtenidas para Tetranychus gloveri Banks sobre hojas de C. nucifera (21). También fueron superiores a las obtenidas para Tetranychus mexicanus McGregor, criado sobre otra especie de palmera (Bactris gasipaes Kunth) (22).
Las características biológicas y poblacionales de R. indica en condiciones de laboratorio permiten estimar lo que está sucediendo en la naturaleza de una forma abstracta, debido a la existencia en el medio de factores ambientales variables como la temperatura, la humedad relativa y las precipitaciones, que pueden reducir drásticamente las poblaciones de fitófagos. Sin embargo, esta información unida al conocimiento de sus atributos biológicos ofrecen elementos importantes sobre aspectos necesarios para el manejo de la especie (23).
Los parámetros poblacionales permiten pronosticar la potencialidad de este ácaro como plaga en el cultivo del cocotero. Además, ofrecen elementos que servirán de base en el manejo de esta especie, ya que las tablas de vida son una herramienta que permite cuantificar su crecimiento poblacional. Elementos que hay que tener en cuenta para hacer cualquier tipo de pronóstico sobre este agente nocivo.
El estudio de las tablas de vida constituye una herramienta básica para elaborar estrategias de control de plagas. Estas estimaciones pueden ser utilizadas para evaluar la resistencia varietal ante una plaga y como patrón para seleccionar enemigos naturales eficientes. Por ello, este estudio ofrece elementos que servirán de base en el manejo de R. indica sobre cocotero.

