










 Servicios personalizados
Servicios personalizados Español (pdf)
Español (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
El frijol común (Phaseolus vulgaris L.) es la leguminosa más estudiada en América Latina y es considerado una fuente de alimentación importante por su alto valor nutricional. En nuestro país, junto con el arroz (Oryza sativa L.), conforma la base de la dieta diaria de los ciudadanos y, a nivel mundial, se reconoce como un alimento clave para la seguridad alimentaria (1). La incidencia de diversas plagas, nematodos fitoparásitos y hongos fitopatógenos (particularmente Sclerotium sp., Fusarium spp., Rhizoctonia spp.) produce afectaciones de hasta un 50 % en la producción de este grano (1).
Hasta el presente, el aumento de la productividad del cultivo y el control de plagas, se basan en aplicaciones de fertilizantes y plaguicidas químicos, que son costosos y tóxicos, siendo perjudiciales a la salud humana y al ambiente. Por ello, se necesita la búsqueda e introducción de nuevas alternativas de manejo como el uso de organismos benéficos, que incrementen los niveles productivos del cultivo, así como su calidad, sin ocasionar daños en los agroecosistemas (2).
Las especies del género Trichoderma, se encuentran dentro de las más estudiadas, debido a su elevada capacidad reproductiva, plasticidad ecológica y mecanismos de acción directos e indirectos (3). Dichos mecanismos le proporcionan a la planta mejor adaptabilidad en el medio, además de su acción como inductor de resistencia sistémica a diferentes patógenos (4). Algunos autores informaron la presencia del hongo colonizando la rizósfera de diversos cultivos y estimulando su crecimiento (5).
El cepario del laboratorio de Micología Vegetal del Centro Nacional de Sanidad Agropecuaria (CENSA) dispone de cepas de Trichoderma asperellum Samuels, Lieckfeldt & Nirenberg que están caracterizadas morfo-fisiológica, patogénica y molecularmente (2,6). Sin embargo, se desconoce la capacidad endofítica y el efecto como estimuladoras del crecimiento de estas cepas en el cultivo del frijol. Por tal motivo, el objetivo de este trabajo fue evaluar el efecto endofítico y estimulante de cepas seleccionadas de T. asperellum en el cultivar comercial de frijol 'BAT-304', en condiciones semicontroladas.
MATERIALES Y MÉTODOS
El experimento se realizó en una casa de vegetación del CENSA (22°59'29.1"N 82°09'12.3"W), a temperatura de 25±2ºC, humedad relativa entre el 80-85 % y fotoperiodo natura). Se usaron macetas que contenían 400 g de suelo (ferralítico rojo típico) y abono orgánico en proporción 3:1 (v/v), esterilizado por un periodo de 30 min en autoclave a 120oC, repitiendo la operación a las 24 horas.
Las cepas Ta. 13, Ta. 78, Ta. 85 y Ta. 90 de T. asperellum (Laboratorio de Micología Vegetal, Centro Nacional de Sanidad Agropecuaria, CENSA) se sembraron previamente en placas Petri de 9 cm de diámetro (Ø) contentivas de Agar Malta (AM) (BIOCEN), e incubadas a una temperatura de 28ºC±2ºC y oscuridad.
Los inóculos de las cepas de T. asperellum (Ta. 13, Ta. 78. Ta. 85 y Ta. 90), se prepararon en forma de suspensión, a partir de los cultivos en Papa Dextrosa Agar (PDA) (BIOCEN) de cinco días a una concentración de 107 conidios.ml-1.
Para la siembra se utilizaron semillas de P. vulgaris cultivar 'BAT-304', suministradas gentilmente por el Instituto de Granos, Cuba con un 100 % de viabilidad, las cuales se sembraron en las macetas previamente inoculadas con las diferentes cepas del biorregulador.
Se evaluaron dos momentos de siembra: 1) las semillas se sembraron inmediatamente después de la inoculación del suelo con el hongo y 2) la siembra de las semillas se realizó siete días después de la inoculación del hongo (T. asperellum). Las macetas destinadas a las plantas control no se inocularon. Se utilizó un diseño experimental completamente aleatorizado. Para todos los análisis se realizaron tres réplicas y tres repeticiones para cada tratamiento y controles.
Durante el experimento, las plantas se regaron en días alternos con 20 ml de agua potable. Transcurridos 15 días posteriores a la brotación de cada momento de siembra, las plantas se retiraron de las macetas y se les midieron los siguientes indicadores: altura de la planta, largo de la raíz, número de hojas y raíces, masa fresca foliar y radicular, masa seca foliar y radicular.
Para evaluar la colonización endofítica de las cepas de T. asperellum, las muestras de raíces se lavaron cuidadosamente y se secaron con papel de filtro Whatman No. 4. Las raíces se desinfectaron con hipoclorito de sodio al 1 % durante 30 s y posteriormente con alcohol al 70 %. Entre cada desinfección, las muestras se lavaron con agua destilada estéril tres veces. Seguidamente se cortaron en secciones de 1 cm y se homogenizaron. Por último, se seleccionaron 12 segmentos al azar y se sembraron en placas Petri de 9 cm (Ø), contentivas con PDA, y se incubaron a 28ºC±2ºC y oscuridad. Las determinaciones se realizaron por triplicado. Las evaluaciones se realizaron hasta la aparición de las estructuras del hongo.
La determinación del contenido de nitrógeno (N), fósforo (P) y potasio (K) se realizó según Paneque et al. (7), a partir de material foliar seco de cada tratamiento. Los resultados de los contenidos foliares se expresaron en porcentaje en base a masa seca.
Las actividades enzimáticas específicas peroxidasa (POD), polifenoloxidasa (PPO), fenilalanina amonio liasa (PAL), así como la detección de isoenzimas POD y PPO se analizaron según métodos descritos por Peteira et al. (8). Los datos se compararon a través de un Análisis de Varianza Bifactorial y la Prueba LSD Fisher (α=0.10) usando el programa InfoStat versión 2017 (9).
RESULTADOS Y DISCUSIÓN
A pesar de que las especies de Trichoderma son generalmente usadas como agentes de control biológico contra hongos fitopatógenos, algunas cepas son capaces de producir metabolitos que incrementan el crecimiento vegetal (10). En general, para ambos momentos de siembra, los tratamientos indujeron incremento significativo en la germinación con respecto al testigo. Todas las plantas inoculadas con las diferentes cepas de T. asperellum germinaron al tercer día de la siembra y al momento de la evaluación, las plantas de frijol mostraron evidencias de estimulación del crecimiento. Por su parte, las plantas control lo hicieron al quinto día de la siembra. Estos resultados coinciden con los de Toghueo et al. (11), quienes informaron que T. atroviridae tiene un alto potencial para promover la germinación de semillas de frijol (100 %) y los de Tančić et al. (12) quienes observaron que nueve aislamientos de Trichoderma mejoraron la germinación de las semillas de soya, en comparación con el control.
Solo la cepa Ta. 78 produjo aumento en el número de hojas trifoliadas con respecto a las plantas no tratadas durante el segundo momento de siembra (Tabla 1). Ta. 78, Ta. 85 y Ta. 90 incrementaron el número de raíces con respecto al testigo, cuando la siembra se realizó al unísono con la inoculación del hongo; mientras que, Ta. 78 produjo este efecto cuando la siembra se realizó siete días después de la inoculación.
La aplicación de las esporas de Ta. 13 y Ta. 85 produjo cambios en la altura de las plantas inoculadas durante el primer momento de siembra. Solo Ta. 85 incrementó la longitud de la raíz con respecto a las plantas control. No se encontraron efectos positivos de las cepas en el resto de los indicadores con respecto a las plantas no tratadas.
Tabla 1 Efecto sobre los indicadores del crecimiento y desarrollo en P. vulgaris cv. BAT-304 por la aplicación de T. asperellum. Medias con una letra común no son significativamente diferentes (p > 0.10) / Effect of soil inoculation with T. asperellum on growth and development indicators of in P. vulgaris (Cv. BAT-304). Media with a common letter are not significantly different (p> 0.10)
| Momento de siembra | Tratamiento | No. de Hojas | No. de Raíces | Altura planta (cm) | Largo de la raíz (cm) | Masa Fresca Foliar (g) | Masa Fresca Radicular (g) | Masa Seca Foliar (g) | Masa Seca Radicular (g) |
|---|---|---|---|---|---|---|---|---|---|
| 13 | 9,63 ± 1,45 b | 11,88 ± 0,78 abcd | 16,96 ± 0,96 d | 4,08 ± 0,21 ab | 1,23 ± 0,14 bc | 0,51 ± 0,05 ab | 0,19 ± 0,08 ab | ||
| 78 | 8,5 ± 1,45 b | 79,29 ± 4,55 bc | 19,55 ± 0,96 ab | 3,59 ± 0,21 cde | 1,69 ± 0,14 a | 0,42 ± 0,05 b | 0,27 ± 0,08 ab | ||
| 85 | 8,08 ± 1,45 b | 4,3 ± 0,21 a | 1,1 ±0,14 bc | 0,48 ± 0,05 ab | 0,25 ± 0,08 ab | ||||
| 90 | 8,43 ± 1,48 b | 83,02 ± 4,65 abc | 19,68 ± 0,98 ab | 3,71 ± 0,22 bcde | 0,92 ± 0,14 c | 0,44 ± 0,05 ab | 0,23 ± 0,08 ab | ||
| T | 9,74 ± 1.48 b | 10,09 ± 0,8 d | 72,78 ± 4,65 c | 17,97 ± 0,98 bcd | 3,92 ± 0,22 abc | 1,36 ± 0,15 ab | 0,54 ± 0,05 a | 0,36 ± 0,08 a | |
| 13 | 9,54 ± 1,45 b | 8,17 ± 0,78 e | 80,13 ± 4,55 bc | 15,91 ± 0,96 d | 3,28 ± 0,21 e | 1,25 ± 0,14 bc | 0,47 ± 0,05 ab | 0,24 ± 0,08 ab | |
| 78 | 82,86 ± 4,55 abc | 18,95 ± 0,96 abc | 3,78 ± 0,21 bcd | 1,33 ± 0,14 b | 0,46 ± 0,05 ab | 0,32 ± 0,08 a | |||
| 85 | 9,63 ± 1,45 b | 11,08 ± 0,78 bcd | 78,55 ± 4,55 bc | 19,93 ± 0,96 ab | 3,78 ± 0,21 bcd | 1,24 ± 0,14 bc | 0,5 ± 0,05 ab | 0,31 ± 0,08 ab | |
| 90 | 10,13 ± 1,45 ab | 10,75 ± 0,78 cd | 80,97 ± 4,55 abc | 19,41 ± 0,96 ab | 3,42 ± 0,21 de | 1,1 ± 0,14 bc | 0,48 ± 0,05 ab | 0,1 ± 0,08 b | |
| T | 9,74 ± 1.48 b | 10,09 ± 0,8 d | 72,78 ± 4,65 c | 17,97 ± 0,98 bcd | 3,92 ± 0,22 abc | 1,36 ± 0,15 ab | 0,54 ± 0,05 a | 0,36 ± 0,08 a |
Estos resultados coinciden con los presentados por Hoyos-Carvajal et al. (10) en un trabajo previo donde se evaluó la habilidad de 101 cepas de Trichoderma spp. (entre las que se incluyeron cepas de T. asperellum) para estimular el crecimiento de la planta y se mostró que los hongos de este género pueden producir efectos positivos, nulos e incluso negativos sobre la longitud de la parte aérea, la longitud de las raíces y la masa seca de las plantas de frijol en estadios tempranos. Los resultados de este estudio coinciden con los expuestos por Chowdappa et al. (13), quienes observaron que la cepa OTPB3 de Trichoderma harzianum Rifai aumentó significativamente el crecimiento de raíces y brotes, el área de la hoja y el índice de vigor de plántulas en el tomate (Solanum lycopersicum L.) cuando se evaluó su efecto sobre la estimulación del crecimiento in vitro.
La respuesta diferencial de las plantas a la acción de T. asperellum dependió del momento de siembra del frijol. Los resultados sugieren que durante la germinación de las esporas podría producirse la liberación de fitohormonas y/o metabolitos estimuladores de crecimiento y desarrollo de la planta, de forma más rápida, que cuando el hongo se aplica en la fase de micelio. En un escenario natural, Trichoderma produce metabolitos secundarios que regulan el crecimiento y desarrollo de las plantas o que afectan la proliferación de microorganismos fitopatógenos en el suelo debido a su producción de antibióticos o sideróforos (14).
En los tejidos vegetales se encontraron cambios considerables en el contenido de fitohormonas después de la inoculación de Trichoderma spp. En el caso de plantas de melón (Cucumis melo L.) inoculadas con T. harzianum, se observaron aumentos significativos en los contenidos de zeatina, ácido indol-3-acético (AIA), ácido abscísico (ABA), ácido 1-aminociclopropano-1-carboxílico (ACC), un precursor de etileno (ET), así como de ácido jasmónico (JA) y ácido salicílico (SA) en el brote (14). Martínez-Medina et al. (15) señalaron que existe una fuerte asociación de la inducción de auxinas con la estimulación del crecimiento de las plantas por Trichoderma, principalmente el AIA, las que estimulan la germinación, el crecimiento y el desarrollo radicular y mejoran la asimilación de nutrientes. Otro regulador importante del crecimiento y desarrollo de las plantas es el ET. Este actúa también como una importante molécula de señalización en las plantas bajo ataque de patógenos, lo que resulta en la inhibición del crecimiento de las plantas (16). Se sabe que algunos aislamientos de Trichoderma producen ACC desaminasa, lo que reduce la disponibilidad del ACC necesario para la biosíntesis de ET, lo que podría resultar en el crecimiento de la raíz de la planta (17).
Entre los posibles mecanismos usados por Trichoderma spp. que mejoran el crecimiento vegetal, se encuentra la activación de H+-ATPasa de membrana plasmática (PM), una enzima clave involucrada en la promoción del crecimiento celular y en el alargamiento de la planta. López-Coria et al. (18) evaluaron la participación de la H+-ATPasa PM en el alargamiento inducido por T. asperellum y lo compararon con el efecto del AIA 10 µM porque este también participa en la activación de H+-ATPasa PM. Las semillas tratadas con T. asperellum produjeron plántulas de mayor porte que las plántulas control, a pesar de que estas también acumularon AIA y aumentaron la acidificación de la raíz lo que sugiere que, además de AIA, T. asperellum excreta otras moléculas que estimulan la H+-ATPasa PM para inducir el crecimiento de la planta.
Las cuatro cepas de T. asperellum (Ta. 13, Ta. 78, Ta. 85, Ta. 90) usadas en el estudio mostraron capacidad endofítica de las raíces de frijol en ambos momentos de siembra (Figura 1). Varios estudios revelaron el potencial de los hongos endófitos para la producción de frijol común, especialmente Trichoderma spp.; entre sus ventajas se encuentran la estimulación del crecimiento, la inducción de mecanismos defensivos o el efecto antagónico directo sobre la plaga diana (10,17,19). Por ejemplo, Trichoderma atroviridae Karsten endofítico podría usarse para promover el crecimiento en frijol, para mejorar la productividad de los cultivos y para controlar la enfermedad de la pudrición de la raíz producida por Fusarium solani (Mart.) Sacc. (11).
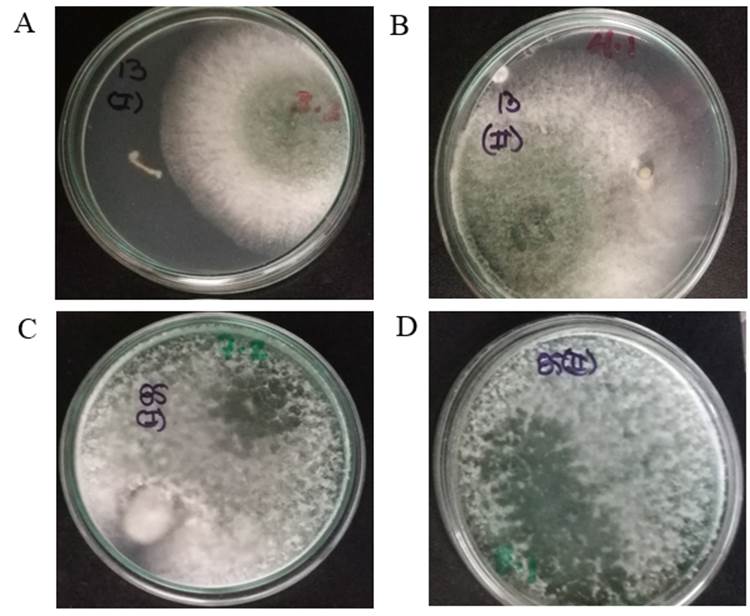
Figura 1 Colonización de las raíces de P. vulgaris (cv. BAT-304) por T. asperellum en plantas sembradas al unísono con la inoculación (A y C: Ta. 13 y Ta. 85, respectivamente) y siete días después de la inoculación (C y D: Ta. 13 y Ta. 85, respectivamente) / Colonization of roots of P. vulgaris (cv. BAT-304) by T. asperellum in plants planted at soil inoculation (A and C: Ta. 13 and Ta. 85, respectively) and seven days after inoculation (C and D: Ta. 13 and Ta. 85, respectively)
La penetración y colonización de la planta por Trichoderma spp., asociada con su capacidad de antibiosis, potencia la síntesis de un arsenal enzimático hidrolítico, degradador de la pared celular de la planta (20). Se piensa que esta entrada en las capas epidérmicas de las raíces confiere un mayor crecimiento, mejora la absorción de nutrientes lo que se revierte en un aumento de la productividad de la planta (21).
T. asperellum favoreció la acumulación de nitrógeno (N) en las hojas de las plantas. Las concentraciones de nitrógeno foliar fueron significativamente mayores con respecto al testigo en plantas inoculadas con las cepas Ta. 85 y Ta. 90, en cualquiera de los momentos de siembra. Por otra parte, la concentración de N foliar producida por la cepa Ta. 78 solo fue significativamente mayor a la del testigo durante el segundo momento de siembra. Las plantas que fueron sembradas siete días después en suelo inoculado con Ta. 78 mostraron la mayor concentración de potasio (K) foliar (Tabla 2); sin embargo, ninguno de los tratamientos aumentó el contenido de fósforo (P) con respecto a las plantas control.
Tabla 2 Contenidos de nutrientes en la biomasa aérea de P. vulgaris (cv. BAT-304) inoculado con T. asperellum. Medias con una letra común no son significativamente diferentes (p> 0.10) / Nutrient content in the aerial biomass in P. vulgaris (cv. BAT-304) sown in soil inoculated with T. asperellum. Media with a common letter are not significantly different (p> 0.10)
| Momento de siembra | Tratamiento | % en base seca | ||
|---|---|---|---|---|
| N | P | K | ||
| Testigo | 3,2 ± 0,6 cd | 0,44 ± 0,06 a | 2,9 ± 0,36 ab | |
|
|
4,41 ± 0,6 bc | 0,27 ± 0,06 b | 3,15 ± 0,36 ab | |
|
|
2,79 ± 0,59 d | 0,36 ± 0,06 ab | 3,03 ± 0,36 ab | |
|
|
0,49 ± 0,05 a | 3,09 ± 0,29 ab | ||
|
|
0,46 ± 0,06 a | 3,17 ± 0,36 ab | ||
| Testigo | 3,2 ± 0,6 cd | 0,44 ± 0,06 a | 2,9 ± 0,36 ab | |
|
|
4,22 ± 0,49 bc | 0,37 ± 0,05 ab | 2,79 ± 0,29 b | |
|
|
0,38 ± 0,06 ab | |||
|
|
0,28 ± 0,05 b | 3,36 ± 0,29 ab | ||
|
|
0,42 ± 0,05 a | 3,39 ± 0,29 ab | ||
Algunos aislados de Trichoderma pueden solubilizar nutrientes cercanos a las raíces, lo que permite que estas sustancias puedan ser asimiladas por la planta. López-Bucio et al. (22) plantearon que la aplicación de Trichoderma, en la etapa temprana del crecimiento del cultivo, permite maximizar sus beneficios en términos de desarrollo de la raíz y la absorción de nutrientes. Se demostró que la proteína QID74 de la pared celular de T. harzianum, rica en cisteína, está involucrada en la modificación de la arquitectura de la raíz, aumentando la superficie de absorción total, facilitando la absorción y la translocación de nutrientes en los brotes, lo que resulta en un aumento de la biomasa de las plantas a través de un uso eficiente de N, P, K y micronutrientes (17).
La adquisición de nitrógeno es un factor importante para el desarrollo y la productividad de los cultivos, debido a que este macronutriente está presente en forma compleja o en una cantidad muy limitada, lo que dificulta su disponibilidad para las plantas. A diferencia de otras leguminosas, P. vulgaris es un pobre fijador de nitrógeno lo cual se atribuye a algunos factores genéticos y su sensibilidad al estrés abiótico y biótico, lo que puede limitar la productividad del cultivo. Como leguminosa, el frijol común forma simbiosis con una amplia gama de bacterias del género Rhizobium que fijan el nitrógeno biológicamente en los nódulos (23).
Los resultados del presente trabajo no coincidieron con los de Hoyos-Carvajal et al. (24), quienes no hallaron incrementos significativos en el contenido foliar de N en plantas frijol inoculadas con aislamientos de Trichoderma spp. Singh et al. (25) demostraron que T. asperellum T42 incrementa el contenido total de nitrógeno en plantas de tabaco (Nicotiana tabacum L.) debido, tanto a cambios en la estructura de la raíz, como a la activación de la enzima nitrato reductasa facilitando la asimilación de N en forma de grupos nitrato, nitro y amonio. Se ha planteado la hipótesis de que el uso de Trichoderma podría reducir las cantidades de fertilizantes nitrogenados utilizados en un 40 a 50 % sin una reducción en el rendimiento de los cultivos (26).
El fósforo, con frecuencia, es el macronutriente menos accesible en muchos ecosistemas y su baja disponibilidad, a menudo, limita el crecimiento de las plantas (27). Contrario a lo esperado, las cepas de T. asperellum usadas durante este estudio no produjeron cambios en el contenido de fósforo foliar con respecto al control. Hoyos-Carvajal et al. (10) observaron que solo algunos aislamientos de T. asperellum son capaces de solubilizar el fosfato a partir de Ca3PO4 y/o roca fosfórica, aumentando su disponibilidad para la planta. Además, en otro estudio encontraron que T. asperellum T30 incrementó el porcentaje de P en el tejido de la planta con respecto al control, pero sin presentar diferencias estadísticas significativas; mientras que, T. asperellum T46 disminuyó el nivel de este nutriente con respecto al control (24). En pepino (Cucumis sativus L.), se encontró que, a partir de la colonización de las raíces por T. asperellum, el hongo aumenta la disponibilidad de P en la planta, lo que lleva a un aumento significativo en su masa seca, longitud de brotes y área foliar (28). Estos resultados sugieren que las cepas de T. asperellum estudiadas podrían no ser capaces de solubilizar el fosfato del suelo lo que debe ser corroborado mediante estudios de solubilización in vitro.
De igual manera, solo la cepa Ta. 78 incrementó el contenido de K foliar, aunque sin mostrar diferencias significativas con respecto al control, resultados semejantes a los obtenidos por Hoyos-Carvajal et al. (24), quienes hallaron en condiciones experimentales con suelos andisoles, que Trichoderma spp. incrementó la concentración foliar en P. vulgaris. Por otro lado, Marra et al. (29) encontraron que las semillas de soya (Glycine max L. ) tratadas con cepas de Trichoderma spp. mostraron una disminución en la concentración de K en comparación con los controles. De manera general, el contenido de nutrientes se favoreció cuando las plantas se sembraron siete después de la inoculación del suelo con el hongo. Estos resultados sugieren que las hifas de Trichoderma colonizan el frijol durante los estadios tempranos de desarrollo vegetal pudiendo modificar la estructura de la raíz lo cual incrementa la disponibilidad de los nutrientes para la planta.
Cada una de las enzimas de defensa fueron inducidas en las hojas por una o más de las cepas de T. asperellum en estudio (Figura 2). Ta. 78 estimuló la inducción de POD, PPO y PAL con diferencias significativas con respecto al control. Para el caso de las producciones de POD y PPO, estas enzimas se expresaron en mayor medida en las plantas sembradas siete días después de la inoculación del hongo, momento para el cual Ta. 78 ya se había establecido en el suelo. También esta cepa indujo los mayores niveles de actividad específica PAL cuando las plantas de frijol se sembraron en el primer momento de siembra.
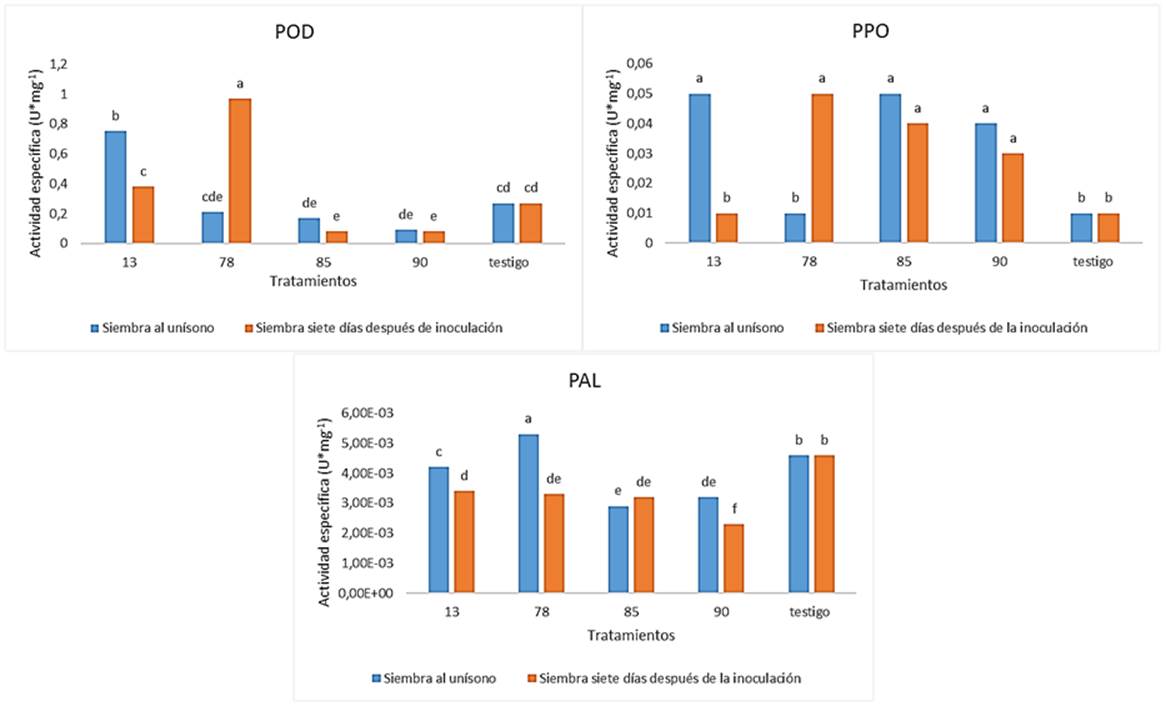
Figura 2 Efecto de cuatro cepas de T. asperellum sobre la producción de enzimas defensivas en el frijol (cv. BAT-304), en dos momentos de siembra de las semillas con relación a la aplicación del hongo al suelo. Medias con una letra común no son significativamente diferentes (p > 0.10) / Effect of T. asperellum on production of defence enzymes in beans (cv. BAT-304). Media with letters in common are not significantly different (p > 0.10)
La cepa Ta. 13 produjo incremento en los niveles de actividad enzimática POD y PPO con diferencias significativas con respecto al control cuando la siembra y la inoculación de Trichoderma se realizaron simultáneamente. Por su parte, Ta. 85 y Ta. 90 aumentaron la actividad enzimática de PPO, con diferencias significativas, con respecto al control, tanto en el primer momento de siembra como en el segundo. Estos resultados semejan a los obtenidos por Chowdappa et al. (13), que mostraron que la cepa OTPB3 de T. harzianum producía inducción de resistencia en plantas de tomate inoculadas mediante el incremento de los niveles de enzimas defensivas entre las que se incluían las POD y las PPO. Zhang et al. (30) observaron que la inoculación de la raíz con la cepa T soybean de T. harzianum aumentó significativamente las actividades de POD, PPO y PAL y otras enzimas antioxidantes como catalasa, superóxido dismutasa, ascorbato peroxidasa y glutatión reductasa de las hojas de pepino, lo que indica que Trichoderma podría conferir resistencia sistémica a todas las plántulas de pepino tratadas, al mejorar las actividades de las enzimas antioxidantes.
La reducción de las actividades enzimáticas en algunos de los tratamientos con T. asperellum podría tener relación con lo planteado por Dildey et al. (20) quienes argumentaron que Trichoderma spp., estimula a la planta al equilibrio de su arsenal enzimático, en su propio beneficio, evitando que la planta ponga en peligro la relación simbiótica durante la interacción con Trichoderma spp. y obstaculice su desarrollo.
Los patrones isoenzimáticos de las POD y la PPO revelaron un total de seis y cuatro bandas, respectivamente (Figura 3). En el caso de las POD, las plantas control mostraron el mayor número de bandas (seis bandas). Los tratamientos con Ta. 13 (primer momento de siembra) y Ta. 78 (segundo momento de siembra) mostraron cada uno cinco isoenzimas y estas presentaron la mayor intensidad lo que se corresponde con los valores de actividad enzimática obtenidos durante el ensayo. En ambos tratamientos mencionados anteriormente, la isoenzima correspondiente con la banda 6 se mostró reprimida. El tratamiento con Ta. 90 en el segundo momento de siembra presentó la menor cantidad de isoenzimas, observándose solo tres bandas. De la misma manera, los tratamientos con Ta. 13 (primer momento de siembra) y Ta. 78 (segundo momento de siembra) presentaron semejante patrón isoenzimático, ambos con cuatro bandas. En estos casos, se evidenció que la isoenzima correspondiente a la banda 4 presentó mayor concentración. El cambio sobre el patrón isoenzimático demostrado por hojas de frijol de las plantas tratadas con las diferentes cepas de T. asperellum confirma el efecto inductor de defensa sistémica de estas cepas. Las POD y las PPO son además enzimas antioxidantes que participan en los procesos de eliminación de los excesos de especies reactivas del oxígeno (30).
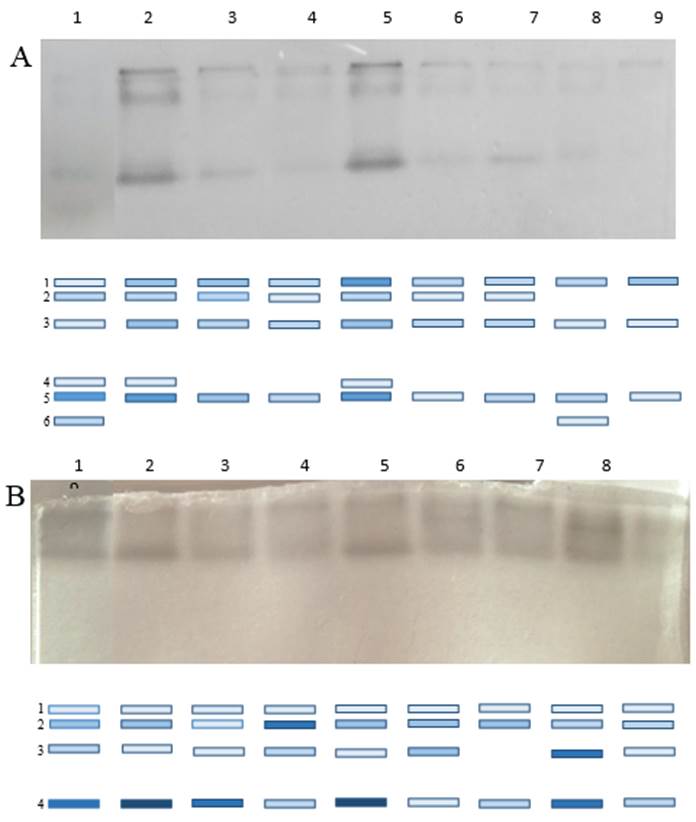
Figura 3 Patrón isoenzimático y cromatograma de enzimas POD (A) y PPO (B) de hojas de frijol tratadas cepas de T. asperellum. 1: control; 2: Ta. 13, momento I; 3: Ta. 13, momento II; 4: Ta. 78, momento I; 5: Ta. 78, momento II; 6: Ta. 85, momento I; 7: Ta. 85, momento II; 8: Ta. 90, momento I; 9: Ta. 90, momento II / Isoenzymatic pattern and chromatogram of POD (A) and PPO (B) enzymes from leaves of bean plants treated with strains of T. asperellum. 1: control; 2: Ta. 13, moment I; 3: Ta. 13, moment II; 4: Ta. 78, moment I; 5: Ta. 78, moment II; 6: Ta. 85, moment I; 7: Ta. 85, moment II; 8: Ta. 90, moment I; 9: Ta. 90, moment II
CONCLUSIONES
Se encontró que la cepa Ta. 85 puede ser usada, fundamentalmente, como promotora del sistema radical de las plantas de frijol. T. asperellum exhibió capacidad endofítica en frijol y su utilidad en el incremento del contenido foliar de N (Ta. 78, Ta. 85 y Ta. 90) y K (Ta. 78). Por su parte, la cepa Ta. 78 podría ser más eficiente en la inducción de resistencia sistémica. Los resultados revelan que las cepas estudiadas pueden ser candidatas para el desarrollo de biofertilizantes.

