










 Servicios personalizados
Servicios personalizados texto en
texto en  Inglés (pdf)
Inglés (pdf)
 Articulo en XML
Articulo en XML Referencias del artículo
Referencias del artículo
 Enviar articulo por email
Enviar articulo por email Citado por SciELO
Citado por SciELO  Similares en
SciELO
Similares en
SciELO 
 Permalink
PermalinkINTRODUCCIÓN
En los bosques naturales y otras áreas forestales dentro de estas, se encuentra la flora utilizada por las abejas, en donde, el polen apícola puede ser considerado como subproducto de los bosques y promoverse como producto forestal no maderable (PFNM), de tal manera que, permite considerar a la apicultura como una actividad que puede ser integrada al manejo y conservación de las especies forestales y de los ecosistemas donde habitan (Chamorro et al., 2013).
La apicultura es una actividad que aprovecha la vegetación, tanto en su estado natural como alterado, así como los cultivos agrícolas y forestales, en este mismo sentido hay especies vegetales que florecen muy temprano, e incentivan la colmena para que comience a reproducirse y otras especies que lo hacen sucesivamente a lo largo del año (Paredes, 2017).
Los estudios realizados por la Organización de las Naciones Unidas para la Alimentación y la Agricultura (FAO), (2018) revela que la polinización es el proceso más importante de la naturaleza que contribuye a la biodiversidad. Debido a que un 90 % de las plantas con flores dependen de la polinización para subsistir, por esta razón, las abejas mantienen los ecosistemas forestales conservando la biodiversidad de estos.
Por otra parte, Méndez et al. (2021), indicaron que la relación entre las abejas y los recursos florales, se realizan a través de estudios palinológicos, en el que permiten identificar las diferentes especies botánicas que contribuyen al alimento de las abejas.
En la provincia de Manabí, en la actualidad diversos cantones se dedican a la actividad apícola, haciendo énfasis en la producción de miel, propóleo, cera, entre otros. El recinto Quimis del cantón Jipijapa, es considerado como una de las zonas pobladas por muchos árboles, pero en especial de Ceiba trischistandra (A. Gray) Bakh. (ceibo) y Prosopis juliflora var. juliflora (algarrobo), que son fuentes directas de néctar y polen, que es un lugar habitado por apicultores que aprendieron a generar ingresos económicos mediante dicha actividad y todo lo que ella engloba. Por todo lo antes expuesto, el objetivo del este estudio fue evaluar la morfología del polen de especies melíferas del bosque seco tropical del recinto Quimis, orientado a la conservación.
MATERIALES Y MÉTODOS
Caracterización del área de estudio
Esta investigación se desarrolló en los apiarios activos en el área de influencia del recinto Quimis, que se localiza en la parroquia Membrillal, del cantón Jipijapa. Este recinto está situado en el kilómetro 21 de la vía que conecta los cantones Portoviejo y Manta, de la provincia de Manabí, en la región de la costa del pacífico ecuatoriano.
Metodología
Identificación el origen botánico y morfológico del polen apícola proveniente de apiarios del recinto Quimis
Para la realización de la captura del polen, se indagó a los apicultores que forman parte de Asociación ASOPROAPIMIEL del recinto, con la finalidad de saber los apiarios activos de la zona, de tal forma que, se procedió a seleccionar 10 colmenas de Apis mellifera, en cinco apiarios activos dentro del recinto, cuya distancia de la vivienda hacia el apiario varía entre los 300 a 500 m, siguiendo la metodología de De Boada et al. (1987) y De Boada y Cogua (1989).
Para recolectar el polen, se ubicaron trampas de caza polen en las piqueras de las colmenas (entrada de la colmena), por un período de 12 a 24 horas específicamente en tres días hábiles con permiso del apicultor durante dos semanas, en los meses de mayo, junio y julio de 2023, según la metodología propuesta por Hidalgo et al. (1990) y Sayas y Huamán (2009).
Para identificar botánicamente el polen, se trasladaron las muestras al laboratorio de biotecnología de la Universidad Estatal del Sur de Manabí (UNESUM) y observadas con un microscopio óptico modelo Better Scientific con un lente objetivo de 4/0.10.
Las muestras de polen se colocaron en placas petri, tras haber sido tamizadas para eliminar impurezas. Luego, se subdividieron por apiario y, dentro de cada uno, se hicieron subdivisiones adicionales según el color y peso de las muestras.
Para la preparación de las muestras, se realizaron dos soluciones. La primera combinó glicerina y agua destilada (5 ml de cada una) en placas Petri (Insuasty et al., 2017), mezclándolas hasta obtener una consistencia homogénea. A esta solución se añadieron los granos de polen limpios para su hidratación y distribución uniforme, dejándolos reposar 48 horas.
Después de este periodo, se colocó una gota de la solución en un portaobjetos, cubriéndola con un cubreobjetos. Para la segunda preparación, se usaron dos gotas de solución de fucsina diluida en 1 ml de etanol, con el fin de resaltar las características del polen. Este proceso se repitió tres veces para garantizar la fiabilidad de los análisis.
Para una mejor visualización de la morfología del polen (Figura 1 a la 4), las muestras se examinaron bajo un microscopio con cámara modelo Euromex, con un lente objetivo de 40/0,65; en el laboratorio de parasitología de la carrera de Laboratorio Clínico de la UNESUM.
Finalmente, para determinar el origen botánico del polen, se consultaron Palinotecas virtuales y recursos como la guía rápida de Polen de las Islas Galápagos (Jaramillo y Del Mar, 2011), el Catálogo Fotográfico de especies de Flora apícola (Velandia et al., 2012) y Montoya et al. (2014).
Una vez identificadas las especies, se procedió a registrar la morfología del polen, para lo cual se tuvo en cuenta las características descritas por Jaramillo y Del Mar, (2011), a saber:
Simetría (radiosimétrico, bisimétrico, asimétrico)
Tamaño (< 10 µm granos de polen muy pequeños y > 200 µm granos de polen gigantes)
Unidades de grano de polen (mónada, diadas, tétradas, poliadas)
Forma de exina (psilada, fosulada, foveolada, escábrida, equinada, baculada, gemada, pilada, reticulada, verrugosa)
Sistema NPC mediante el número, posición, caracteres del polen.
Forma que presenta el polen.
Según los autores mencionados, el análisis microscópico de las muestras de polen recolectadas en un período específico revela las variedades de especies que las abejas han utilizado como fuentes de polen.
Evaluación de Parámetros
Para evaluar los parámetros poblacionales, se emplearon herramientas de ofimática como Microsoft Excel, así como programas especializados como Infostat y GeoGebra. Utilizando los datos recopilados, se calcularon la frecuencia relativa y absoluta en relación con la cantidad de especies identificadas por apiario. Este análisis incluyó tanto las características morfológicas como el peso en gramos (g) del polen recolectado.
La taxonomía y nomenclatura, así como la categoría de amenaza de las especies citadas en el área de estudio se revisaron en las obras de Ministerio del Ambiente (MAE, 2013); Aguirre et al. (2014); Chimarro et al. (2023); Jaramillo et al. (2024). Los nombres comunes fueron proporcionados por los guías locales como describe Jiménez et al. (2021)
RESULTADOS Y DISCUSIÓN
Identificación del origen botánico y morfológico del polen proveniente de apiarios del recinto Quimis
Los resultados del análisis microscópico de la carga de polen se reflejan en la Tabla 1, donde se aprecian 21 tipos polínicos correspondientes a especies que pertenecen a 13 familias de plantas, distribuidas alrededor de los cinco apiarios objeto de estudio en el recinto Quimis.
Tabla 1. - Especies inventariadas en el Bosque Seco Tropical del recinto Quimis
| N.º | Especie | Familia |
| 1 |
|
Convolvulaceae |
| 2 |
|
Fabaceae |
| 3 |
|
Malvaceae |
| 4 |
|
Convolvulaceae |
| 5 |
|
Muntingiaceae |
| 6 |
|
Malvaceae |
| 7 |
|
Fabaceae |
| 8 |
|
Primulaceae |
| 9 |
|
Capparaceae |
| 10 |
|
Polygonaceae |
| 11 |
|
Fabaceae |
| 12 |
|
Poaceae |
| 13 |
|
Myrtaceae |
| 14 |
|
Capparaceae |
| 15 |
|
Fabaceae |
| 16 |
|
|
| 17 |
|
Boraginaceae |
| 18 |
|
Fabaceae |
| 19 |
|
Bignoniaceae |
| 20 |
|
|
| 21 |
|
Malvaceae |
Las especies inventariadas por familias se presentan seguidamente según las características morfológicas del polen, incluyendo unidades de polen, simetría, aberturas, forma de exina, forma polar/ecuatorial y tamaño.
Familia Convolvulaceae
Convolvulus arvensis L. (Figura 1, No. 1a - 1b).
Descripción botánica: Hierbas perennes con yemas en la superficie del suelo, adaptadas en el bosque seco pluvioestacional y bosque seco andino; hojas simples con borde lobulado, forma triangular, de textura carnosa; tipo de tallo herbáceo de corteza lisa (Jiménez et al., 2021). Unidad de polen tipo mónada; granos con simetría radiosimétrico; aberturas tipo pantocolporado; forma de exina psilada; forma P/E tipo esferoidal; tamaño grande.
Xenostegia medium (L.) D. F. Austin & G. W. Staples (Figura 1, No. 2a -2b).
Descripción botánica: Hierbas con tallos de 50 cm de altura, adaptados en bosque seco pluvioestacional y bosque seco andino; hojas simples, borde lobulado, forma del limbo cordada, de textura membranosas; tipo de tallo herbáceo de corteza lisa (Jiménez et al., 2021). Unidad de polen tipo mónada; granos con simetría radiosimétrico; aberturas tipo polipantoporado; forma de exina equinada; forma P/E tipo esferoidal; tamaño grande.
Familia Bignoniaceae
Handroanthus billbergii (Bureau & K. Schum.) S.O. Grose (Figura 1, No. 3a - 3b)
Descripción botánica: Árbol de gran tamaño, hojas compuestas, flores con cáliz campanulado, corola tubular (Aguirre, 2012). Unidad de polen tipo mónada; granos de polen radiosimétrico; abertura triporado; forma de exina psilada; forma P/E tipo esferoidal; tamaño mediano.
Familia Cannabaceae
Trema micrantha (L.) Bl. (Figura 1, No. 4a - 4b)
Descripción botánica: Árbol perennifolio entre 5 m -13 m de altura, adaptado en bosque seco pluvioestacional, bosque seco andino, bosque siempre verde de tierras bajas, chocó; hojas simples, borde entero, forma del limbo elíptico; de textura membranosa (Jiménez et al., 2021). Unidad de polen tipo mónada; grano de polen bisimétrico; abertura diporados; forma de exina psilada; forma P/E tipo esferoidal; tamaño pequeño.
Familia Capparaceae
Colicodendron scabridum (Kunth) Hutchinson (Figura 1, No. 5a - 5b).
Descripción botánica: Árboles de 15 m 30 m de altura, adaptados en bosque seco pluvioestacional y bosque seco andino; hojas simples, borde entero, forma del limbo, zarcillo, de textura coriácea, duras, semejantes al cuero (Jiménez et al., 2021). Unidad de polen tipo mónada; granos con simetría bisimétrico; aberturas tipo tricolpado; forma de exina psilada; forma P/E tipo prolato-esferoidal; tamaño pequeño.
Capparicordis crotonoides (Kunth) Iltis & Cornejo (Figura 1, No. 6a - 6b).
Descripción botánica: Plantas leñosas entre 2 m 5 m, adaptado en bosque seco pluvioestacional; hojas simples, borde entero, forma del limbo lanceolado, de textura coriáceo, duras, semejantes al cuero (Jiménez et al., 2021). Unidad de polen tipo mónada; granos con simetría bisimétrico; aberturas tipo tricolpado; forma de exina fosulada; forma P/E sub-prolato; tamaño mediano.
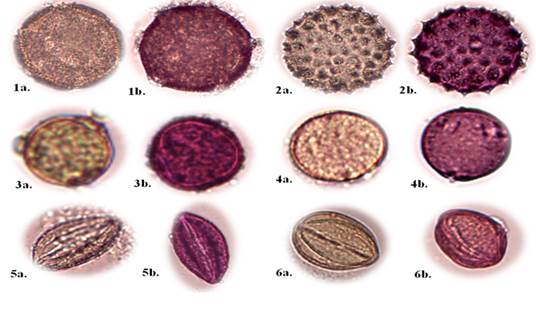 Nota: (A) grano de polen sin fuscina; (B) grano de polen con fuscina Familia Boraginaceae
Nota: (A) grano de polen sin fuscina; (B) grano de polen con fuscina Familia BoraginaceaeFig. 1. - Descripción de las características morfológicas del polen (400x)
Cordia macrantha Chod. (Figura 2, No. 7a - 7b).
Descripción botánica: Árbol de 15 m de altura, adaptadas en bosque seco pluvioestacional y bosque seco andino, hojas simples, flores simples blancas enteras, corteza gris (Aguirre, 2012). Unidad de tipo de polen: Mónada; granos con simetría radiosimétrico; aberturas tipo tricolporado; forma de exina reticulada; forma P/E prolato; tamaño pequeño.
Familia Fabaceae
Prosopis pallida (Willd.) Kunth (Figura 2, No. 8a - 8b).
Descripción Botánica: Árboles pequeños o medianos, de 8 m - 15 m, adaptadas en bosque seco pluvioestacional y bosque seco andino; hojas compuestas bifoliadas; borde entero; forma del limbo linear; de textura membranosa, sumamente blanda (Jiménez et al., 2021). Unidad de tipo de polen: Mónada; granos con simetría radiosimétrico; aberturas tipo tricolporado; forma de exina psilada; forma P/E prolato; tamaño mediano.
Caesalpinia paipai Ruiz & Pav. (Figura 2, No. 9a -9b).
Descripción botánica: Árboles pequeños o medianos, de 8 m 15 m, adaptados en bosque seco tropical; hojas compuestas bifoliadas; borde de la hoja entero; forma de la hoja oval; textura de hoja membranosa, sumamente blanda (Jiménez et al., 2021). Unidad de tipo de polen: Mónada; granos con simetría radiosimétrico; aberturas tipo tricolporado; forma de exina reticulada; forma del polen oblato esferoidal; tamaño mediano.
Cassia fistula L. (Figura 2, No. 10a -10b).
Descripción botánica: Es un árbol caducifolio de tamaño mediano, alcanzando 10 m de altura, con un tronco recto de hasta 5 m y 1 m de diámetro. Posee corteza gris pálida en ejemplares jóvenes y marrón oscuro en los viejos. Sus hojas son alternas, pinnadas, con 4-8 pares de foliolos ovados. Las flores son amarillas, en racimos colgantes, con cáliz oblongo y corola de cinco pétalos subiguales (Bhandari et al., 2013). Unidad de tipo de polen: Mónada; granos con simetría radiosimétrico; aberturas tipo tricolporado; forma de exina psilada; forma del polen esferoidal; tamaño pequeño.
Mimosa sensitiva L. (Figura 2, No. 11a -11b).
Descripción botánica: Arbustos de hasta 3 m de altura con tallos teretes o subteretes, acúleos, unguiculados y estrigosos. Hojas con estípulas pubescentes y setosas, pecíolos de 2-9 cm y folíolos oblongo-elípticos a oblanceolados. Inflorescencias con pedúnculos de 1-3.5 cm y capítulos esferoides híspidos. Flores fértiles, 4-meras y 4-andras, con cáliz papiforme y corola glabra o pubérula. Craspedios generalmente 2-5-articulados y híspidos (Secretaría de Global Biodiversity Information Facility [GBIF] 2023). Unidad de tipo de polen: Romboidal; granos con simetría asimétrica; aberturas tipo inaperturado; forma de exina psilada; forma del polen oblato; tamaño muy pequeño.
Leucaena leucocephala(Lam.) de Wit (Figura 2, No. 12a -12b).
Descripción botánica: Es un arbusto o árbol pequeño, perenne y sin espinas, de hasta 8 m de altura con raíces profundas. Sus hojas son alternas, bipinnadas, con 12 a 18 pares de folíolos pequeños y glabros. Las flores son blancas a blanco amarillentas, formadas en cabezas globosas axilares. El fruto es una vaina lineal, delgada, de 12 a 14 cm de largo, que contiene de 15 a 30 semillas ovado-oblongas o elípticas. Las semillas son verdes cuando inmaduras y marrón oscuro al madurar (Nehdi et al., 2014). Unidad de tipo de polen: Mónada; granos con simetría radiosimétrico; aberturas tipo tricolporado; forma de exina reticulada; forma del polen prolato-esferoidal; tamaño mediano.
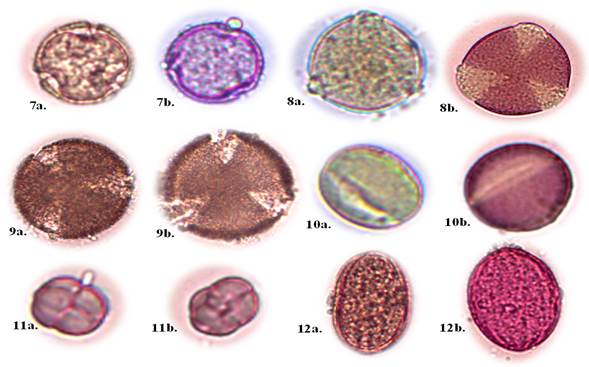 Nota: (A) grano de polen sin fuscina; (B) grano de polen con fuscina Familia Malvaceae
Nota: (A) grano de polen sin fuscina; (B) grano de polen con fuscina Familia MalvaceaeFig. 2. - Descripción de las características morfológicas del polen (x400).
Eriotheca ruizii (K. Schum.) A. Robyns (Figura 3, No. 13a -13b).
Descripción botánica: Árboles de 15-30 m, adaptadas en bosque seco pluvioestacional; hoja simple; borde lobulado; forma elíptica; con textura membranosa sumamente blanda (Jiménez et al., 2021). Unidad de tipo de polen: Mónada; granos con simetría radiosimétrico; aberturas tipo tricolporado; forma de exina reticulada; forma del polen oblato; tamaño pequeño.
Ceiba trischistandra (A. Gray) Bakh. (Figura 3, No. 14a -14b).
Descripción botánica: Árboles de gran tamaño sobre los 30 m, adaptados en bosque seco pluvioestacional y bosque seco andino; tipo de hoja compuesta, palmadas; borde entero; forma del limbo eliptica; de textura cartaceas, estilos papel o cartulina (Jiménez et al., 2021). Unidad de tipo de polen: Mónada; granos con simetría radiosimétrico; aberturas tipo tetracolporado; forma de exina reticulada; forma del polen esferoidal; tamaño grande.
Hibiscus rosa-sinensisL. (Figura 3, No. 15a -15b).
Descripción botánica: Árbol o arbusto de 23 m de altura con tallos ramificados. Hojas alternas, simples, ovadas u ovadolanceoladas, dentadas, de color verde intenso y lustrosas, de 612 cm. Flores de 812 cm de diámetro, actinomorfas, hermafroditas, solitarias, con 5 pétalos más largos que el cáliz, en colores como rojo intenso, blanco, amarillo o anaranjado. Fruto en cápsula loculicida, polispermo, con semillas reniformes, glabras o pubescentes (Greuter y Rankin, 2017). Unidad de tipo de polen: Mónada; granos con simetría radiosimétrico; aberturas tipo pantoporado; forma de exina equinada; forma del polen esferoidal; tamaño pequeño.
Familia Muntingiaceae
Muntingia calabura L. (Figura 3, No. 16a -16b).
Descripción botánica: Árboles de 15 m - 30 m de altura, adaptados en bosque seco pluvioestacional y bosque seco andino; tipo de hojas simples; con bordes dentados; forma del limbo lobulado; de textura carnosa (Jiménez et al., 2021). Unidad de tipo de polen: Mónada; granos con simetría radiosimétrico; aberturas tipo tricolporado; forma de exina reticulada; forma del polen esferoidal; tamaño pequeño.
Familia Myrtaceae
Eucalyptus globulus Labill. (Figura 3, No. 17a -17b).
Descripción botánica: Árbol que normalmente alcanza 45 m, pero puede superar los 70 m. Corteza lisa, de color blanco crema, amarilla o gris, que se desprende en tiras. Hojas juveniles sésiles, opuestas y glaucas; hojas adultas lanceoladas a estrechamente lanceoladas. Umbelas con 1, 3 o 7 flores. Yemas turbinadas u obcónicas; frutos obcónicos, hemisféricos o subglobulares con disco a nivel o ascendente y valvas 3 o 5 (Darriba y Pando, 2016). Unidad de tipo de polen: Mónada; granos con simetría radiosimétrico; aberturas tipo tricolporado; forma de exina psliada; forma del polen oblato; tamaño grande.
Familia Poaceae
Zea mays L. (Figura 3, No. 18a -18b).
Descripción botánica: planta anual, con fotosíntesis C4, cultivada por el valor alimenticio de sus frutos, ya sea para consumo humano como de animales; espiga cilíndrica. Las espiguillas se hallan dispuestas alrededor del eje principal, como ocurre en la inflorescencia femenina; las anteras son mesifijas. Las tecas normalmente tienen dehiscencia longitudinal, pero hay anteras con dehiscencia por poro apical (Gutiérrez, 2023).
Unidad de tipo de polen: Mónada; granos con simetría bisimétrico; aberturas tipo monoporados; forma de exina escábrida; forma del polen oblato esferoidal; tamaño grande.
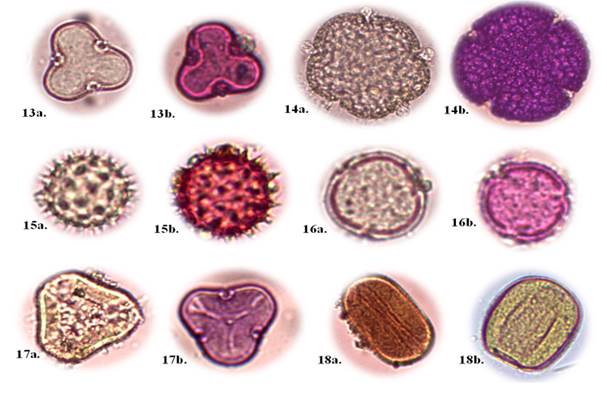
Familia Polygonaceae
Triplaris cumingiana Fisch. & Mey. ex C. A. Mey. (Figura 4, No. 19a -19b).
Descripción botánica: Árbol de 10 m -18 m de altura; flores agrupadas en racimos de 5 cm -35 cm de longitud, rojo cuando son jóvenes y luego amarillentas; flores femeninas con perianto 3lobulados; flores masculinas en grupos de 3-5, con perianto 6-lobulado, 9 estambres (Zamora y Jiménez, 2000). Unidad de tipo de polen: Mónada; granos con simetría bisimétrico; aberturas tipo tricolporado; forma de exina psilada; forma del polen prolato; tamaño mediano.
Familia Primulaceae
Bonellia sprucei (Mez) B. Ståhl & Källersjö (Figura 4, No. 20a -20b).
Descripción Bótanica: Árboles pequeños o medianos, de 8 m - 15 m, adaptados al bosque seco pluvioestacional; tipo de hojas compuestas, imparipinnadas; bordes enteros; forma del limbo oblanceolada; de textura coriaceas, dura semenjante al cuero (Jiménez et al., 2021). Unidad de tipo de polen: Mónada; granos con simetría radiosimétrico; aberturas tipo tricolporado; forma de exina reticulada; forma del polen sub-prolato; tamaño mediano.
Familia Verbenaceae
Lantana camara L. (Figura 4, No. 21a -21b).
Descripción botánica: Arbusto leñoso perenne de 2 m a 5 m de altura; hojas opuestas, lanceoladas; bordes crenados a dentados, haz áspero, flores axilares y terminales densas (Matienzo et al., 2003). Unidad de tipo de polen: Mónada; granos con simetría bisimétrico; aberturas tipo estefanocolpado; forma de exina reticulada; forma del polen esferoidal; tamaño mediano.

Frecuencia de las familias y especies poliníferas más visitadas por la Apis mellifera de acuerdo con los datos obtenidos en cada apiario
Los resultados de la frecuencia relativa indicaron que las familias botánicas que presentan mayor interés como recurso polínico para las abejas son, Fabaceae con un 23,1 %, seguido de la Malvaceae con el 14,29 %, a diferencia de las restantes familias que presentaron valores semejantes entre ellas.
La familia con mayor número de especies melíferas fue la Fabaceae, en este sentido se destacan las especies, Prosopis pallida, Caesalpinia paipai, Cassia fistula, Mimosa sensitiva, Leucaena leucocephala.
En el caso de las especies que resultaron con una frecuencia menor que el 6 % en relación con la recolección de polen, se ha comprobado que, la especie Apis mellifera utiliza estas especies de plantas como un recurso alternativo cuando existe una baja disponibilidad de oferta floral que aporten gran cantidad de polen en el área (Girón, 1995). Este autor reporta que las abejas colectan polen de muy pocas especies de plantas en una proporción mayor del 10 %, las cuales pueden considerarse como un "recurso alimenticio importante", mientras que pueden colectar polen de un gran número de especies vegetales, pero en pequeñas proporciones y a estas plantas se les denomina "recurso alimenticio alternativo"; por su parte, Hidalgo et al. (1990) consideran un aspecto diferente relatando que, Apis mellifera utiliza como fuente de polen solo unas pocas especies y que otras son utilizadas ocasionalmente, basándose en la selección de aquellas especies que le sean más ventajosas.
En lo referente a la investigación de la morfología de polen, se evidenció la presencia de 21 especies melíferas correspondientes a 13 familias, siendo las especies más frecuentes, Convolvulus arvensis, Ceiba trischistandra, Eriotheca ruizii, Prosopis pallida, en este mismo sentido, los resultados son similares a los reportados por Hidalgo et al. (1990), quienes registraron 21 tipos polínicos que corresponden a 13 familias; de igual manera esta información se acerca a los resultados obtenidos por Ramírez et al. (2016), quienes reportaron 17 especies distribuídas en 13 familias, siendo los más importantes Dyssodia papposa, Tithonia tubaeformis y Leucaena leucocephala, durante los meses de noviembre a marzo en las regiones centro y norte del estado de Guerrero, México, no obstante estos resultados difieren a los documentados por Girón (1995), quién reportó 91 tipos polínicos de los cuales el más frecuente fue de la especie Coffea arabica dentro del municipio de Salgar en Antioquia, Colombia.
En el orden de las ideas anteriores destacan las características morfológicas específicas de los granos de polen que, en la familia Fabaceae predomina el polen de tipo mónada. En cuanto a la forma, prevalecieron las morfologías prolato y oblato-esferoidal, seguidas por oblato y prolato-esferoidal. En términos de aberturas, la tricolporada fue la más común observada. Respecto a la exina, se identificaron morfologías psiladas y reticuladas, predominantemente. Estos resultados coinciden con los reportes de Ventura y Huamán (2008), quienes informaron características morfológicas similares en las especies de la familia Fabaceae, todo lo cual corrobora lo descrito por Ferreira et al. (2013), quienes reportaron el tipo mónada como el más frecuente entre los granos de polen estudiados.
Dentro de las categorías que abordan las características morfológicas del polen, la simetría, que determina la vista polar del grano, mostró una mayor frecuencia para la simetría radiosimétrica, seguida por la simetría bisimétrica, que fue observada en aproximadamente seis especies. Solamente una especie presentó asimetría, lo que significa que no exhibe ningún plano de simetría. Estos resultados concuerdan con la investigación de Ferreira et al. (2013), quienes, en su estudio de 1 668 especies, concluyeron que, en la naturaleza, los granos de polen isopolares y radioisométricos son los más comunes, seguidos de los apolares radioisométricos y los heteropolares bilaterales, aunque en una proporción menor.
Tras las observaciones realizadas, esta investigación representa el primer estudio morfológico del polen en el bosque seco tropical del recinto Quimis, lo que agrega un importante valor a la comprensión de la vegetación en la zona. Además, se ha comparado con trabajos previos realizados en Ecuador y varias regiones de América Latina, como es el caso de Ferreira et al. (2013).
En el caso de los recursos polínicos más frecuentes en los cinco apiarios objeto de estudio, se comprobó que proceden de las especies Convolvulus arvensis, Eriotheca ruizii, Ceiba trischistandra, Prosopis pallida con el 8,3 %. En ese mismo contexto las demás especies presentan un porcentaje bajo, sin embargo, estas plantas de alguna forma, logran ser utilizadas por la especie Apis mellifera como recurso polinífero alternativo y esto sucede cuando existe una baja disponibilidad de especies que aporten gran cantidad de polen alrededor de los apiarios.
Con referencia a lo anterior, Ramírez et al. (2016) sostienen que la procedencia floral del néctar, del cual las abejas producen la miel, y de las cargas de polen, son una fuente crucial de proteínas para la colonia, que puede ser identificada a través de análisis palinológicos. Por otro lado, Hidalgo et al. (1990) destacan la importancia de comprender la relación entre insectos y plantas, ya que esta relación arroja luz sobre las preferencias alimenticias de las abejas. De este modo, al conocer la vegetación de una región, se puede evaluar el potencial apícola de un área.
Frecuencia del peso en (g) del polen recolectado por mes de acuerdo con la cantidad de especies identificadas por cada apiario
En la Tabla 2, se expone el peso del polen como variable independiente mientras que el número de especies se considera como la variable dependiente y refleja la relación entre la cantidad de especies y el peso en gramos (g) de polen recolectado a lo largo de tres meses de observación en cada uno de los apiarios.
Tabla 2. - Cantidad de peso en g por número de especies por apiario del recinto Quimis
| Peso (g) | No. de especies por apiario |
| 44 | 11 |
| 15 | 11 |
| 42 | 15 |
| 27 | 14 |
| 24 | 9 |
Según se observa en la Tabla 2, los pesos del polen oscilan entre 15 g y 44 g, y el número de especies que va desde 9 hasta 15. Estos datos fueron analizados y se obtuvo una línea de tendencia que corresponde a un polinomio de grado 3, con un coeficiente de determinación R2 de 0,907. Esto sugiere una relación significativa entre el peso del polen y la diversidad de especies, lo que indica que a medida que aumenta el peso del polen recolectado, tiende a aumentar el número de especies presentes en los apiarios (Figura 5).
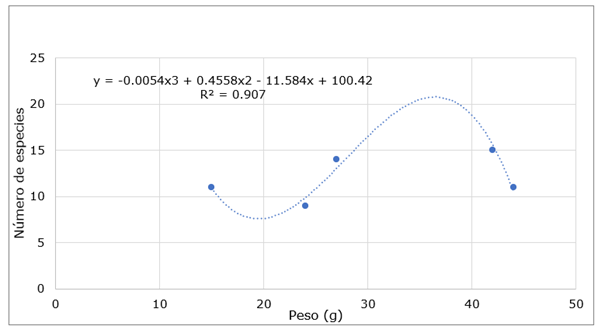
Fig. 5. - Cálculo de Polinomio grado 3 en relación con el peso del grano de polen de las especies melíferas del Bosque Seco Tropical del recinto Quimis
Mediante el cálculo de un polinomio de grado 3 relacionados con el peso del grano de polen en el Bosque Seco Tropical del recinto Quimis, se ha logrado obtener una herramienta que brinda una comprensión más precisa de la relación entre el peso del polen y la cantidad de especies presentes en la zona de estudio. Este modelo permite estimar tanto la cantidad mínima como máxima de especies que podrían existir en función del peso del polen. Los resultados muestran un ajuste a un modelo polinomio de grado 3, con un coeficiente de determinación (R2= 0,907). Este alto valor de R2 sugiere que el modelo polinómico es una buena representación de la relación entre el peso del polen y el número de especies, aspecto que indica que aproximadamente el 90,7 % de la variabilidad en el número de especies puede explicarse por la variación en el peso del polen, lo que refuerza la validez del modelo Ecuación 1, 2, 3 y 4.
Aplicando el cálculo diferencial de una variable real Ecuación 1, 2, 3 y 4:
Por todo lo antes expuesto, se puede obtener los extremos globales en el dominio de definición de interés, el valor mínimo global calculado es 19,52 g y aproximadamente ocho especies y el valor máximo 36,45 g y aproximadamente 21 especies. Estos hallazgos indican que, en la zona investigada, la estimación de la cantidad de especies melíferas se sitúa en un rango aproximado de 8 a 21.
En resumen, se han identificado los extremos globales de la función cúbica de tendencia, lo que proporciona información valiosa sobre la relación entre el peso del polen y la diversidad de especies en la zona de estudio.
Este estudio subraya la importancia del análisis estadístico y matemático para comprender las complejas relaciones biológicas en la ecología de las abejas, todo lo cual, resalta el papel crucial del peso del polen en la diversidad de especies, ofreciendo información relevante para la toma de decisiones en la gestión de apiarios y la conservación de la biodiversidad. Además, contrasta con estudios anteriores, como el de Jáuregui (2011), que emplearon diferentes métodos y enfoques para evaluar el rendimiento del polen y su impacto en la diversidad de especies melíferas.
Frecuencia con base en las características morfológicas identificadas en el polen de cada especie melífera
La Tabla 3 muestra cantidad de especies melíferas en función de las características morfológicas del polen. Las categorías estudiadas incluyen unidades de polen, simetría, aberturas, forma de exina, forma de polen y tamaño. Cada fila de la tabla presenta tanto la Frecuencia Absoluta (FA) como la Frecuencia Relativa (FR), además de mostrar la Frecuencia Absoluta Acumulada (FAA) y la Frecuencia Relativa Acumulada (FRA).
Tabla 3. - Cantidad de especies melíferas de acuerdo con las características morfológicas del polen
| Unidades de polen | FA | FR (%) | FAA | FRA |
| Mónada | 20 | 0,95 | 20 | 0,95 |
| Romboidal | 1 | 0,05 | 21 | 1,00 |
| Simetría | - | - | - | - |
| Asimétrico | 1 | 0,05 | 1 | 0,05 |
| Bisimétrico | 6 | 0,29 | 7 | 0,33 |
| Radiosimétrico | 14 | 0,67 | 21 | 1,00 |
| Aberturas | - | - | - | - |
| Diporados | 1 | 0,05 | 1 | 0,05 |
| Estefanocolpado | 1 | 0,05 | 2 | 0,10 |
| Inaperturados | 1 | 0,05 | 3 | 0,14 |
| Monoporados | 1 | 0,05 | 4 | 0,19 |
| Pantocolporado | 1 | 0,05 | 5 | 0,24 |
| Pantoporado | 1 | 0,05 | 6 | 0,29 |
| Polipantoporado | 1 | 0,05 | 7 | 0,33 |
| Tetracolporado | 1 | 0,05 | 8 | 0,38 |
| Tricolpado | 2 | 0,10 | 10 | 0,48 |
| Tricolporado | 10 | 0,48 | 20 | 0,95 |
| Triporado | 1 | 0,05 | 21 | 1,00 |
| Forma de exina | - | - | - | - |
| Equinada | 2 | 0,10 | 2 | 0,10 |
| Escábrida | 1 | 0,05 | 3 | 0,14 |
| Fosulada | 1 | 0,05 | 4 | 0,19 |
| Psilada | 9 | 0,43 | 13 | 0,62 |
| Reticulada | 8 | 0,38 | 21 | 1,00 |
| Forma del polen | - | - | - | - |
| Esferoidal | 8 | 0,38 | 8 | 0,38 |
| Esférico | 1 | 0,05 | 9 | 0,43 |
| Oblato | 3 | 0,14 | 12 | 0,57 |
| Oblato esferoidal | 2 | 0,10 | 14 | 0,67 |
| Prolato | 3 | 0,14 | 17 | 0,81 |
| Prolato-esferoidal | 2 | 0,10 | 19 | 0,90 |
| Sub-prolato | 2 | 0,10 | 21 | 1,00 |
| Tamaño | - | - | - | - |
| Grande | 5 | 0,24 | 5 | 0,24 |
| Mediano | 8 | 0,38 | 13 | 0,62 |
| Muy pequeño | 1 | 0,05 | 14 | 0,67 |
| Pequeño | 7 | 0,33 | 21 | 1,00 |
Nota: Frecuencia absoluta (FA); Frecuencia relativa (FR); Frecuencia absoluta acumulada (FAA); Frecuencia relativa acumulada (FRA).
De acuerdo con los resultados de la Tabla 3, específicamente, en la categoría de unidades de polen, se destaca que el 0,95 % corresponde a especies de tipo mónada. En relación con la simetría, el 0,67 % se caracteriza por ser de tipo radiosimétrico, mientras que el 0,48 % presenta aberturas tricolporadas. Con respecto a la forma de la exina, el 0,43 % exhibe una forma psilada, y el 0,38 % muestra una forma esferoidal. Por último, en lo que concierne al tamaño, el 0,38 % se clasifica como de tamaño mediano.
También se puede observar que las familias con dos o más especies fueron analizadas en función de sus características morfológicas para evaluar posibles relaciones entre ellas. Entre las familias estudiadas, Fabaceae se destacó al presentar cinco especies poliníferas, seguida de Malvaceae con tres especies, por otro lado, las familias Capparaceae y Convolvulaceae presentaron únicamente dos especies cada una.
Es relevante destacar que, en la mayoría de los caracteres, las especies de cada familia presentaron diferencias significativas, excepto en las unidades de polen y la simetría, donde se observó una mayor uniformidad. En este orden de ideas, las dos especies de la familia Convolvulaceae, compartieron el mismo tamaño de polen, mientras que las dos especies de la familia Capparaceae, exhibieron el mismo tipo de aberturas.
CONCLUSIONES
El estudio en cinco apiarios del bosque seco tropical de Quimis reveló 21 especies melíferas de 13 familias y 21 géneros, destacando Ceiba trischistandra, Convolvulus arvensis, Eriotheca ruizii y Prosopis pallida.
Se observaron diferencias morfológicas del polen entre especies de una misma familia, excepto en la unidad de polen y simetría.

